A new karst-dwelling bent-toed gecko(Squamata:Gekkonidae:Cyrtodactylus)from Xiangkhoang Province,northeastern Laos
2018-06-28RomanNazarovOlivierPauwelsEvgeniyKonstantinovAnatoliyChulisovNikolaiOrlovNikolayPoyarkovJr
Roman A.Nazarov,Olivier S.G.Pauwels,Evgeniy L.Konstantinov,Anatoliy S.Chulisov,Nikolai L.Orlov,Nikolay A.Poyarkov,Jr.
1Zoological Museum,Moscow State University,Moscow 125009,Russia
2Royal Belgian Institute of Natural Sciences,Brussels B-1000,Belgium
3Institute of Natural Sciences,Kaluga State University named after K.I.Tsiolkovskii,Kaluga 248023,Russia
4Zoological Institute,Russian Academy of Sciences,St.Petersburg 199034,Russia
5Department of Vertebrate Zoology,Biological Faculty,Lomonosov Moscow State University,Moscow 119234,Russia
6Joint Russian-Vietnamese Tropical Research and Technological Center,63 Nguyen Van Huyen Road,Nghia Do,Cau Giay,Hanoi,Vietnam
INTRODUCTION
The diversity of the gecko genusCyrtodactylushas increased at a phenomenal rate.Of the more than 250 species recognized within the genus,about 80%of the diversity has been described in the twenty-first century(Uetz et al.,2018). Numbers ofCyrtodactylusspecies in Thailand and Vietnam have shown considerable expansion,including at least 33 and 39 recognized species in each country,respectively(Pauwels et al.,2016).Moreover,several species remain to be named in Vietnam,in particular within theCyrtodactylus irregularisspecies complex(Nguyen et al.,2017).Although Laos is located between these two species-rich countries and harbors a varied geological relief propitious to micro-endemism,itsCyrtodactylusfauna is currently known to include only 21 species(Luu et al.,2016a;Nazarov et al.,2014).Such low species number may be due to the fewer herpetological surveys conducted in Laos compared to Vietnam and Thailand,particularly on the karst reliefs(Teynié&David,2014).
Our team co-described fi ve of theCyrtodactylusspecies currently recorded from Laos(Nazarov et al.,2014;Ngo&Pauwels,2010),and we are pursuing our efforts to better inventory the diversity of the genus in this country.
During fieldwork in the forested karst massifs in the Laotian province of Xiangkhoang,we(A.S.C.and E.L.K.)encountered three geckos that differed in color pattern and scalation from known species,and which we show hereafter to belong to an undescribed species.Morphological characters,such as scalation and dorsal color and pattern,indicate that this newly discoveredCyrtodactyluspopulation clearly belongs to theC.phongnhakebangensisspecies groupsensuLuu et al.(2016a),which includes a dozen species from Laos and Vietnam.In agreement with the definition of this group provided by Luu et al. (2016a:132),this new species exhibits a maximum adult SVL comprised between 73.0 and 100.6 mm,0 or 1 supranasal,a DorTub(14-18)between 10 and 24,no webbing between fingers or toes,tubercles nearly absent on forelimbs but present on hind limbs,precloacal and femoral pores in males(in total 47)between 20 and 60,number of postcloacal tubercles(5 or 6 in male)between 3 and 8,enlarged subcaudals,and dorsum displaying well-defined dark bands.
MATERIALS AND METHODS
Sampling
Field surveys were conducted in Xiangkhoang Province,Laos,from May to July 2008 within the frameworks of the Memorandum of Understanding(MOU)on the Academic and Scientific Cooperation between the National University of Laos and Kaluga State University,Russia(MOU No.72 20-11-2012).Specimens were collected by hand from 1900 h to 2300 h.Specimens were photographed in life and subsequently euthanized in a closed vessel with a piece of cotton wool containing ethyl acetate(Simmons,2002).Tissue samples were preserved separately in 95%ethanol and specimens were fixed in 85%ethanol,after which they were transferred to 70%ethanol for permanent storage. Specimens were subsequently deposited in the herpetological collection of the Zoological Institute of the Russian Academy of Science(ZISP)and in the herpetological collection of the Zoological Museum of Lomonosov Moscow State University(ZMMU),Moscow,Russia.
The specimens accessed for molecular analyses are listed in Table 1. Our sampling is to date the most comprehensive sampling covering all known lineages of theC.phongnhakebangensisspecies complex.A map showing the distribution of theC.phongnhakebangensisspecies complex in Laos and adjacent territories and the location of the sampling site for the present work is provided in Figure 1.Museum abbreviations used are listed as follows:CAS:California Academy of Sciences,San Francisco;CUMZ R:Chulalongkorn University Museum of Zoology(Reptiles),Bangkok;IRSNB:Institut Royal des Sciences Naturelles de Belgique,Brussels;KZM:Khorat Zoo Museum,Nakhon Ratchasima;MNHN:Muséum National d’Histoire Naturelle,Paris;PSUZC-RT:Prince of Songkhla University Zoological Collection,Reptile Section,Songkhla;QSMI:Queen Saovabha Memorial Institute,Thai Red Cross Society,Bangkok;THNHM:Thailand Natural History Museum,National Science Museum,Technopolis,Pathum Thani;ZMMU:Zoological Museum,Moscow State University,Moscow;ZISP:Zoological Institute,Saint Petersburg.Comparisons with congeneric species are based on literature data and on direct examination of preserved specimens(Appendix I).
Morphological descriptions
Measurements and meristic counts follow Pauwels et al.(2016).Measurements were taken on the right side.Paired meristic characters are given in left/right order. Numbers of supralabial and infralabial scales were counted from the largest scale immediately posterior to the dorsal inflection of the posterior portion of the upper jaw to the rostral and mental scales,respectively;the number of longitudinal rows of body tubercles was counted transversely across the center of the dorsum from one ventrolateral skin fold to the other;the number of longitudinal rows of ventral scales was counted transversely across the center of the abdomen from one ventrolateral skin fold to the other;subdigital lamellae beneath the toes were counted from the base of the first phalanx to the claw;dorsal dark bands between limb insertions are those strictly contained on the dorsum between the posterior insertion of the anterior limbs and the anterior insertion of the posterior limbs. In previous literature,dorsal bands often include those contained between the anterior insertion point of the anterior limbs and the posterior point of insertion of the posterior limbs,thus interspecific comparisons on band numbers were performed with caution.
The following measurements were taken with a digital caliper to the nearest 0.1 mm:AG:axilla to groin length,taken from the posterior margin of the forelimb at its insertion point on the body to the anterior margin of the hind limb at its insertion point on the body;EarL:ear length,the greatest horizontal distance of the ear opening;ForeaL:forearm length,taken on the dorsal surface from the posterior margin of the elbow while flexed 90°to the inflection of the flexed wrist;HeadH:head height,the maximum height of head from the occiput to the throat;Head L:head length,from the posterior margin of the retroarticular process of the lower jaw to the tip of the snout;Head W:head width,measured at the angle of the jaws;Internar:internarial distance,measured between the nares across the rostrum;Interorb:interorbital distance,measured between the anterior edges of the orbits;ML:mental length,the maximum length of mental shield;MW:mental width,the maximum width of mental shield;NosOrb:nostril to orbit distance,from the posterior margin of the external nares to the anterior margin of the orbit;OrbD:orbit diameter,the greatest horizontal diameter of the orbit;OrbEar:orbit to ear distance,from the anterior edge of the ear opening to the posterior edge of the orbit;RH:rostral height,the maximum height of the rostral shield;RW:rostral width,the distance between border of rostral shield and the first supralabial scales on right and left sides;SnOrb:snout to orbit distance,from the tip of the snout to the anteriormost margin of the orbit;SVL:snout-vent length,taken from the tip of snout to the vent;TailL:tail length,taken from the vent to the tip of the tail,original or regenerated;TailW:tail width,taken at the base of the tail immediately posterior to the postcloacal swelling;TibiaL:tibia length,taken on the ventral surface from the posterior surface of the knee while flexed 90°to the base of heel. Meristic characters abbreviations:DorTub:number of longitudinal rows of dorsal tubercles at midbody;EnlFemSc:enlarged femoral scales;FemPi:femoral pits;FemPo:femoral pores;FemPreclPo:number of femoral and precloacal pores in continuous series;IL:infralabial scales;InterorbSc:interorbital scales;ParaTub:number of paravertebral tubercles between the limbs insertions,counted in a straight line immediately left of the vertebral column;PreclPi:precloacal pits(shallow depressions without waxy exudates);PreclPo: precloacal pores(deeper than pits,and with waxy exudates);SL:supralabial scales;SLF4:subdigital lamellae beneath 4thfinger(basal and distal lamellae combined);SLT4:subdigital lamellae beneath 4thtoe(basal and distal lamellae combined);Ven:number of ventral scale rows.

Figure 1 General distribution of the genus Cyrtodactylus in Laos and surrounding areas
DNA isolation,PCR,and sequencing
For molecular phylogenetic analyses,total genomic DNA was extracted from ethanol-preserved femoral muscle tissue(Table 1)using standard phenol-chloroform-proteinase K( final concentration 1 mg/mL)extraction procedures with consequent isopropanol precipitation(protocols followed Hillis et al.,1996 and Sambrook et al.,1989).The isolated total genomic DNA was visualized using agarose electrophoresis in the presence of ethidium bromide. The concentration of total DNA was measured in 1µL using NanoDrop 2000(Thermo Scientific,USA),and consequently adjusted to ca.100 ng DNA/µL.
The mitochondrial cytochrome oxidase subunit I(COI)was selected as a genetic marker to clarify the taxonomic position of the newly discovered population ofCyrtodactylus. A total of 655 bp ofCOIwas amplified using a mitochondrial marker widely used as a barcoding marker for vertebrates,including both reptiles and amphibians (Murphy etal.,2013; Nagy etal.,2012; Smith et al.,2008),and which proved to be useful for species identification and assessment of cryptic diversity in various groups of lizards,including the genusCyrtodactylus(Brennan et al.,2017;Hartmann et al.,2013;Luu et al.,2016a;Nazarov et al.,2012,2014;Nguyen et al.,2013,2014,2017;Solovyeva et al.,2011).Primers used both for PCR and sequencing were the VF1-d(5′-TTCTCAACCAACCACAARGAYATYGG-3′)and the VR1-d(5′-TAGACTTCTGGGTGGCCRAARAAYCA-3’)(following Ivanova et al.,2006). The obtained fragments were sequenced in both directions for each sample,and a consensus sequence was generated.PCR analyses were performed in 25µL reactions using ca.50 ng genomic DNA,10 pmol of each primer,15 nmol of each dNTP,50 nmol additional MgCl2,Taq PCR buffer(10 mmol/L Tris-HCl,pH 8.3,50 mmol/L KCl,1.1 mmol/L MgCl2,and 0.01%gelatin),and 1 U of Taq DNA polymerase.The PCR conditions for theCOIgene fragment followed Nazarov et al.(2012)and included an initial denaturation step at 95°C for 3 min;5 cycles at 95°C for 30 s,annealing at 45°C for 1 min,extension at 72°C for 2 min,followed with 35 cycles at 95°C for 30 s,annealing at 51°C for 1 min,extension at 72°C for 2 min,and a fi nal extension at 72°C for 5 min.
The PCR products were loaded onto 1.5%agarose gels in the presence of ethidium bromide and visualized using agarose electrophoresis. If distinct bands were produced,products were purified using 2µL,from a 1:4 dilution of ExoSapIt(Amersham,UK),per 5µL of PCR product prior to cycle sequencing.A 10 µL sequencing reaction included 2 µL of template,2.5 µL of sequencing buffer,0.8 µL of 10 pmol primer,0.4µL of BigDye Terminator version 3.1 Sequencing Standard(Applied Biosystems),and 4.2µL of water.The cycle sequencing reaction included 35 cycles of 10 s at 96°C,10 s at 50°C,and 4 min at 60°C.Cycle sequencing products were purified by ethanol precipitation. Sequence data collection and visualization were performed on an ABI 3730xl automated sequencer(Applied Biosystems,USA).The obtained sequences were deposited in GenBank under the accession numbers MG791873-MG791875(Table 1).
Phylogenetic analyses
A total of 66COIsequences of congeners were obtained from GenBank for phylogenetic analyses,including the three sequences obtained in this study(Table 1).
Nucleotide sequences were initially aligned using ClustalX 1.81(Thompson et al.,1997)with default parameters,and then optimized manually in BioEdit 7.0.5.2(Hall,1999)and MEGA 6.0(Tamura et al.,2013).Mean uncorrected genetic distances(P-distances)between sequences were determined with MEGA 6.0.MODELTEST v.3.06(Posada&Crandall,1998)was used to estimate the optimal models of DNA evolution. The best- fitting model selected forCOIdataset was K80+I+G for the first and the second codon positions and GTR+G for the third codon position,as suggested by the Akaike Information Criterion(AIC).
Phylogenetic trees were inferred using two different methods:Bayesian inference(BI)and maximum likelihood(ML).BI was conducted in MrBayes 3.1.2(Huelsenbeck&Ronquist,2001;
Ronquist&Huelsenbeck,2003). Metropolis-coupled Markov chain Monte Carlo(MCMCMC)analyses were run with one cold chain and three heated chains for four million generations and sampled every 1 000 generations.Five independent MCMCMC runs were performed and 1 000 trees were discarded as burn-in.Confidence in tree topology was assessed by posterior probability(BI PP)(Huelsenbeck&Ronquist,2001).The ML analyses were conducted using Treefinder(Jobb et al.,2004).TheCOIsequence ofCyrtodactylus elokDring was used as an outgroup following Luu et al.(2016a).Confidence in tree topology was tested by non-parametric bootstrap analysis(ML BS)with 1 000 replicates(Felsenstein,2004).Wea prioriregarded tree nodes with bootstrap(ML BS)values of 70%or greater and posterior probabilities(BI PP)values over 0.95 as sufficiently resolved,ML BS values between 70%and 50%(and BI PP between 0.95 and 0.90)were regarded as tendencies,and ML BS values below 50%(BIPP below 0.90)were considered unresolved(Felsenstein,2004;Huelsenbeck&Hillis,1993).
RESULTS
Genetic differentiation Sequence data
The fi nal alignment of the examined mtDNACOIgene fragment consisted of 672 sites:386 sites were conserved and 286 sites were variable,of which 270 were found to be potentially parsimony-informative.The transition-transversion bias(R)was estimated as 3.66.Nucleotide frequencies were A=23.14%,T=26.77%,C=30.53%,and G=19.95%.
mtDNA genealogy
Results of the phylogenetic analyses implemented in the present paper are shown in Figure 2. The BI and ML analyses resulted in essentially similar topologies slightly differing from each other only in associations at several poorly supported basal nodes. In general,the genetic differentiation of theCyrtodactylus phongnhakebangensisspecies group members was highly consistent with the results reported in previous studies by Nazarov et al. (2014)and Luu et al. (2016a). The partial fragment of theCOIgene strongly supports monophyly of theC.phongnhakebangensis(1.0/100,hereafter node support values are given for BI PP/ML BS,respectively),C.wayakonei(1.0/86),andC.irregularis(0.99/83)species groups.Within theC.phongnhakebangensisspecies group,C.pageliSchneider,Nguyen,Schmitz,Kingsada,Auer&Ziegler,2011 formed a sister lineage with respect to all other members of theC.phongnhakebangensisspecies group(0.97/75).The remaining members of theC.phongnhakebangensisspecies group were clustered in two subclades with moderate to strong support values:(1)subclade consisting ofC.khammouanensisNazarov,Poyarkov,Orlov,Nguyen,Milto,Martynov,Konstantinov&Chulisov,2014,C.ruffordLuu,Calame,Nguyen,Le,Bonkowski&Ziegler,2016,C.bansocensisLuu,Nguyen,Le,Bonkowski&Ziegler,2016,C.soudthichakiLuu,Calame,Nguyen,Bonkowski&Ziegler,2015,C.jaegeriLuu,Calame,Bonkowski,Nguyen&Ziegler,2014,C.sommerladiLuu,Bonkowski,Nguyen,Le,Schneider,Ngo&Ziegler,2016,andC.roesleriZiegler,Nazarov,Orlov,Nguyen,Vu,Dang,Dinh&Schmitz,2010(0.95/64);and(2)subclade consisting ofC.hinnamnoensisLuu,Bonkowski,Nguyen,Le,Schneider,Ngo&Ziegler,2016,C.darevskiiNazarov,Poyarkov,Orlov,Nguyen,Milto,Martynov,Konstantinov&Chulisov,2014,C.calameiLuu,Bonkowski,Nguyen,Le,Schneider,Ngo&Ziegler,2016,C.phongnhakebangensisZiegler,Rösler,Herrmann&Vu,2002,C.lomyenensisNgo&Pauwels,2010,C.multiporusNazarov,Poyarkov,Orlov,Nguyen,Milto,Martynov,Konstantinov&Chulisov,2014,C.teynieiDavid,Nguyen,Schneider&Ziegler,2011,C.cf.jarujiniUlber,1993,andCyrtodactylussp.from Ban Thathom,Xiangkhoang Province,Laos(0.94/78).
The newly discoveredCyrtodactyluspopulation from Ban Thathom,Xiangkhoang Province,was placed within the first subclade of theC.phongnhakebangensisspecies group and was clustered in one group withC.multiporus,C.teyniei,andC.cf.jarujini(1.0/99).Among these species,the Ban Thathom population appeared to be a sister lineage ofC.cf.jarujinifrom the Bolikhamxay Province of Laos(1.0/100).
Genetic distances
The uncorrected geneticP-distances among and within theCOIgene fragment of the studied members of theCyrtodactylus phongnhakebangensisspecies group are summarized in Table 2.The observed interspecific distances in theCOIgene within theC.phongnhakebangensisspecies groupvariedfromP=4.2% (betweenC.darevskiiandC.hinnamnoensis)toP=19.4%(betweenC.soudthichakiandC.hinnamnoensis)(Table 2). These values slightly overlapped with interspecific comparisons between theC.phongnhakebangensisspecies groupmembers and the outgroupCyrtodactylusspecies(18.1%<P<24.4%,data not shown in Table 2).The observed intraspecific distances in our analysis varied fromP=0%toP=3.8%,with the last value corresponding to the genetic differentiation between mtDNA lineages ofC.pageli(Table 2).

Figure 2 Bayesian inference(BI)tree for the Cyrtodactylus phongnhakebangensis species group and other Cyrtodactylus species groups inhabiting Laos and adjacent areas based on analysis of COI gene fragments

Table 1 Specimens and sequences of Cyrtodactylus representatives used in molecular analyses of mtDNA gene fragment

Table 2 Uncorrected P-distances(percentages)between COI sequences of the Cyrtodactylus phongnhakebangensis species group members included in phylogenetic analyses(below the diagonal),and standard error estimates(above the diagonal)
The newly discoveredCyrtodactyluspopulation from Ban Thathom,Xiangkhoang Province,Laos,is genetically divergent from all other members of theC.phongnhakebangensisspecies group,and it is most closely related toC.multiporus(P=9.5%),C.teyniei(P=9.3%),andC.cf.jarujini(P=4.5%).The last population from Bolikhamxay Province of Laos was tentatively identi fi ed asC.cf.jarujiniby Luu et al.(2016a);however,its taxonomic status requires further con fi rmation.
Systematics
Based on the morphological,chromatical,and genetic distinctiveness of the newly discoveredCyrtodactyluspopulation from all other populations in northern Laos and neighboring areas(see Diagnosis and Comparisons),and also on the results of phylogenetic analyses of the partialCOIgene fragment,indicating that this population represents a clearly distinct mtDNA lineage,different from all congeners for which homologous sequences are available(see Results),we conclude that the Ban Thathom population represents a previously undescribed new species,which we describe and name herein.
Cyrtodactylus thathomensis sp.nov.(Figures 3-6)
Holotype:ZMMU R-14919-1(Figure 3);an adult male from the north-western slope of a limestone hill(N18°59′48.9′′,E103°35′30.6′′;alt.271 m a.s.l.)near Ban(=Village)Thathom,Xiangkhoang Province,Laos.Collected on 14 May 2008 by E.L.Konstantinov and A.S.Chulisov.
Paratypes:ZMMU R-15384 and ZISP 29731;two adult females from the same locality
Diagnosis:Cyrtodactylus thathomensissp.nov.can be distinguished from all other congeneric species by its medium body size(maximal known SVL 75.5 mm);dorsal tubercles in 14-18 rows at midbody;midbody scale rows 30-36 across belly between ventrolateral skin folds;a continuous series of 10-12 pore-bearing(male)or pitted(females),enlarged precloacal scales,separated by a diastema from a series of enlarged femoral scales bearing 18 or 19 pores(male)or 1-9 pits(females)on each femur;absence of precloacal groove;transversely enlarged median subcaudal scales;and four dark dorsal bands between limb insertions.
Description of holotype:Adult male. SVL 70.3 mm.TailL 79.0 mm(last 54 mm regenerated). Head relatively long(HeadL 20.1;HeadL/SVL 0.29)and wide(HeadW 13.4;HeadW/HeadL 0.67),not markedly depressed(HeadH 8.4),distinct from neck. Loreal region inflated,canthus rostralis slightly prominent. Snout elongate(SnOrb/HeadL 0.41),rounded,longer than orbit diameter(OrbD/SnOrb 0.62).Scales on snout small,rounded to oval,granular to weakly conical,mostly homogeneous,larger than those on crown,interorbital,and occipital regions.Eye relatively large(OrbD/HeadL 0.25);pupil vertical with crenelated margins;supraciliaries short,larger anteriorly. Ear opening vertically oval,of moderate size(EarL/HeadL 0.06);orbit to ear distance subequal to orbit diameter(OrbEar/OrbD 1.02).Rostral much wider(3.5 mm)than deep(2.0 mm);rostral crease straight,starting from the upper middle of the rostral,going down half the rostral height. Two enlarged supranasals separated from one another by one internasal. Rostral contacting first supralabial on each side,nostrils,two supranasals and one internasal.Nostrils rounded,more or less laterally directed,each surrounded by supranasal,rostral, first supralabial and two postnasals.Three or four rows of small scales separate orbit from supralabials.Mental triangular,wider(3.4 mm)than deep(2.2 mm).A single pair of greatly enlarged postmentals in broad contact behind mental,each bordered anteromedially by mental,anterolaterally by first infralabial,posterolaterally by an enlarged lateral chinshield,and posteriorly by two granules(in total three granules contact the postmentals)(Figure 3E).Supralabials to mid-orbital position 9/9,enlarged supralabials to angle of jaws 10/11.Infralabials 9/9(Figure 3D).Interorbital scale rows across narrowest point of frontal bone 30.

Figure 3 Male holotype of Cyrtodactylus thathomensis sp.nov.(ZMMU R-14919)
Body moderately slender,relatively short(AG/SVL 0.43)with poorly defined,non-denticulate,ventrolateral skin folds.Dorsal scales weakly heterogeneous,domed,with tubercles about four times size of adjacent dorsal scales extending from neck onto tail base,smaller tubercles on postocular region,crown and occiput;tubercles smooth or bearing a very small keel,tubercles on posterior trunk and sacral region most prominent.Dorsal tubercles in 18 rows at midbody,typically separated from one another by two dorsal granules(Figure 3F).Paravertebral tubercles 26.Ventral scales larger than dorsal scales,smooth,oval and subimbricate,largest in precloacal region.Midbody scale rows across belly between ventrolateral folds 30.Gular region with homogeneous,smooth,juxtaposed granular scales.A patch of enlarged precloacal scales on top of which lies a continuous series of ten pore-bearing scales,separated by a diastema of 3/4 enlarged poreless scales from a continuous series of 19/18 pore-bearing femoral scales.Enlarged femoral scales nearly two times the size of the scales of the adjacent anterior scale row(Figure 3B).No precloacal groove.Hemipeneal bulges evident.Postcloacal spurs bearing 6/5 enlarged conical scales(Figure 3C).
Scales on palm and sole smooth,rounded to oval or hexagonal,slightly domed.Scalation on dorsal surfaces of hind limbs similar to body dorsum with enlarged tubercles interspersed among smaller scales;tubercles smaller and rare on forelimbs. Forelimbs and hind limbs moderately long(ForeaL/SVL 0.16,TibiaL/SVL 0.19),moderately slender.Digits long,slender,inflected at interphalangeal joints,all bearing robust,slightly recurved claws. Basal subdigital lamellae broad,oval to rectangular,without scansorial surfaces;lamellae distal to digital inflection narrow;SLF4 17/17;SLT4 20/20. Subcaudals scales larger than supracaudal scales,forming a row of strongly enlarged transverse plates.
Coloration in preservative:Dorsal ground color of head,neck,body,limbs and tail light brown.Dorsal surface of head with irregular dark brown markings.Rostral,supralabials and infralabials dark brown,posterior ones heavily maculated with beige.On each side a postocular stripe reaching the nape but not meeting the one of the opposite side as it breaks into spots(i.e.,a discontinuous nuchal collar).Upper surface of limbs showing irregular dark brown bands.Dorsum showing four dark brown bands between limb insertions.Each of the four bands on dorsum posteriorly limited by a discontinuous series of whitish tubercles,similarly to the band above shoulders,the one on the neck and the one above sacrum.Original part of the tail showing three dark brown bands.Regenerated part of the tail light brown.Under surfaces of the head,throat,venter and members uniformly beige.Coloration of the holotype in life is shown in Figure 3A.In life its dorsal ground color is darker than in preservative.The tubercles posteriorly bordering the four bands on dorsum are lighter and more contrasting than after preservation.The yellow color of the outer extremities of the supra ciliaries disappears in preservative.
Variation:Morphometric and meristic values for the type series are provided in Table 3.Morphological and coloration characters of the paratypes agree in most respects with those of the holotype,differing only in minor details(Table 3 and remarks hereafter).Rostral crease in paratypes like in holotype.One internasal in paratype ZISP 29731 like in holotype(Figure 4C),no internasal in paratype ZMMU R-15384(hence supranasals in large contact)(Figure 5C).The female paratype ZISP 29731 shows a continuous row of 52 enlarged femoral-precloacal scales including,from left to right:1 pitless scale+9 scales with pits+12 pitless scales+10 precloacal scales with pits+11 pitless scales+9 scales with pits+1 pitless scale(Figure 4E).The female paratype ZMMU R-15384 shows a continuous row of 55 enlarged femoral-precloacal scales including,from left to right:10 scales with pits+11 pitless scales+12 enlarged precloacal scales with pits+12 pitless scales+8 scales with pits+2 pitless scales(Figure 5E).General background dorsal color in preservative lighter in the female paratypes than in the male holotype.The paratypes’tails,original and complete,show nine dark brown bands,and they are longer than SVL(TailL/SVL ratio 1.16-1.27;see Figures 4A-5A,Figure 6).
Comparison with congeneric species:Synoptic tables comparing the main morphological characters of LaoCyrtodactyluswith the species known from adjacent regions were provided by Teynié&David(2014:471-472)and Luu et al. (2016a,2016b,2016c). Among these species,we are comparing hereafterCyrtodactylus thathomensissp.nov.with all congeneric species found within a 500-km radius from its type-locality(a radius far superior to any maximal distance between two localities known for any species in the Indochinese Region,Thailand and Myanmar,cf.maps provided by Ellis&Pauwels,2012;Nazarov et al.,2014:Figure 1;Grismer et al.,2015:Figure 1-considering thatCyrtodactylus intermediusis a species complex composed of at least six species,loc.cit.:114;Nguyen et al.,2014,2017:Figure 1).
By its possession of transversely enlarged subcaudals,the new species is readily distinguished fromCyrtodactylus angularis(Smith,1921),C.buchardiDavid,Teynié&Ohler,2004,C.cryptusHeidrich,Rösler,Vu,Böhme&Ziegler,2007,C.jarujini,C.papilionoidesUlber&Grossmann,1991,C.pseudoquadrivirgatusRösler,Nguyen,Vu,Ngo&Ziegler,2008,andC.vilaphongiSchneider,Nguyen,Le,Nophaseud,Bonkowski&Ziegler,2014. The possession of enlarged femoral scales separatesCyrtodactylus thathomensissp.nov.fromC.bobroviNguyen,Le,Pham,Ngo,Hoang,Pham&Ziegler,2015,C.buchardi,C.chauquangensisHoang,Orlov,Ananjeva,Johns,Hoang&Dau,2007,C.cryptus,C.otaiNguyen,Le,Pham,Ngo,Hoang,Pham&Ziegler,2015,C.pageli,C.pseudoquadrivirgatus,C.spelaeusNazarov,Poyarkov,Orlov,Nguyen,Milto,Martynov,Konstantinov&Chulisov,2014,C.vilaphongiandC.wayakoneiNguyen,Kingsada,Rösler,Auer&Ziegler,2010.

Figure 4 Female paratype of Cyrtodactylus thathomensis sp.nov.(ZISP 29731)

Figure 5 Female paratype of Cyrtodactylus thathomensis sp.nov.(ZMMU 15384)

Table 3 Meristic and morphometric(in mm)data for the type series of Cyrtodactylus thathomensis sp.nov.
The presence of precloacal and femoral pores separated by a diastema in maleCyrtodactylus thathomensissp.nov.distinguishes this species fromC.angularis,C.bansocensis,C.bobrovi,C.buchardi,C.calamei,C.chanhomeaeBauer,Sumontha&Pauwels,2003,C.chauquangensis,C.cryptus,C.cucphuongensisNgo&Chan,2011,C.darevskii,C.hinnamnoensis,C.jaegeri,C.jarujini,C.khammouanensis,C.lomyenensis,C.martiniNgo,2011,C.multiporus,C.otai,C.pageli,C.papilionoides,C.phongnhakebangensis,C.pseudoquadrivirgatus,C.puhuensisNguyen,Yang,Le,Nguyen,Orlov,Hoang,Nguyen,Jin,Rao,Hoang,Che,Murphy&Zhang,2014,C.roesleri,C.rufford,C.sommerladi,C.soudthichaki,C.spelaeus,C.vilaphongiandC.wayakonei.Cyrtodactylus teynieiwas described based on a single female holotype. Teynié&David(2014)reported the first known male;it showed precloacal and femoral pores,but they did not mention if the femoral and precloacal pored scales were separated by a diastema or not.It should be noted that,while in the male holotype ofCyrtodactylus bansocensisthe precloacal and femoral pore-bearing scales are separated by a diastema,they are not in the male paratype(Luu et al.,2016c).
Its banded dorsalpattern separatesCyrtodactylus thathomensissp. nov. fromC.buchardi,C.jarujini,C.multiporus,C.pseudoquadrivirgatus,C.spelaeusandC.teyniei,which show blotched patterns.Additional differences distinguishing the new species fromCyrtodactylus jarujiniinclude its smaller SVL(75.5 vs.90 mm inC.jarujini)and a lower total number of pores in males(47 vs. 52-54).Additional differences withCyrtodactylus multiporusinclude the new species’much smaller SVL(75.5 vs.98 mm inC.multiporus)and a much lower total number of pores in males(47 vs.58-60).FromCyrtodactylus teynieithe new species differs also by its smaller SVL(75.5 vs.89.9 mm inC.teyniei),generally lower number of midbody scale rows across belly between ventrolateral folds(30-36 vs.36-38),and a much lower total number of pores in males(47 vs.58).

Figure 6 Live female paratype of Cyrtodactylus thathomensis sp.nov.(ZISP 29731)in dorsal view(Photo by E.L.Konstantinov)
Among the remaining species living within the 500-km radius of its type-locality,Cyrtodactylus thathomensissp.nov.can be differentiated fromC.auribalteatusSumontha,Panitvong&Deein,2010 by its much smaller SVL(75.5 vs.98.1 mm),lower DorTub(14-18 vs.22-24),lower Ven(30-36 vs.38-40),and higher FemPo on each side(18 or 19 vs.4 or 5)and PreclPo(10 vs. 6)in males;fromC.bichnganaeNgo&Grismer,2010 by its much smaller SVL(75.5 vs.99.9 mm),much higher EnlFemSc on each side(20-22 vs.11-13),and much higher FemPo on each side in males(18 or 19 vs.9);fromC.doisuthepKunya,Panmongkol,Pauwels,Sumontha,Meewasana,Bunkhwamdi&Dangsri,2014 by its smaller SVL(75.5 vs.90.5 mm),lower DorTub(14-18 vs.19 or 20),presence of femoral pores(vs. pits)in males,higher PreclPo(10 vs.6),and 4(vs.5 or 6)dark bands between limb insertions;fromC.dumnuii Bauer,Kunya,Sumontha,Niyomwan,Pauwels,Chanhome&Kunya,2010 by its generally lower DorTub(14-18 vs.18-22),its lower Ven(30-36 vs.40),and much higher FemPo on each side(18 or 19 vs.6 or 7)and PreclPo(10 vs.5 or 6)in males;fromC.huongsonensisLuu,Nguyen,Do&Ziegler,2011 by its by its smaller SVL(75.5 vs.89.8 mm),lower Ven(30-36 vs.41-48),higher PreclPo(10 vs.6)and much higher FemPo on each side(18 or 19 vs.7-10)in males,and its discontinuous(vs.continuous)nuchal loop;fromC.interdigitalisUlber,1993 by its generally lower DorTub(14-18 vs.18-22),its lower Ven(30-36 vs.37-42),much higher FemPo on each side(18 or 19 vs.9)and lower PreclPo(10 vs.14)in males,and absence(vs.presence)of webbing;fromC.intermedius(Smith,1917)by its lower Ven(30-36 vs.40-50)and much higher EnlFemSc on each side(20-22 vs.6-10);fromC.inthanonKunya,Sumontha,Panitvong,Dongkumfu,Sirisamphan&Pauwels,2015 by its generally lower DorTub(14-18 vs.18-20),and much higher FemPo on each side(18 or 19 vs.6)and PreclPo(10 vs.5)in males;fromC.khelangensisPauwels,Sumontha,Panitvong&Varaguttanonda,2014 by its much smaller SVL(75.5 vs.95.3 mm),and much higher FemPo on each side(18 or 19 Po vs.6 or 7 Po or Pi)and PreclPo(10 vs.2-5)in males;fromC.kunyai Pauwels,Sumontha,Keeratikiat&Phanamphon,2014 by its much higher FemPo on each side(18 or 19 vs.5 or 6)and PreclPo(10 vs.3)in males;and fromC.soni Le,Nguyen,Le&Ziegler,2016 by its much smaller SVL(75.5 vs.103.0 mm),higher DorTub(14-18 vs.10-13),lower Ven(30-36 vs.41-45),and much higher FemPo on each side(18 or 19 vs.6-8)and PreclPo(10 vs.6 or 7)in males.
Besides differentiatingCyrtodactylus thathomensissp.nov.from all congeneric species found within a 500-km radius,its combination of characters presented in the Diagnosis allows to unambiguously separate it from all species found in Bangladesh,Cambodia,Myanmar,Thailand and Vietnam(see,among other references in the literature cited,Bauer,2003;Connette et al.,2017;Grismer et al.,2012;Le et al.,2016;Mahony et al.,2009;Mahony,2009;Panitvong et al.,2014;Pauwels&Sumontha,2014;Pauwels et al.,2014,2016;Sumontha et al.,2014,2015).
Distribution and natural history:Cyrtodactylus thathomensissp.nov.is so far known only from its type-locality.The types were collected on karst boulders on a steep forested limestone hill(Figure 7).

Figure 7 Habitat of Cyrtodactylus thathomensis sp.nov.at the type-locality
Phylogenetic position:The new species is a member of theC.phongnhakebangensisspecies groupsensuLuu et al.(2016a)within which it is most closely related toC.multiporus,C.teynieiandC.cf.jarujini(see Results).
Etymology:The speci fi c epithet“thathomensis”is a Latinized toponymic adjective,referring to the type locality of the new species,Ban Thathom.We suggest the following common names:Ki Chiem Thathom(Lao),Tuk Khai Thathom(Thai),Thathom Bent-toed Gecko(English),Tatomskiy Krivopalyi Gekkon(Russian),Thathomkromvingergekko(Dutch),andCyrtodactyle de Thathom(French).
DISCUSSION
Mitochondrial genealogy indicatesCyrtodactylus thathomensissp. nov. as a member of theC.phongnhakebangensisspecies group,within which it appears to be most closely related toC.multiporusandC.teyniei.Table 4 compares the main diagnostic characters of the species included in theC.phongnhakebangensisspecies groupsensuLuu et al.(2016a)and our new species.All species in this group,including the one newly described here,live in the Cammon Plateau and the southern edge of the Xiangkhoang Plateau in the Annam Cordillera.This species group currently represents 15 out of the 22Cyrtodactylusspecies presently recorded from Laos(i.e.,68%),with the remaining seven species belonging to theC.interdigitalis,C.irregularisandC.wayakonei species groupssensuLuu et al.(2016a).
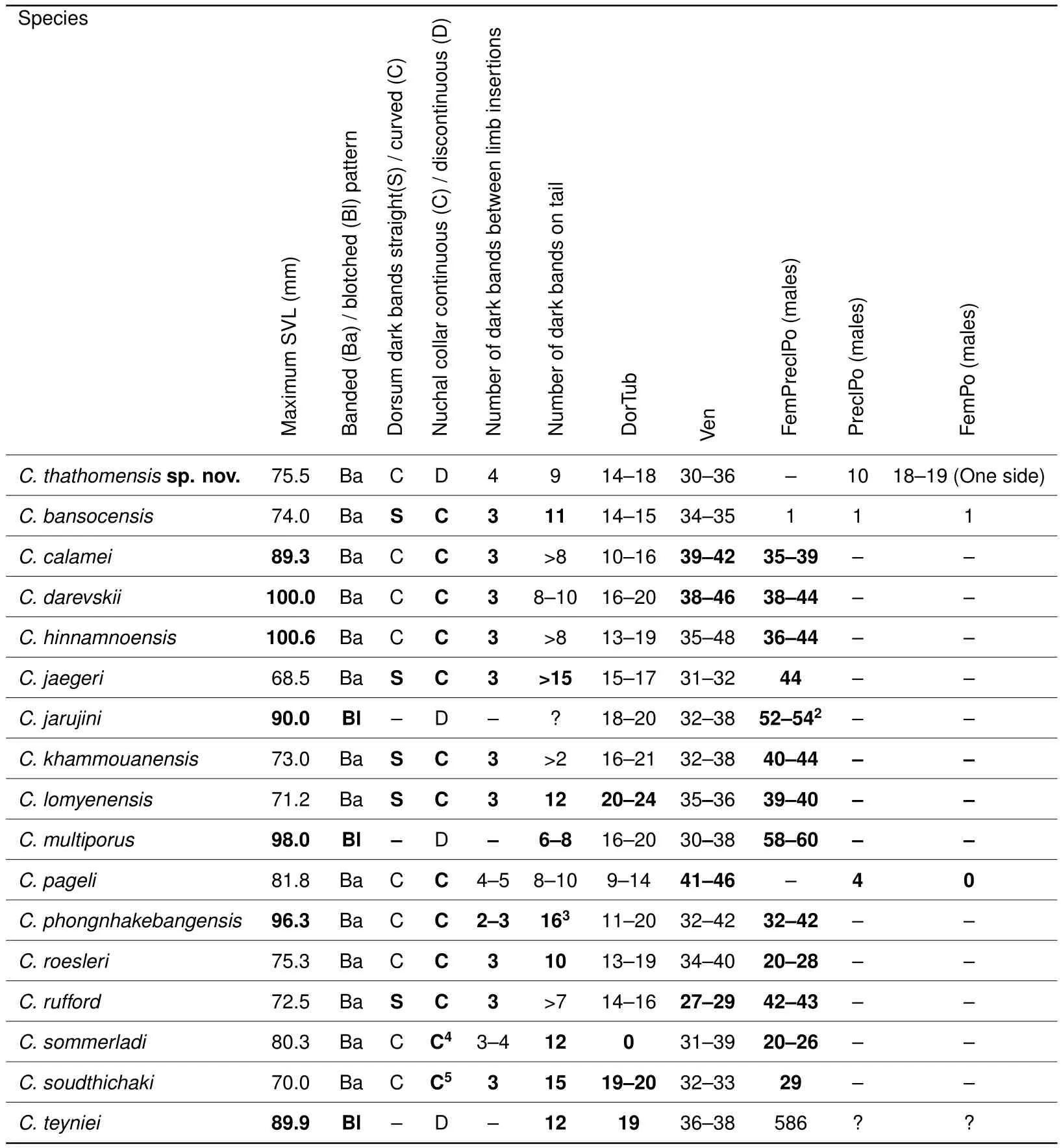
Table 4 Main diagnostic characters of species included in the Cyrtodactylus phongnhakebangensis species group sensu Luu et al.(2016a)
As far as we know,Cyrtodactylus thathomensissp.nov.is not found in the pet trade nor used in traditional medicine.The type-locality not being located within a protected area,the main potential threats to this new gecko species are habitat destruction through deforestation and limestone exploitation.To date,however,there appears to be no immediate concern as to the conservation status of this species,despite its limited distribution.Our new discovery stresses again the necessity to systematically survey karst massifs to inventory their unique,often micro-endemic and fragile,biodiversity.
COMPETING INTERESTS
The authors declare that they have no competing interests.
AUTHORS’CONTRIBUTIONS
R.A.N.and N.A.P.designed the study.E.L.K.,A.S.C.and N.L.O.collected data. N.A.P.performed molecular experiments. R.A.N.and O.S.G.P.examined morphology.N.A.P.conducted phylogenetic analyses.O.S.G.P.,R.A.N.and N.A.P.wrote the manuscript,N.L.O.and E.L.K.revised the manuscript.All authors read and approved the final manuscript.
ACKNOWLEDGEMENTS
We are very grateful to Assoc.Prof.Houngphet Chanthavong(Faculty of Forestry,National University of Laos)for organizing fieldwork and permitting specimen collection;work in Laos was carried out within the frameworks of the MOU on the Academic and Scientific Cooperation between the National University of Laos and Kaluga State University,Russia(MOU No. 72 20-11-2012).Th export of collected specimens was allowed due to export permits issued by the Ministry of Agriculture and Forestry of Lao PDR,Department of Forestry(certificate 511 No.271/08)and signed by the CITES Management Authority of Lao PDR.We thank Georges Lenglet(IRSNB),Valentina Orlova,Evgeniy Dunayev(Zoological Museum,Moscow State University),and Kumthorn Thirakhupt(CUMZ)for providing working facilities.We are grateful to the four anonymous reviewers for useful comments on the earlier version of this manuscript.RAN thanks Evgenya Lazareva for help in preparing illustrations for this work.N.A.P.thanks Duong Van Tang for assistance in the lab.
REFERENCES
Bauer AM.2003.Descriptions of seven newCyrtodactylus(Squamata:Gekkonidae)with a key to the species of Myanmar(Burma).Proceedings of the California Academy of Sciences,54(25):463-498.
Brennan IG,Bauer AM,Ngo VT,Wang YY,Wang WZ,Zhang YP,Murphy RW.2017.Barcoding utility in a mega-diverse,cross-continental genus:keeping pace withCyrtodactylusgeckos.Scientific Reports,7:5592.
Connette GM,Oswald P,Thura MK,LaJeunesse Connette KJ,Grindley ME,Songer M,Zug GR,Mulcahy DG.2017.Rapid forest clearing in a Myanmar proposed national park threatens two newly discovered species of geckos(Gekkonidae:Cyrtodactylus).PLoS One,12(4):e0174432.
David P,Nguyen TQ,Schneider N,Ziegler T.2011.A new species of the genusCyrtodactylusGray,1827 from central Laos(Squamata:Gekkonidae).Zootaxa,2833:29-40.
Ellis M,Pauwels OSG.2012.The bent-toed geckos(Cyrtodactylus)of the caves and karst of Thailand.Cave and Karst Science,39(1):16-22.
Felsenstein J.2004.Inferring Phylogenies.Sunderland:Sinauer Associates.Grismer LL,Wood Jr PL,Ngo VT,Murdoch ML.2015.The systematics and independent evolution of cave ecomorphology in distantly related clades of Bent-toed Geckos(genusCyrtodactylusGray,1827)from the Mekong Delta and islands in the Gulf of Thailand.Zootaxa,3980(1):106-126.
Grismer LL,Wood Jr PL,Quah ESH,Anuar S,Muin MA,Sumontha M,Ahmad N,Bauer AM,Wangkulangkul S,Grismer JL,Pauwels OSG.2012.A phylogeny and taxonomy of the Thai-Malay Peninsula Bent-toed Geckos of theCyrtodactylus pulchelluscomplex(Squamata:Gekkonidae):combined morphological and molecular analyses with descriptions of seven new species.Zootaxa,3520:1-55.
Hall TA.1999.BioEdit:a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT.Nuclear Acids Symposium Series,41:95-98.
Hartmann T,Geissler P,Poyarkov Jr NA,Ihlow F,Galoyan EA,Rödder D,Böhme W.2013.A new species of the genusCalotesCuvier,1817(Squamata:Agamidae)from southern Vietnam.Zootaxa,3599(3):246-260.Hillis DM,Moritz C,Mable BK.1996.Molecular Systematics.2nded.Sunderland:Sinauer Associates.
Huelsenbeck JP,Hillis DM.1993.Success of phylogenetic methods in the four-taxon case.Systematic Biology,42(3):247-264.
Huelsenbeck JP,Ronquist F.2001.MRBAYES:Bayesian inference of phylogenetic trees.Bioinformatics,17(8):754-755.
Ivanova NV,deWaard JR,HebertPDN.2006.An inexpensive,automation-friendly protocol for recovering high-quality DNA.Molecular Ecology Notes,6(4):998-1002.
Jobb G,von Haeseler A,Strimmer K.2004.TREEFINDER:a powerful graphicalanalysis environmentfor molecularphylogenetics.BMC Evolutionary Biology,4:18.
Le DT,Nguyen TQ,Le MD,Ziegler T.2016.A new species ofCyrtodactylus(Squamata:Gekkonidae)from Ninh Binh Province,Vietnam.Zootaxa,4162(2):268-282.
Luu VQ,Bonkowski M,Nguyen TQ,Le MD,Schneider N,Ngo HT,Ziegler T.2016a.Evolution in karst massifs:cryptic diversity among bent-toed geckos along the Truong Son Range with descriptions of three new species and one new country record from Laos.Zootaxa,4107(2):101-140.
Luu VQ,Calame T,Nguyen TQ,Le MD,Bonkowski M,Ziegler T.2016b.Cyrtodactylus rufford,a new cave-dwelling bent-toed gecko(Squamata:Gekkonidae)from Khammouane Province,central Laos.Zootaxa,4067(2):185-199.
Luu VQ,Nguyen TQ,Le MD,Bonkowski M,Ziegler T.2016c.A new species of karst-dwelling bent-toed gecko(Squamata:Gekkonidae)from Khammouane Province,central Laos.Zootaxa,4079(1):87-102.
Mahony S.2009.Taxonomic status ofCyrtodactylus khasiensis tamaiensis(Smith,1940)and description of a new species allied toC.chrysopylosBauer,2003 from Myanmar(Reptilia:Gekkonidae).Hamadryad,34(1):62-74.
MahonyS,AhmedM,HossainMK,KabirMM,HasanMK.2009.Cyrtodactylus ayeyarwadyensisBauer,2003(Squamata:Gekkonidae)in Bangladesh with habitat details of new collection localities and a discussion of morphological variation.Salamandra,45(4):245-250.
Murphy RW,Crawford AJ,Bauer AM,Che J,Donnellan SC,Fritz U,Haddad CFB,Nagy ZT,Poyarkov NA,Vences M,Wang WZ,Zhang YP.2013.Cold Code:the global initiative to DNA barcode amphibians and nonavian reptiles.Molecular Ecology Resources,13(2):161-167.
Nagy ZT,Sonet G,Glaw F,Vences M.2012.First large-scale DNA barcoding assessment of reptiles in the biodiversity hotspot of Madagascar,based on newly designed COI primers.PLoS One,7(3):e34506.
Nazarov RA,Poyarkov NA,Orlov NL,Phung TM,Nguyen TT,Hoang DM,Ziegler T.2012.Two new cryptic species of theCyrtodactylusirregulariscomplex(Squamata:Gekkonidae)from southern Vietnam.Zootaxa,3302:1-24.
Nazarov RA,Poyarkov Jr NA,Orlov NL,Nguyen NS,Milto KD,Martynov AA,Konstantinov EL,Chulisov AS.2014.A review of genusCyrtodactylus(Reptilia:Sauria:Gekkonidae)in fauna of Laos with description of four new species.Proceedings of the Zoological Institute RAS,318(4):391-423.
Ngo VT.2011.Cyrtodactylusmartini, anothernew karst-dwellingCyrtodactylusGray,1827(Squamata:Gekkonidae)from Northwestern Vietnam.Zootaxa,2834:33-46.
Ngo VT,Pauwels OSG.2010.A new cave-dwelling species ofCyrtodactylusGray,1827(Squamata:Gekkonidae)from Khammouane Province,southern Laos.Zootaxa,2730:44-56.
Nguyen SN,Yang JX,Le TNT,Nguyen LT,Orlov NL,Hoang CV,Nguyen TQ,Jin JQ,Rao DQ,Hoang TN,Che J,Murphy RW,Zhang YP.2014.DNA barcoding of Vietnamese bent-toed geckos(Squamata:Gekkonidae:Cyrtodactylus)and the description of a new species.Zootaxa,3784(1):48-66.
Nguyen SN,Le TNT,Tran TAD,Orlov NL,Lathrop A,MacCulloch RD,Le TDT,Jin JQ,Nguyen LT,Nguyen TT,Hoang DD,Che J,Murphy RW,Zhang YP.2013.Phylogeny of theCyrtodactylus irregularisspecies complex(Squamata:Gekkonidae)from Vietnam with the description of two new species.Zootaxa,3737(4):399-414.
Nguyen SN,Zhou WW,Le TNT,Tran ADT,Jin JQ,Vo BD,Nguyen LT,Nguyen TT,Nguyen TQ,Hoang DD,Orlov NL,Che J,Murphy R,Zhang YP.2017.Cytonuclear discordance,cryptic diversity,complex histories,and conservation needs in Vietnamese Bent-toed Geckos of theCyrtodactylus irregularis species complex.Russian Journal of Herpetology,24(2):133-154.
Panitvong N,Sumontha M,Tunprasert J,Pauwels OSG.2014.Cyrtodactylus saiyoksp.nov.,a new dry evergreen forest-dwelling Bent-toed Gecko(Squamata:Gekkonidae)from Kanchanaburi Province,western Thailand.Zootaxa,3869(1):64-74.
Pauwels OSG,Sumontha M.2014.Cyrtodactylus samroiyot,a new limestone-dwellingBent-Toed Gecko(Squamata: Gekkonidae)from Prachuap Khiri Khan Province,peninsular Thailand.Zootaxa,3755(6):573-583.
Pauwels OSG,Sumontha M,Bauer AM.2016.A new bent-toed gecko(Squamata: Gekkonidae:Cyrtodactylus)from Phetchaburi Province,Thailand.Zootaxa,4088(3):409-419.
Pauwels OSG,Sumontha M,Panitvong N,Varaguttanonda V.2014.Cyrtodactyluskhelangensis,a new cave-dwelling Bent-toed Gecko(Squamata: Gekkonidae)from Lampang Province,northern Thailand.Zootaxa,3755(6):584-594.
Posada D,Crandall KA.1998.MODELTEST:testing the model of DNA substitution.Bioinformatics,14(9):817-818.
Ronquist F,Huelsenbeck JP.2003.MRBAYES 3:Bayesian phylogenetic inference under mixed models.Bioinformatics,19(12):1572-1574.
Sambrook J,Fritsch EF,Maniatis T.1989.Molecular Cloning:A Laboratory Manual.Cold Spring Harbor:Cold Spring Harbor Laboratory Press.
Simmons JE.2002.Herpetological collecting and collections management.Revised ed.Herpetological Circular No.31.Salt Lake City:Society for the Study of Amphibians and Reptiles,1-153.
Smith MA,Poyarkov Jr NA,Hebert PDN.2008.CO1 DNA barcoding amphibians:take the chance,meet the challenge.Molecular Ecology Notes,8(2):235-246.
Solovyeva EN,Poyarkov NA,Dunaev EA,Duysebayeva TN,Bannikova AA.2011.Molecular differentiation and taxonomy of the sunwatcher toad headed agama species complexPhrynocephalussuperspecieshelioscopus(Pallas 1771)(Reptilia:Agamidae).Russian Journal of Genetics,47(7):842-856.Sumontha M,Pauwels OSG,Suwannakarn N,Nutatheera T,Sodob W.2014.Cyrtodactylus wangkulangkulae(Squamata:Gekkonidae),a new bent-toed gecko from Satun Province,southern Thailand.Zootaxa,3821(1):116-124.Sumontha M,Pauwels OSG,Panitvong N,Kunya K,Grismer LL.2015.A new lowland forest bent-toed gecko(Squamata:Gekkonidae:Cyrtodactylus)from Ranong Province,peninsular Thailand.Zootaxa,3911(1):106-118.
Tamura K,Stecher G,Peterson D,Filipski A,Kumar S.2013.MEGA6:molecular evolutionary genetics analysis version 6.0.Molecular Biology and Evolution,30(12):2725-2729.
Teynié A,David P.2014.Amphibiens et reptiles des formations karstiques du Laos.Bulletin de la Société Herpétologique de France,148:451-489.
Thompson JD,Gibson TJ,Plewniak F,Jeanmougin F,Higgins DG.1997.The Clustal_X windows interface: fl exible strategies for multiple sequence alignment aided by quality analysis tools.Nucleic Acids Research,25(24):4876-4882.
Uetz P, Freed P, Hošek J. 2018. The reptile database.http://www.reptile-database.org.
Ulber T.1993.Bemerkungen über cyrtodactyline Geckos aus Thailand nebst Beschreibungen von zwei neuen Arten(Reptilia:Gekkonidae).Mitteilungen aus dem Zoologischen Museum in Berlin,69(2):187-200.
APPENDIX I
List of examined specimens(Distr.=District,Prov.=Province).
Cyrtodactylus bidoupimontis:ZMMU R-13368(holotype),northern slope of Bidoup Mountain,near Klong Klanh village,Bidoup-Nui Ba National Park,Da Chais commune,Lac Duong Distr.,Lam Dong Prov.,Vietnam;ZMMU R-13369-1-5(paratypes),along the road to Nha Trang,6-7 km from Klong Klanh,Da Nhim river valley in Bidoup-Nui Ba National Park,Lam Dong Prov.,Vietnam;additional specimens listed in Nazarov et al.(2012).
Cyrtodactylus brevipalmatus:THNHM 3121-3123,THNHM 3125,Khao Phanoen Thung Camp,Kaeng Krachan National Park,Kaeng Krachan Distr.,Phetchaburi Prov.,Thailand.
Cyrtodactylus bugiamapensis:ZISP 26323(paratype),ZMMU R-13366(holotype),ZMMU R-13367-1-4(paratypes),Bu Gia Map National Park,Dak Ka stream valley,Bu Gia Map commune,Bu Gia Map Distr.,Binh Phuoc Prov.,Vietnam;additional specimens listed in Nazarov et al.(2012).
Cyrtodactylus chanhomeae:CUMZ R 2003.62(paratype),Thep Nimit Cave,Khun Khlon Subdistrict,Phraputthabata Distr.,Saraburi Prov.,Thailand;IRSNB 2585(holotype),Phraya Chat-tan Cave,Khun Khlon Subdistrict,Phraputthabata Distr.,Saraburi Prov.,Thailand.
Cyrtodactylusdoisuthep: CUMZR 0.2318(paratype),QSMI1168(paratype),THNHM 22543(holotype),Doi Suthep,Muang Distr.,Chiang Mai Prov.,Thailand.
Cyrtodactylus dumnuii:CUMZ R 2009-6-24-5-6(paratypes),KZM 002(paratype),THNHM 15904(holotype),THNHM 15905(paratype),Tham(Cave)Phabartmaejon,Ban Thakilek,Mae-Na Subdistrict,Chiang Dao Distr.,Chiang Mai Prov.,Thailand.
Cyrtodactylus erythrops:THNHM 15377(holotype),Tham Lod,Pangmapha Distr.,Mae Hong Son Prov.,Thailand.
Cyrtodactylus intermedius:IRSNB 17011,Nakhon Ratchasima,Thailand;ZMMU R-11213-1,Phnom Bakor National Park,Cambodia.
Cyrtodactylus inthanon: CUMZ R 0.2320(paratype),THNHM 22550(holotype),THNHM 25605(paratype),Doi Inthanon,Jom Thong Distr.,Chiang Mai Prov.,Thailand.
Cyrtodactylus khelangensis:CUMZ R 0.2318(paratype),PSUZC-RT 2012.9(paratype),THNHM 22548(holotype),limestone cave in Pratu Pha,Mae Mo Distr.,Lampang Prov.,Thailand.
Cyrtodactylus kunyai:THNHM 2560(holotype),limestone cave in Suan Hin Pha Ngam,Nong Hin Distr.,Loei Prov.,Thailand.
Cyrtodactylus lekaguli:IRSNB 2678(paratype),MNHN 1998.0600,Tham Phung Chang,Phang-Nga City,behind Provincial Hall,Muang Distr.,Phang-Nga Prov.,Thailand;MNHN 1999.7706,Tham Reusi,Muang Distr.,Phang-Nga Prov.,Thailand.Cyrtodactyluscf.lekaguli:IRSNB 16558,Nakhon Si Thammarat Prov.,Thailand.
Cyrtodactylus oldhami:MNHN 1999.7626,MNHN 1999.7698,Phang-Nga Wildlife Breeding Station,Muang Distr.,Phang-Nga Prov.,Thailand.
Cyrtodactylus peguensis peguensis:MNHN 1998.0544,Ban Kok Jaroen,Tap-Phut Distr.,Phang-Nga Prov.,Thailand.
Cyrtodactylus phetchaburiensis:IRSNB 2682(holotype),Tham Khao Tomo,Khao Tomo,Klatluang Subdistrict,Tha Yang Distr.,Phetchaburi Prov.,Thailand;IRSNB 2683(paratype),Phetchaburi Prov.,Thailand.
Cyrtodactylus phuketensis:PSUZC-RT 2010.58(holotype),QSMI 1170(paratype),THNHM 15378(paratype),Ban Bangrong,Thalang Distr.,Phuket Island,Phuket Prov.,Thailand.
Cyrtodactylus ranongensis:PSUZC-RT 2012.7-8(paratypes),THNHM 22545(holotype),Ban Ton Kloy,Kampuan Subdistrict,Suk Samran Distr.,Ranong Prov.,Thailand;THNHM 22546(paratype),along a street near Phetchkasem Road,Ban Kampuan,Kampuan Subdistrict,Suk Samran Distr.,Ranong Prov.,Thailand.
Cyrtodactylussaiyok: THNHM25602(holotype),CUMZR0.2320(paratype),Moo 3,Wang Krajae Subdistrict,Sai Yok Distr.,Kanchanaburi Prov.,Thailand.
Cyrtodactylussamroiyot: CUMZR 0.2320(paratype),QSMI1167(paratype),THNHM 22549(holotype),Ban Bang Pu,Sam Roi Yot Distr.,Prachuap Khiri Khan Prov.,Thailand.
Cyrtodactylussanook: QSMI1165(paratype),PSUZC-RT 2012.4(paratype),THNHM 22541(holotype),Wat Tham Sanook,Banna Subdistrict,Muang Distr.,Chumphon Prov.,Thailand.
Cyrtodactylus sumonthai:CAS 223819(paratype),CUMZ R 2002.2.5.1(paratype),IRSNB 2625(paratype),Khao Wong,Rayong Prov.,Thailand;IRSNB 2626(paratype),Tham Tao,Khao Wong,Rayong Prov.,Thailand;IRSNB 2624(holotype),Tham Khang Khao,Khao Wong,Rayong Prov.,Thailand.
Cyrtodactylus thirakhupti:CUMZ R 2003.120(holotype),CUMZ R 2003.121(paratype),IRSNB 2590(paratype),Tham Khao Sonk,Thachana Distr.,Surat Thani Prov.,Thailand.
Cyrtodactylus tigroides:CUMZ R 2002.296C(holotype),IRSNB 2586(paratype),Ban Tha Sao,Sai Yok Distr.,Kanchanaburi Prov.,Thailand.
Cyrtodactylus wangkulangkulae:THNHM 22547,limestone cave near Wangsaithong Waterfall,Manang Distr.,Satun Prov.,Thailand.
APPENDIX II
General distribution of the genusCyrtodactylusin Laos and surrounding areas,legend to Figure 1(Prov.=Province).
Localities:1-Cyrtodactyluscf.martini-Xishuangbanna,Yunnan Prov.,China;2-C.martini-Lai Chau,Lai Chau Prov.,Vietnam(type locality);3-C.bichnganae-Son La,Son La Prov.,Vietnam(type locality);4-C.wayakonei-Kao Rao Cave,Ban Nam Eng,Vieng Phoukha,Luang Nam Tha Prov.,Laos(type locality);5-C.otai-Hang Kia,Hang Kia-Pa Co N.R.,Mai Chau District,Hoa Binh Prov.,Vietnam(type locality);6-C.bobrovi-Hau 3,Ngoc Lau Commune,Ngoc Son-Ngo Luong N.R.,Lac Son District,Hoa Binh Prov.,Vietnam(type locality);7-C.huongsonensis-Huong Son,My Duc,Hanoi,Vietnam(type locality);8-C.cucphuongensis-Cuc Phuong N.P.,Nho Quan,Ninh Binh Prov.,Vietnam(type locality);9-C.soni-Da Han,Van Long Wetland N.R.,Gia Vien District,Ninh Binh Prov.,Vietnam(type locality);10-C.puhuensis-Pu Hu,Thanh Hoa Prov.,Vietnam(type locality);11-C.cf.puhuensis-Houphan Prov.,Laos;12-C.vilaphongi-Ban Xieng Muak,Luang Prabang,Luang Prabang Prov.,Laos(type locality);13-C.chauquangensis-Chau Quang,Quy Hop,Nghe An Prov.,Vietnam(type locality);14-C.cf.interdigitalisandC.spelaeus-Khuang Lang N.P.,Kasi,Vientiane Prov.,Laos(type locality ofC.spelaeus);15-C.pageli-Phoukham Cave,Ban Na Thong,Vang Vieng,Vientiane Prov.,Laos(type locality);16-Cyrtodactylus thathomensissp.nov.-Ban Thathom,Xiangkhouang Prov.,Laos(type locality);17-C.cf.jarujini-Phou Khao Khouay N.P.,Bolikhamxay Prov.,Laos;18-C.interdigitalis-Nam Lik River Valley,Ban That Wang Monh,Vientiane Prov.,Laos;19-C.teyniei-Ban Na Hin(Nahin),Nam Kading NBCA,Bolikhamxay Prov.,Laos(type locality);20-C.jarujini-Phu Wua W.S.,Nong Dern,Bung Kan,Nong Khai Prov.,Thailand(type locality);21-C.cf.teyniei-Nahin,Khammouane Prov.,Laos;22-C.cf.roesleri-Phou Hin Boun N.P.,Konglor,Khammouane Prov.,Laos;23-C.soudthichaki-Khun Don,Phou Hin Poun N.P.,Khammouane Prov.,Laos(type locality);24-C.jaegeri-Thakhek,Khammouane Prov.,Laos(type locality);25-C.lomyenensis-Lomyen Cave,Gnommalath,Khammouane Prov.,Laos(type locality);26-C.rufford-Nang Log cave,Gnommalath District,Khammouane Prov.,Laos(type locality);27-C.darevskii,C.khammouanensisandC.multiporus-Na Home,Boulapha,Khammouane Prov.,Laos(type locality);28-C.bansocensis-Peopalam Cave,Ban Soc,Bualapha District,Khammouane Prov.,Laos(type locality);29-C.calamei,C.hinnamnoensis,C.sommerladi,C.cryptus-Hin Nam No N.P.,Khammouane Prov.,Laos(type locality forC.calamei,C.hinnamnoensisandC.sommerladi);30-C.roesleri-Ke Go N.R.,Ha Tinh-Quang Binh provincial border,Vietnam;31-C.phongnhakebangensis,C.cryptusandC.roesleri-Phong Nha-Ke Bang N.P.,Minh Hoa,Quang Binh Prov.,Vietnam(type locality);32-C.kunyai-Suan Hin Pha Ngam,Nong Hin District,Loei Prov.,Thailand(type locality);33-C.interdigitalis-Tham Yai Nam Nao,Nam Nao N.P.,Phetchabun Prov.,Thailand(type locality);34-C.pseudoquadrivirgatus-Huong Hoa,Quang Tri Prov.,Vietnam;35-C.pseudoquadrivirgatus-A Luoi,Thua Thien-Hue Prov.,Vietnam(type locality);36-Cyrtodactylussp.1-Ba Na,Da Nang,Vietnam;37-C.buchardi-Kiatngong,Xepian NBCA,Champasak Prov.,Laos;38-C.taynguyenensis-Kon Plong,Kon Tum Prov.,Vietnam;39-C.taynguyenensis-Krong Pa(type locality),Kon Ka Kinh N.P.and Kon Chu Rang N.R.,Gia Lai Prov.,Vietnam.
杂志排行

Zoological Research的其它文章
- A new genus and three new species of miniaturized microhylid frogs from Indochina(Amphibia:Anura:Microhylidae:Asterophryinae)
- A new species of the genus Theloderma Tschudi,1838(Amphibia:Anura:Rhacophoridae)from Tay Nguyen Plateau,central Vietnam
- A new species of the genus Leptolalax(Anura:Megophryidae)from southern Vietnam
- A new species of smooth skink(Squamata:Scincidae:Scincella)from Cambodia