山西沁河上游植物种间关系及功能群划分
2018-06-19田益瑆郭东罡
田益瑆,郭东罡
(山西大学环境与资源学院,山西 太原 030006)
沁河属于黄河一级支流,是黄河三门峡至花园口区间两大支流之一,全长为456 km,流域面积为12 900 km2,包括山西省和河南省16个县(区),流域内煤炭资源丰富,人类活动历时悠久。沁河发源于山西省太岳山山区,上游流经县域均为我国的产煤大县,其生态系统服务功能尤为重要,但是关于沁河上游湿地植被群落生态学的系统研究并不多见。
本研究在样方调查的基础上,采用χ2检验、联结系数(AC)和共同出现百分率(PC),研究了沁河上游湿地植物群落的种间关联性,并对种间相关的测定结果进行了生态分析[1-3],旨在为保护沁河源头水环境质量、湿地生物多样性和科学评价当地生态本底等提供科学数据。
1 研究区域概况
沁河上游主要位于山西省长治市境内,长度为86.40km,处于东经 112°3′12.5552″~112°24′46.4175″,北纬 36°22′42.7105″~36°52′28.2353″。属典型的暖温带大陆性季风气候,年均气温5.5~7.8℃,1月份平均气温 -6.8~-10.4℃,7月份平均气温17.4~20.9℃;通常的无霜期是90~150 d,年均降水量656.7 mm;年平均日照时数2 519 h。土壤主要为冲积土、沼泽土和草甸土等。
2 研究方法
2.1 样地设置与调查
在踏查的基础上,采用系统随机取样法,在沁河上游沿岸,从源头和主要水源补给区顺流而下,尽量避开人为干扰,在河岸两侧设置了7块样地,在每块样地内,平行于河岸设置6条1 m×10 m样线,河岸两侧各设置3条,每条样线间隔5 m,在每条样线上取10个1 m×1 m的样方,共计样方数420个。在每个样方内记录植物种名、盖度、高度、多度、物候期、生活型和生活力等,同时记录相关环境因子情况[4]。各样地基本概况列于表1。

表1 样地基本概况
2.2 数据处理
种间联结性测定,χ2检验是运用二元定性数据测定种间联结,将40个高频种和420个样方组成40×420的种-样方矩阵,以这些数据为基础,得出这40个高频种780个种对应的2×2列联表,再以此表为基础对所列出的种对之间的种间关联系数进行计算,即对a,b,c与d值进行计算。然后利用χ2检验,与AC和PC相结合对物种间联结性进行测定。
2.2.1 常用的用于确定物种间是否存在关联关系的χ2检验方法 依据所列出的联表中的χ2统计量,通过如下公式进行检验。

其中,n指的是样方总数,a指的是2个物种均出现的样方数量,b指的是A物种出现而B物种不出现的样方数量,c指的是B物种出现而A物种不出现的样方数量,d指的是A物种和B物种都不出现的样方数量。当ad>bc时为正联结,当ad<bc时为负联结。当χ2>χ20.05(1)(此时χ2对应的自由度是1,显著性P对应的值为0.05)说明种间的关联性显著;χ2>χ20.01(1)(此时 χ2的值对应的自由度为1,显著性P值为0.01)则说明是极显著的,反之则是不显著。χ20.05(1)=3.841,χ20.01(1)=6.635,即当3.841<χ2<6.635 为关联显著,χ2>6.635 为极显著,χ2<3.841为关联不显著。
2.2.2 联结系数AC AC系数可以用来对χ2检验结果进行检验,并表示种间联结程度。
若 bc>ad,且 d≥a,则有下式。

若 bc>ad,且 d<a,则有下式。

若ad≥bc,则有下式。
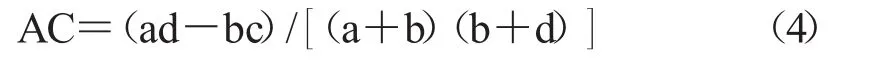
AC的值域为[-1,1]。当AC所对应的值越趋近于1的情况下,说明对应的物种间的正联结性越强;反之,AC值更加趋近于-1,则说明物种间的负联结性越强;当AC值为0时,物种间完全独立。
2.2.3 共同出现百分率PC PC值可以表示物种间的正联结程度。

式中,PC的值域为[0,1]。当PC值越靠近1时,说明物种之间的正联结程度越强[5-7]。
3 结果与分析
3.1 群落组成
本研究选取了研究区域内频度>5%的物种为高频草本物种,结果显示,共有40个高频种,属于15科35属,其中,水生植物与湿生植物有8种,中生植物与旱生植物有32种。主要高频种为狗尾草(Setaria viridis)、节节草(Commelina diffusa)、棒头草(Polypogon fugax)、青蒿(Artemisia carvifolia)、碱茅(Puccinellia distans)、早开堇菜(Viola prionantha)、芦苇(Phragmites australis)、花苜蓿(Medicago ruthenica)、车前(Plantago asiatica)、旋覆花(Inula japonica)等。
3.2 种间联结测定的χ2值、AC值和PC值特征分析
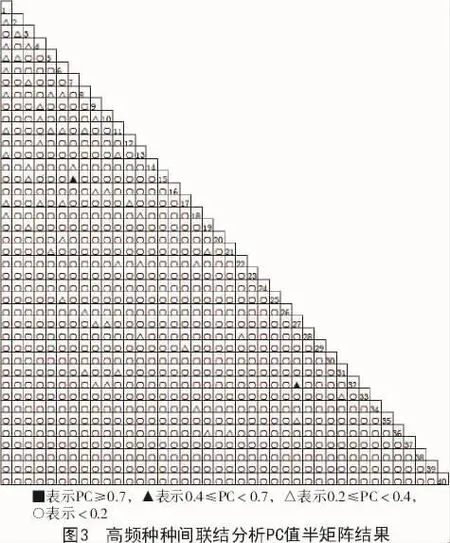
根据表2以及图1,2,3可知,由于所测指标性质不同,χ2,AC值和PC值3个指数有所差异。χ2能够体现种间联结的显著状况,但它只能体现种间联结程度是否显著,对于种间联结程度不显著的种对,不能准确体现种对间的联结程度。AC和PC更能够客观地对通过χ2检验结果为不显著联结种对间所对应的联结程度进行检验[7-9]。联结系数AC和共同出现百分率PC虽然能反映种对间联结程度强弱,但缺乏统一的标准划分种间联结程度。假若物种出现频率低,特别是在a值是0的时候,AC和PC对应的值对物种间关联程度有放大作用,甚至有时候会得到相反的结果。如果葎草、柳叶菜、地榆、荆三棱等物种的出现频率较低,次数较少,这些物种和其他高频种一起出现的样方数的a值等于0的概率较大。极显著负关联(AC<0.60)高达265对,正是这种原因所造成的。当物种出现次数较多时,共同出现百分率PC值能够避免由于2个物种均未出现的样方数d偏高而造成联结系数AC值偏高的问题,但PC所对应的值只能对种对之间的正联结程度做出较为准确的反应。发生此种情况的原因是由于在a值比较小的情况下,会对d值进行忽略,对a,b,c的作用值进行夸大,同时夸大了负联结程度。这种现象在出现次数较少物种出现时更为明显。即物种的分布较为均匀时,AC和PC的数值是准确的,而在沁河上游河岸带生态系统里,由于受到人为或自然因素的影响,河岸带生态系统受到破坏,物种的分布均匀度较差[10-12]。
总之,选择40个草本高频种。经过χ2检验的同时,把AC和PC所对应的值作为用于检验的辅助参数,对沁河河岸带生态系统里面存在的主要高频种种间存在的关联性进行分析[13-14]。

表2 χ2检验值、AC值、PC值
3.3 种间联结性

χ2检验结果说明,在全部的40个草本高频种所进行组成的780个种对中,表现为正联结一共有319对,占总量的40.90%,表现为负联结的有458对,占总量的58.72%,表现为无关联的种对为3对,占总种对数的0.38%。正负联结种对数之比为0.70∶1,由此可知,沁河上游河岸带草本群落种对间正负联结差异较大,河岸带群落不够稳定,易于受到外部因素干扰。种对间呈显著联结以及呈极显著联结的种对数较少,种对间显著正联结以及极显著正联结的种对有9对,占总种对数的1.15%,种对间显著负联结以及极显著负联结的种对有1对,占总种对数的0.13%,表明沁河上游河岸带草本群落种对间联结性较差,种间的独立性强,生态位分离明显,在生长过程中,物种之间相互影响较小,容易受到人为或自然因素影响,河岸带草本群落稳定性差。呈极显著正联结的种对为:芦苇和荻,花苜蓿和拂子茅,车前和旋覆花,薄荷和荆三棱,矛叶荩草和红鳞扁莎等;呈显著正联结的种对为:车前和委陵菜,鼠掌老鹳草和披针薹草;呈极显著负联结的种对为狗尾草和芦苇;无关联种对为:多花胡枝子和苍耳,紫菀和柳叶菜,紫菀和葎草。
AC值结果表明,沁河上游河岸带草本群落中40个高频种种对间呈极显著正联结性(AC≥0.6)的种对有0对,显著正联结(0.2≤AC<0.6)种对有35对,占总对数的4.49%,不显著正联结(0.05≤AC<0.2)种对有147对,占总对数的18.85%;无联结(-0.05≤AC<0.05)的种对有159对,占总种对数的20.38%;不显著负联结(-0.2≤AC<-0.05)的种对有49对,占总对数的6.28%,显著负联结(-0.6≤AC<-0.2)的种对有125对,占总对数的16.3%,其中,表现为极显著负联结(AC<-0.6)的一共有265对,占33.97%。由此可知,该区域草本植物物种间联结性较弱,各物种偏向独立分布。
PC值结果表明,沁河上游河岸带草本群落中40个高频种种对间极显著正联结性(PC≥0.7)的种对有0对;显著正联结(0.4≤PC<0.7)种对有2对,占总对数的0.26%;不显著正联结(0.2≤PC<0.4)的种对有55对,占总对数的7.05%;无联结(PC<0.2)的种对有723对,占总种对数的92.69%。PC值可以对物种间的正联结存在的强弱关系进行较为准确地表示。
综上所述,种间联结测定的χ2值、AC值和PC值3种方法的结果均表明,沁河上游河岸带草本群落物种间正联结性弱,物种之间的相互影响程度不强,群落不够稳定,易受到人为或自然因素的干扰[15]。
3.4 功能群划分及分析
在群落内各个物种生态习性各不相同,种群之间的关联揭示了由于生境的影响不同群落的物种之间空间分布的关系[16-18]。本研究基于种间正联结性为基础、负联结性为界限以及草本高频种对水分因子的响应,对草本植物功能群进行了划分[19-20]。各功能群间的物种间存在正相关关系,这是由物种间的同一资源利用模式以及功能群内同类生态位所决定的[21]。功能群的划分不仅需要基于种间关系,还需要基于水分因子和其他环境因子[22-23]。

山西沁河上游草本植物群落的40个高频种可分为4个功能群:Ⅰ.水生植物功能群:包括芦苇、水芹以及荆三棱。该功能群物种对水分条件要求高,常见于地下水位较深、水分充足的环境中,广泛分布在沁河湿地的河道以及常年积水的沼泽等地区。Ⅱ.湿生植物功能群:包括旋覆花、薄荷、拂子茅、红鳞扁莎、柳叶菜。种间关系大多数为显著正关联以及正关联,生态习性相近。该功能群物种对水分条件要求较高,常见于地下水位较浅的地方,抗旱属性差,不能长时间在缺水环境下生长,广泛分布在沁河季节性积水的沼泽地和河漫滩等地区。Ⅲ.中生植物功能群:包括节节草、棒头草、青蒿、碱茅、野大豆、刺菜、蕨麻、紫菀、蒲公英、龙牙草以及地榆。该功能群物种主要分布在沁河河岸边以及沼泽地边等河滩一级阶地等潮湿地区。Ⅳ.中旱生植物功能群:包括狗尾草、早开堇菜、花苜蓿、车前、阿尔泰狗娃花、猪毛蒿、荻、委陵菜、鼠掌老鹳草、小花鬼针草、艾草、多花胡枝子、披碱草、矛叶荩草、隐子草、披针薹草、荩草、沙打旺、苍耳、多茎委陵菜以及葎草。该功能群物种主要分布在沁河河滩二级阶地以及湿地附近的路旁、林下、田边和河漫滩地势较高处地区。
通过功能群的划分可以分析得出,不同的功能群对生境具有不同的趋同性适应。而水分因子成为沁河上游湿地草本植物划分功能群的重要环境因子,起着决定性的作用。功能群的不同也反映了草本植物群落随着水分梯度的变化而产生明显的演替规律。
4 讨论与结论
利用高频种种间关系和水分条件划分植物功能群的方法,是对湿地生态系统功能群进行分类的一种尝试。高频种在生态系统中控制群落结构,能较好地反映一个区域的生态系统状况,因此,以高频种的选择为研究对象对植物功能群进行划分。沁河上游湿地在不同样地草本植物高频种的变化,揭示了高频种对水分条件变化的不同反应。例如,中旱生植物功能群广泛分布在湿地附近的路旁、林下、田边和河漫滩地势较高处地区,而水生植物功能群广泛分布在沁河湿地的河道以及常年积水的沼泽等地区。
对植物功能群的研究已经成为研究群落动态稳定性、植被分布格局、生态系统总体生产力的重要手段[12],所以,对于功能群划分的研究有着重要意义。湿地植物功能群的划分,将植物功能群中的物种作为一个整体研究其对水分因子以及外界干扰响应的规律,要比单个物种的研究更容易归纳出规律性变化[20]。功能群的划分要以种间关系为基础,结合其他因子,例如海拔[23]、物种生态习性等因子,具体划分方法要根据研究地区实际情况决定。
以高频种种间关系为基础,结合水分条件对沁河上游草本植物群落划分功能群符合生态学原理,符合当地实际生态现状。如果只是以物种间相关性为依据划分功能群,芦苇与荻为显著正相关,但2种物种并不同属一个功能群,芦苇常年生长在沁河河道以及常年积水的沼泽、滩涂,为水生植物功能群,而荻广泛分布于沁河湿地两旁的田边、林下等地势较高处,为中旱生植物功能群,二者生态习性大相径庭,并不能划分为同一个功能群。沁河上游湿地的草本植物在长期自然演替规律下,具有相近生态习性的物种会共同组成一个功能群,在同属一个功能群内物种间具有正联结性,经常结伴出现,对水分条件具有共同的反映趋势,生态位相近,在群落中相互利用、相互促进生长;而表现为负相关性的物种,对生境的反映趋势不同,很少共同出现,成为划分功能群的界限。在本研究中,沁河湿地水分生态功能群随着水分梯度的增加,土壤水分增加,水生植物与湿生植物的比例随之增加,在这种生境下,中旱生植物生长得到抑制,而水生植物与湿生植物的竞争力得到加强,因此,在群落中获得绝对优势;相反,水分条件减少,土壤水分降低,水分不充足时,耐旱的中旱生植物为优势物种。
沁河上游湿地草本植物物种间呈极显著以及显著相关的种对较少,整体物种间相关性较弱,这是由于所选取样地生境差异较大,有沁河湿地的河道以及常年积水的沼泽、季节性积水的沼泽地和河漫滩、沁河河岸边、湿地附近的路旁、林下、田边和河漫滩地势较高处等环境,而且物种生态学特性差异较大[13]。同时,40个高频种种对间正相关种对数小于负相关种对数。这是由于沁河上游附近农业的发展,人口的增长,沁河上游湿地附近地区成为人类活动区域,人为干扰因素增加,对湿地植物群落造成影响,使得植物群落不够稳定,景观破碎化加剧。沁河上游湿地生态系统虽然物种组成比较丰富,但也非常脆弱,人为因素以及自然因素的影响使得其生物、环境均受到一定程度的威胁。因此,应以沁河上游湿地草本植物功能群划分研究为基础,加强沁河上游的污染防治,并采取人工措施恢复该地区环境,使物种具有多样性,促进沁河上游湿地的健康发展[24]。
[1]KLEYER M.Validation of plant functional types across two contrasting landscapes[J].Journal of Vegetation Science,2002,13(2):167-178.
[2] SMITH T M,WOODWARD F I,SHUGART H H.Plant function types[M].Cambridge:Cambridge University Press,1997.
[3]李荣平,刘志民,蒋德明,等.植物功能型及其研究方法[J].生态学杂志,2004,23(1):102-106.
[4]贾慧君,李帅,郝婧,等.黄河中游(龙门-汾河入黄口)水分因子与湿地植物多样性的相关关系研究 [J].山西农业科学,2017,45(8):1325-1330.
[5]WOODWARDF I,CRAMER W.Plant functional types and climatic changes:introduction[J].Journal of Vegetation Science,1996,7(3):306-308.
[6]胡楠,范玉龙,丁圣彦,等.陆地生态系统植物功能群研究进展[J].生态学报,2008,28(7):3302-3311.
[7]郭屹立,卢训令,丁圣彦.伊洛河河岸带生态系统草本植物功能群划分[J].生态学报,2012,32(14):146-154.
[8] GRAFF P,AGUIAR M R,CHANETON E.Shifts in positive and negative plant interactions along a grazing intensity gradient[J].E-cology,2007,88:188-199.
[9]严慧玲,刘东明,王俊,等.河北邢汾高速公路沿线植物群落优势种的种间关系[J].河南大学学报(自然科学版),2015,45(4):76-83.
[10]王烨,秦浩,张峰,等.浊漳河干流湿地草本植物群落优势种种间关系及功能群划分 [J].生态学杂 志,2015,34(8):2109-2114.
[11]房飞,胡玉昆,张伟,等.高寒草原植物群落种间关系的数量分析[J].生态学报,2012,32(6):1898-1907.
[12]段后浪,赵安,姚忠.鄱阳湖区典型草洲植物种间关联性研究[J].湿地科学,2017,15(1):107-113.
[13]BROOKER R W,MAESTRE F T,CALLAWAY R M.Facilitation inplant communities:The past,the present,and the future[J].Journal of Ecology,2008,96:18-34.
[14]王建良,赵成章,张伟涛,等.秦王川湿地盐角草和盐地碱蓬种群的空间格局及其关联性 [J].生态学杂志,2017,36(9):2494-2500.
[15]刘亚琦,刘加珍,陈永金,等.孔雀河下游断流河道的环境特征及物种间关系[J].生态学报,2017,37(8):2706-2718.
[16]范晓,张峰,庞春花,等.山西汾河流域下游草本植物功能群[J].生态学杂志,2013,32(10):2716-2720.
[17]李潮,谢应忠,许冬梅,李侠.宁夏荒漠草原植物群落的种间关系[J].草业科学,2013,30(11):1801-1807.
[18]李兰兰,许诺,李洪远,等.七里海湿地植物种间关系的数量分析[J].水土保持通报,2014,34(4):70-75.
[19]程雷星,陈克龙,苏旭,等.青海湖流域小泊湖湿地植物种间关系研究[J].干旱区地理,2014,37(5):1005-1011.
[20]吴华,张建利,喻理飞,等.草海流域水源功能区植物持水功能群划分[J].水土保持研究,2014,21(2):138-143.
[21]刘珏宏,高慧,张丽红,等.内蒙古锡林郭勒草原大针茅-克氏针茅群落的种间相关特征分析 [J].植物生态学报,2010,34(9):1016-1024.
[22]秦晓娟,高璐,邓永利,等.山西平陆黄河湿地植功能群划分[J].山西大学学报(自然科学版),2014,37(3):454-460.
[23]赵艳云,刘京涛,陆兆华.渤海湾贝壳堤湿地芦苇种群与蒙古蒿种群空间分布格局和种间关系 [J].湿地科学,2017,15(2):187-193.
[24]王思元,牛萌.湿地系统的生态功能与湿地的生态恢复[J].山西农业科学,2009,37(7):55-57.
