西双版纳热带季雨林林冠层CO2浓度特征及其影响因素
2018-06-15巩在武
张 娜, 巩在武
(1.南京信息工程大学 法政学院, 南京 210044; 2.山西省阳泉市气象局, 山西 阳泉 045000)
陆地生态系统是人类赖以生存与持续发展的生命支持系统,而森林作为陆地生态系统的主体,在全球碳循环中起着极其重要的作用,同时森林生态系统的生产、呼吸等生态过程都受到温度的制约[1-2]。地温和土壤积温是一种重要的热量资源,它影响植物种子的萌发、生长发育、开花和结果,特别是对根系活动以及各种酶的活动影响极大。地温还制约着土壤中多种理化和生物作用的速率,从而影响植物生长[3-4]。林内地温和植物生长有直接关系,特别是对植物的蒸腾、根的呼吸、土壤微生物的活动、凋落物的分解以及土壤肥力等均有重要作用[5]。另外,土壤温度直接影响着土壤中微生物、地面凋落物及腐殖质的呼吸作用,对夜间生态系统净CO2交换有着重要的作用[6]。因此,地温是表征植物生产能力的重要参数之一,生态系统的光合、呼吸和生物量积累等碳循环过程都受温度的制约。
一般情况下,如果植物的生长所需水分能够达到满足,那么温度将会对其生长发育过程产生不可忽视的影响,在全球气温不断上升的背景下,土壤的分解作用将增强,且能够影响到土壤呼吸的季节变化,进而使得土壤的呼吸范围明显增大,促进了土壤中CO2的释放[7]。昼夜温差具有较大的变化,对于植物而言,其呼吸效应也不同,因此研究温度与生态呼吸之间的关系具有重要的现实意义,从而对生态系统的年呼吸量及季节过程进行相应估计,大量的研究表明,在温度适当上升的情况下,生态系统的呼吸指数会明显提升,但是对于特定的生态环境而言,环境指标的温度选择依然没有明确结论,在实际研究过程中气温、地温及其加权平均等指标都被选用过[8]。对于热带雨林而言,其具有多样化的作物群落分布,且层次性明显,这就导致其光照的获取存在显著差异,进而对植被下的地温产生间接的显著影响;此外,四季变换会导致植物茂盛程度不同甚至是落叶,这将导致地表获取光照的程度相差较大,从而导致不同的地温变化[9]。碳在生态循环中具有重要角色,而CO2通量是其中的重要一环,制约着生态系统中的光合作用及生物呼吸[10],土壤呼吸作用能够促进碳循环,而地温对其呼吸作用具有显著的制约作用,大量的研究表明二者具有显著正相关,因而对于森林地温的深入研究具有重要现实意义。
西双版纳地处北回归线以南,终年受西南季风控制,属热带季风气候,热带季节雨林是该地区重要的森林类型之一[11]。热带雨林是目前生态系统碳收支研究关注的重点,而土壤温度的变化在某种程度上影响着该区域的碳收支平衡,因而对地温的深入研究有利于正确回答该地区碳源汇的问题[12]。单从纬度来说,西双版纳很难形成大片的热带雨林,但是在该地区独特的气候条件、太阳高度及林窗边缘共同影响下,该地区形成了独特的热带季节雨林,并且林窗区域气温—地温差有明显的季节差异[13]。虽然目前对该地区的生物量、风、太阳辐射等有了较深入的研究,但对热带季节雨林地温变化规律的研究尚不多见。本文的观测数据由西双版纳热带季节雨林内铁塔上温度自记仪以及红外温度仪记录,2015年全年不间断观测获取,对热带季节雨林群落冠层植物温度及对应高度气温、冠层上、冠层下气温的日变化和季节变化特征进行分析,获得一些热带季节雨林冠层温度及冠层气温的分布规律。初步探讨热带季节雨林的太阳辐射特征、小气候特征、风时空变化特征和碳通量年变化特征及其影响因素,以期为深入研究热带雨林的植物更新与生长规律以及热带森林碳循环提供科学依据。
1 材料与方法
1.1 研究区概况
观测点位于云南省西双版纳州勐腊县境内的国家级自然保护区内(21°57′N,101°12′E,海拔756 m),距中国科学院西双版纳热带雨林生态系统定位研究站约10 km。本区位于热带的北缘,一年中有雾凉季(11月—翌年2月份)、干热季(3—4月份)、湿热季或雨季(5—10月)之分,太阳辐射年总量5 000~5 600 MJ/a,年均日照时数1 787.8 h,年均温21.4℃,最热月均温25.3℃,最冷月均温15.6 ℃。年均降雨量为1 557 mm,但分布不均,雨季降水占全年的83%左右。该地区独特的天气现象为雾日较多,年雾日可达(258±58)d,在雾凉季月平均雾日均超过23 d,最多的1月可达26.1 d;一般在夜间22∶30左右开始起雾,直到第2天12:00以后才逐渐消散;与雾凉季相比,干热季的雾生成时刻较迟,维持时间较短,一般在23:00之后才开始起雾,而在10:00左右就逐渐消散。
观测样地所在的热带季节雨林,林分平均高度35 m左右,郁闭度为0.9,结构复杂,分层现象明显,乔木层按高度即可分为3层,上层高30 m以上,藤本及附生植物丰富,板根及茎花现象显著。该雨林群落胸径≥10 cm的立木密度为每1 hm2386棵,每1 hm2基面积为30.03 m2,平均每木基面积为0.078 m2,平均树高为18.6 m。群落垂直结构复杂,分层现象明显。乔木层按高度可分为3 层:上层高30 m以上,优势种主要为千果榄仁(Terminaliamyriocarpa)和绒毛番龙眼(Pometiatomentosa);中层高度16~30 m,常见种有梭果玉蕊(Barringtoniafusicarpa)、白颜树(Gironnierasubaequalis)和山蕉(Mitrephoramaingayi)等;下层高16 m 以下,主要种包括云树(Garciniacowa)、假广子(Knemaerratica)、细罗伞(Ardisiasinoaustralis)、蚁花(Mezzettiopsiscreaghii)和毒鼠子(Dichapetalumgelonioides)等。灌木层除上层乔木的幼苗幼树外,常见种有染木树(Saprosmaternatum)、狭叶鸡眼藤(Morindaangustifolia)和玉叶金花(Mussaendasp.)等。草本层与灌木层处于同一层次,种类不多,常见有凤尾蕨(Pterissp.)、海芋(Alocasiamacrorrhiza)和沧江南星(Arisaemabonatianum)等,藤本及附生植物丰富,板根及茎花现象显著。
1.2 观测仪器设置
该区域全年季节差距较大,一般情况下,上年的11月至第二年的2月间称为雾凉季,3—4月称为干热季,雨季主要集中在5—10月,其中5—7月为雨季前期。开路红外气体分析仪安装在观测铁塔的48.8 m(林冠上方CO2浓度)和4.2 m(林内近地层CO2浓度)高处,测定林冠上方和林内近地层的CO2浓度和风速、风向,使用CR5000数据采集器按照10 Hz频率采集数据,并输出30 min的平均值。在70 m高处安装点状光量子探头用于测定光合有效辐射;同时在70 m高处安装雨量计测定降雨量,此两项数据采集时间间隔为30 min。温湿计(HMP45C,芬兰VAISALA公司)测定林冠上和林冠下的大气温度;由红外温度计(IRTS-P,美国APOGEE公司)测定冠层表面温度。每月中旬,使用LAI-2000在铁塔上分层进行测定。地表呼吸通量的测定则使用静态箱—气相色谱法,每周测定1次,每次3个处理,每个处理6个重复。土壤温度(0—40 cm)使用105型热电耦(Campbell Scientific Inc.,USA)测定,通过数据采集器控制采样时间间隔30 min。
与此同时,用LI-8150土壤CO2通量全自动连续测量系统(LI-COR,USA)全天监测土壤呼吸,在每个土壤环的测量时间为3 min,测量间隔为5 min,全天24 h不间断测量土壤呼吸,每个土壤环测定3次重复;由于每个土壤环每小时会被气室罩住约为3 min,而其他时间处于室外的开放状态,被隔绝时间较短,因此我们可以忽略降雨过程所此造成的误差。
1.3 数据处理
通过数据采集系统(CR5000,Campbell ScientificInc.,USA)采集数据,并通过程序控制每30 min自动输出各层CO2浓度的平均值,存储于站点的计算机内,用移动存储盘取回室内进行处理。由于仪器为开路,CO2浓度的测量往往受空气湿度的影响,造成部分数据不正常,必须对这类数据进行剔除;同时,也对时间变化曲线上相邻两个点之间CO2浓度相差大于50 mg/m3的数据进行剔除。对剔除后的数据进行如下的插补:对于缺失数据少于4个的情况(即数据缺失时间段小于2 h),使用线性内插;对于缺失数据缺失大于或等于4个的情况(即数据缺失大于或等于2 h),使用剔除后数据同一时刻的月平均值进行替换,完成插补。
2 结果与分析
2.1 热带季节雨林地温时空分布特征
由图1可知,各层地温均呈现正弦变化,愈靠近表层这个变化趋势愈明显,地温的最低值和最高值出现时刻随深度增加呈现滞后现象,显示了不同深度地温日变化的相位存在明显差异,但是随着深度的增加,其变幅急剧减小,到40 cm变幅已趋于0,相同时间地温均变现为30 cm>20 cm>10 cm>0 cm。不同深度的地温日变化趋势相似,均呈现单峰分布。最低值出现在1月,而最高值出现在7月,7月以后不同深度地温急剧下降,相同月份地温均变现为30 cm>20 cm>10 cm>0 cm。
2.2 热带季节雨林林冠温度与相同高度气温分布
西双版纳热带季节雨林林冠温度与相同高度的气温在同一季节有相似的日变化特征(图2)。热带季节雨林冠层温度在雾凉季、干热季和雨季均表现出一致的变化规律,呈明显的单峰变化曲线,在1:00,24:00最低,在14:00左右出现最高。在雾凉季林冠温度和同层高度大气温度均较低,日较差较大,干热季二者温度均有升高,且日较差最大,至于雨季,二者温度均较高,但日较差是3个季节中最小的。由图2还可知,西双版纳热带季节雨林气温存在显著的日变化,气温的日较差在雾凉季和干热季最高可达15℃,白天较高的温度下充足的太阳辐射有利于植物进行光合作用,夜间较低的温度下植物呼吸作用渐弱,温度存在较大的日较差有利于植物进行有机物质的积累,是西双版纳热带季节雨林拥有较强固定碳能力的一个重要原因。不同季节林冠上、林冠下大气温度都有较为明显的日变化特征,在1:00,24:00最低,在14:00左右出现最高。热带季节雨林林冠上、林冠下的大气温度均表现为雾凉季气温较低,日较差较大;干热季气温稍高,日较差最大;雨季气温高,且日较差较小的分布特征。

图1热带季节雨林地温时空分布特征

图2热带季节雨林林冠温度与相同高度气温分布
2.3 热带季节雨林平均CO2浓度的日变化
植被—大气—土壤连续体(SPAC)之间相互作用形成森林CO2浓度的分布格局。图3显示了热带季节雨林林冠上方CO2浓度的日变化。在日尺度上,林冠上方平均CO2浓度的时间变化趋势十分相近:清晨在太阳辐射的作用下,林冠上方的平均CO2浓度不断上升;CO2浓度在夜间出现最低值,表明了昼间植物的光合作用对大气中CO2的吸收作用;随后,伴随着植物光合作用的增强,平均CO2浓度不断升高,在9:00左右达到最大,之后出现滞留阶段,在9:00—15:00出现下降趋势;入夜以后,因受植物呼吸作用和夜间稳定大气状况的影响,CO2不断蓄积,林冠上方的平均CO2浓度不断增加,之后,CO2浓度迅速下降。林内近地层平均CO2浓度的日变化与林冠上方有所不同,不同月份间也有较大差异,总体上随着月份呈倒V的变化趋势,在7—9月份达到最大,1—2月,11—12月份较低,主要受温度和雨季的影响。
2.4 热带季节雨林平均CO2浓度的季节变化
从林冠上方和林内近地层的平均CO2浓度的季节变化(图4)可知,昼间林冠上方和林内近地层平均CO2浓度均以14:00最高,在夜间最低;其中林冠上方和林内近地层平均CO2浓度日变化呈一致的倒V字变化规律。在雾凉季,热带季节雨林内近地层平均CO2浓度在562~654 mg/m3变动,而在其林冠上方平均CO2浓度在552~631 mg/m3变化;在干热季,热带季节雨林内近地层平均CO2浓度在587~689 mg/m3变动,而在其林冠上方平均CO2浓度在572~671 mg/m3变化;在雨季,热带季节雨林内近地层平均CO2浓度在572~675 mg/m3间变动,而在其林冠上方平均CO2浓度在561~653 mg/m3间变化。相同时间,CO2浓度均表现为林冠上方>林内近地层;不同季节CO2浓度均干热季>雨季>雾凉季。
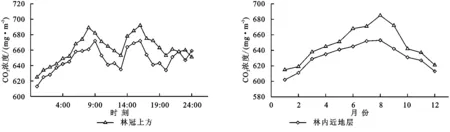
图3热带季节雨林平均CO2浓度的日变化

图4热带季节雨林平均CO2浓度的季节变化
2.5 热带季节雨林土壤呼吸日变化和季节特性
图5为热带季节雨林土壤呼吸日变化和季节特性,由图可知,热带季节雨林土壤呼吸存在明显的日变化规律,表现为单峰型,且峰值出现的时间在14:00左右达到最大,最低值出现在早上6:00,在6:00—10:00,土壤呼吸升高缓慢,12:00以后急剧上升,达到最大值以后急剧降低,土壤呼吸日变化范围为0.42~1.96 μmol/(m2·s)。与此同时,热带季节雨林土壤呼吸存在明显的季节变化规律,表现为单峰型,在7—8月份达到峰值,在12月份最低,在1—7月份,土壤呼吸升高缓慢,8月份以后,土壤呼吸急剧下降,其中土壤呼吸季节变化范围为0.53~1.95 μmol/(m2·s)。
2.6 热带季节雨林温度和土壤呼吸对CO2浓度的响应
为进一步探讨热带季节雨林温度和土壤呼吸对CO2浓度的响应,国内外学者一般采用线性模型、二次方程、指数模型等多种方法进行拟合,本试验结果中将土壤呼吸强度与林冠温度进行曲线拟合,筛选出最佳拟合度的方程(R2最大,p值最小),得出的不同拟合方程,由表1可知,土壤呼吸与林冠温度之间关系以指数方程拟合最好。不同季节热带雨林土壤呼吸、CO2浓度均与林冠温度的指数关系达到了极显著水平(p<0.01),且指数模型的决定系数最大,故指数模型的拟合效果最好,从指数模型来看,雾凉季土壤呼吸与林冠温度的指数关系具体表现为y=5×10-4x2.157(p<0.001),干热季指数关系具体表现为y=6×10-4x1.985(p<0.001),雨季指数关系具体表现为y=4×10-4x3.068(p<0.001);雾凉季林冠上方CO2浓度与林冠温度的指数关系具体表现为y=0.0589x-1.478+3.489(p<0.001),干热季指数关系为y=0.0421x-2.214+6.987(p<0.001),雨季指数关系为y=0.0475x-1.9568+4.185(p<0.001);林内近地层CO2浓度与林冠温度的指数关系具体表现为y=0.288x-1.789+1.587(p<0.001),干热季指数关系为y=0.389x-2.741+4.495(p<0.001),雨季指数关系为y=0.502x-1.578+3.078(p<0.001)。
2.7 热带季节雨林CO2浓度与环境因子相关分析
由表2可知,林冠上方CO2浓度与光合有效辐射之间皆表现出正相关,随着光合有效辐射的增强,CO2浓度相应地升高,这与上面的分析是一致的。由于西双版纳热带季节雨林存在明显的落叶现象,旱季辐射虽然很强,叶片凋落造成光合碳吸收减少,CO2浓度高;雨季辐射减弱,但是叶面积指数高,光合碳吸收总量多,CO2浓度低;林冠上方CO2浓度与叶面积指数之间显著的负相关关系也印证了这一点。林冠下方CO2浓度与光合有效辐射之间的正相关关系较林冠上方强,这是因为辐射的增加对土壤和生态系统温度的增温有促进作用,进而使呼吸作用增加,CO2浓度上升,辐射的直接影响只是一个表象。而地温与林冠上方CO2浓度和林内近地层CO2浓度均没有显著的相关性。林内近地层CO2浓度与土壤温度以及土壤呼吸的相关系数为正,土壤呼吸可以认为是影响和控制近地层CO2浓度的主要因子。


图5 热带季节雨林土壤呼吸日变化和季节特性

表2 热带季节雨林CO2浓度与环境因子相关分析
3 讨论与结论
土壤是由大小不同的固相颗粒按不同比例组合而成的,土壤对水分和热量均具有吸收作用[14]。土壤温度的日变化与太阳辐射、日照、气团活动等要素有关。一方面,西双版纳热带季节雨林内太阳辐射被各层植被反射和吸收[15];另一方面,土壤对太阳辐射也有削弱作用,土层深度越深削弱作用越明显,因而土壤的最高温一般出现在表层土壤,并且表层土壤由于受太阳辐射以及昼夜气温交替影响较大,地温呈现出明显的日变化特征;而深层土壤受到外界环境的影响较小,使得深层地温的昼夜变化较小,接近于恒值,这与其他热带地区的研究结果基本一致[16]。由于土壤对热量的吸收作用,表层地温达到极值的时间要稍稍落后于气温达到极值的时间,并且日温度最高值出现时刻随深度增加呈现后移。
通过研究发现,对于冠层上方而言,其CO2浓度体现出明显的日变化差异,这主要是由于大气层具有显著差异的活动变化,在夜晚,大气边界层的底部会受气温影响形成较为稳定的边界层,而其上方形成所谓的残留层,待太阳照射增强的时候,源源不断的太阳辐射进入大气层,此时与原有的稳定边界层形成明显的热力对流,其中的稳定边界层很快消失,进而形成较大的混合层,并对上方的残留层形成显著的侵蚀[17],至上午11:00左右,将完全打破原有的残留层,在大气混合作用之下形成通风足够的边界层。边界层能够对近地层大气产生显著影响,从而对林冠上方的大气施加影响,而在夜晚大气处于相对稳定状态,植物呼吸能够产生较多的CO2积累,该浓度能够在太阳升起之前形成峰值[18]。待太阳升起,林冠上方大气受到太阳辐射后逐渐不稳,CO2也将逐渐消散,导致其浓度迅速降低;中午时分,大气残留层基本消散,CO2浓度接近大气正常水平[19];在13:00左右,在太阳辐射等作用下林冠大气中的CO2浓度最不稳定,但随着时间的推移,其稳定性将明显增强,随着太阳辐射达到峰值,对于光合作用而言,其日变化曲线也形成单峰型[20],而此时植物的光合作用依然比较明显,并没有产生“午休”,光合作用此时会吸收不少的CO2,这就会导致其浓度在大气之下,离林冠越近,其浓度越低,在下午15:00左右达到最小值[21];下午随着太阳辐射的减弱,植物光合作用也受到此影响,而此时大气日渐稳定,尤其是夜晚大气较为稳定,植物呼吸产生的CO2在林冠上方不断积累,故此在太阳升起之前达到峰值[22]。大气影响作用之下将会影响CO2浓度的空间分布,在研究植物群落光合作用的过程中,这方面的研究相对较少,而CO2在光合作用中发生着无可取代的作用,作为光合作用的能量之一,必然影响整个生态循环,因此在实际研究过程中必然要考虑其时空分布。
对于近4个月的雾凉季来说,浓雾天气较多,太阳辐射量大大降低,这直接导致植物光合作用所需能量来源降低,土壤和植物释放的CO2不足以满足光合作用所需,因此在林冠上方形成较高浓度的CO2[23],在雾凉季过后,随着干热季的到来,空气日渐干燥,土壤水分逐渐降低,此时逐渐出现林木大面积换叶的现象,形成西双版纳独特的雨林现象;尽管在4月出现全年较高水平的太阳辐射,但是因叶片脱落等原因导致光合作用显著下降,导致在光合作用过程中难以消化大量的CO2,另外,随着土壤温度的上升,土壤呼吸作用增强,其能够释放较多的CO2,此阶段的林冠覆盖面积较少,在风速的作用之下CO2更容易被吹拂到林冠上方,使得此阶段的CO2浓度处于较高水平,尤其是3月份这一现象更加明显,很容易出现浓度的峰值[24]。逐渐进入雨季,林木生长日益旺盛,雨林发育加快,光合作用能力自然显著增强,但降雨导致太阳辐射降低,对光合作用形成显著制约,故CO2浓度并没有出现显著降低[25];进入6月,植物生长达到鼎盛期,太阳辐射量也能够满足植物生长,光合作用效果显著,这使得此时期的CO2浓度进入低谷。进入9月后气温降低,这使得林冠的呼吸作用减弱,导致CO2浓度处于降低。从雾凉季开始,太阳辐射大幅下降,光合作用也随之降低,碳吸收能力下降明显,故而CO2浓度将会提升[26]。
就热带雨林而言,在温差的影响之下,其林冠温度也呈现典型的昼夜差异,昼夜温差能够达到10℃,白天温度较高,能够促进植物的光合作用,而在下午温度降低的情况下植物的呼吸能力下降,植物能够进行有机质积累,随着大气温度在下午两点左右能够达到较高水平,土壤呼吸对于温度的反应较为敏感,因此在此阶段土壤呼吸形成相对峰值[27]。通过研究发现,土壤呼吸具有明显的日变化特点,林冠在白天能够充分吸收利用外界能量,而夜间则保持相对稳定状态,从而对林内外气温起到稳定的调节作用,利于气温保持。不少学者的研究发现,对于森林及草原等土壤而言,其呼吸表现出明显的单峰特征[28],通过本研究发现雨林土壤呼吸也具有较为显著的单峰曲线变化特点,当天的高峰一般出现在下午两点左右,而低值出现在早上六点左右,此时地表温度明显低于其他时段,且具有较高的湿度[22-23],根系的呼吸等活动能力相对较弱。
本研究中,林冠上方CO2浓度与光合有效辐射之间皆表现出正相关,随着光合有效辐射的增强,CO2浓度相应地升高,这与上面的分析是一致的。由于西双版纳热带季节雨林存在明显的落叶现象,旱季辐射虽然很强,叶片凋落造成光合碳吸收减少,CO2浓度高;雨季辐射减弱,但是叶面积指数高,光合碳吸收总量多,CO2浓度低;林冠上方CO2浓度与叶面积指数之间显著的负相关关系也印证了这一点[29]。林冠下方CO2浓度与光合有效辐射之间的正相关关系较林冠上方强,这是因为辐射的增加对土壤和生态系统温度的增温有促进作用,进而使呼吸作用增加,CO2浓度上升,辐射的直接影响只是一个表象[30]。而地温与林冠上方CO2浓度和林内近地层CO2浓度均没有显著的相关性。林内近地层CO2浓度与土壤温度以及土壤呼吸的相关系数为正,土壤呼吸可以认为是影响和控制近地层CO2浓度的主要因子。
参考文献:
[1] 周玉荣,于振良,赵士洞.我国主要森林生态系统碳贮量和碳平衡[J].植物生态学报,2000,24(5):518-522.
[2] Pausch J, Kuzyakov Y. Carbon input by roots into the soil: Quantification of rhizodeposition from root to ecosystem scale.[J]. Global Change Biology,2018,24(1):1-12.
[3] Walker A P, Zaehle S, Medlyn B E, et al. Predicting long-term carbon sequestration in response to CO2enrichment: How and why do current ecosystem models differ[J]. Global Biogeochemical Cycles, 2015,29(4):476-495.
[4] 肖强,肖洋,欧阳志云,等.重庆市森林生态系统服务功能价值评估[J].生态学报,2014,34(1):216-223.
[5] Yue K, Peng Y, Peng C, et al. Stimulation of terrestrial ecosystem carbon storage by nitrogen addition: a meta-analysis[J]. Scientific Reports, 2016,6:19895.
[6] 吴雅琼,刘国华,傅伯杰,等.森林生态系统土壤CO2释放随海拔梯度的变化及其影响因子[J].生态学报,2015,35(11):306-313.
[7] Walker T N, Garnett M H, Ward S E, et al. Vascular plants promote ancient peatland carbon loss with climate warming[J]. Global Change Biology, 2016,22(5):1880-1889.
[8] 李贵才,韩兴国,黄建辉,等.森林生态系统土壤氮矿化影响因素研究进展[J].生态学报,2015(7):148-156.
[9] Pries C E H, Schuur E A G, Natali S M, et al. Old soil carbon losses increase with ecosystem respiration in experimentally thawed tundra[J]. Nature Climate Change, 2016,6(2):214-218.
[10] Qin Y, Chen J, Yi S. Plateau pikas burrowing activity accelerates ecosystem carbon emission from alpine grassland on the Qinghai-Tibetan Plateau[J]. Ecological Engineering, 2015,84:287-291.
[11] 陈全胜,李凌浩,韩兴国,等.典型温带草原群落土壤呼吸温度敏感性与土壤水分的关系[J].生态学报,2015(4):175-180.
[12] 杨玉盛,陈光水,王小国,等.中国亚热带森林转换对土壤呼吸动态及通量的影响[J].生态学报,2015(7):156-162.
[13] 张红星,王效科,冯宗炜,等.黄土高原小麦田土壤呼吸对强降雨的响应[J].生态学报,2015(12):408-415.
[14] 王小国,朱波,王艳强,等.不同土地利用方式下土壤呼吸及其温度敏感性[J].生态学报,2015(5):290-298.
[15] Nagelkerken I, Connell S D. Global alteration of ocean ecosystem functioning due to increasing human CO2emissions[J]. Proceedings of The National Academy of Sciences, 2015,112(43):13272-13277.
[16] 张方月,王清奎,于小军. CO2浓度升高和N沉降对中亚热带森林土壤呼吸的短期影响[J].生态学杂志,2015,34(6):1638-1643.
[17] 阿力木,阿巴斯,何清,等.肖塘地区夏季土壤CO2浓度日变化特征及影响因素[J].沙漠与绿洲气象,2016(2):63-69.
[18] Motte L G D L, Mamadou O, Beckers Y, et al. Rotational and continuous grazing does not affect the total net ecosystem exchange of a pasture grazed by cattle but modifies CO2, exchange dynamics[J]. Agriculture Ecosystems & Environment, 2018,253:157-165.
[19] 龙凤玲,李义勇,方熊,等.大气CO2浓度上升和氮添加对南亚热带模拟森林生态系统土壤碳稳定性的影响[J].植物生态学报,2014,38(10):1053-1063.
[20] 项文化,黄志宏,闫文德,等.森林生态系统碳氮循环功能耦合研究综述[J].生态学报,2015(7):307-314.
[21] 刘魏魏,王效科,逯非,等.全球森林生态系统碳储量,固碳能力估算及其区域特征[J].应用生态学报,2015,26(9):2881-2890.
[22] 周国逸,张德强,李跃林,等.长期监测与创新研究阐明森林生态系统功能形成过程与机理[J].中国科学院院刊,2017,32(9):1036-1046.
[23] 胡芳,杜虎,曾馥平,等.广西不同林龄喀斯特森林生态系统碳储量及其分配格局[J].应用生态学报,2017,28(3):721-729.
[24] Novick K A, Oishi A C, Ward E J, et al. On the difference in the net ecosystem exchange of CO2between deciduous and evergreen forests in the southeastern United States[J]. Global Change Biology, 2015,21(2):827-842.
[25] 牛晓栋,江洪,张金梦,等.浙江天目山老龄森林生态系统CO2通量特征[J].应用生态学报,2016,27(1):1-8.
[26] 李银,陈国科,林敦梅,等.浙江省森林生态系统碳储量及其分布特征[J].植物生态学报,2016,40(4):354-363.
[27] Knauer J, Zaehle S, Reichstein M, et al. The response of ecosystem water-use efficiency to rising atmospheric CO2concentrations: sensitivity and large-scale biogeochemical implications[J]. New Phytologist, 2017,213(4):1654-1666.
[28] 许文强,杨辽,陈曦,等.天山森林生态系统碳储量格局及其影响因素[J].植物生态学报,2016,40(4):364-373.
[29] 魏书精,罗碧珍,孙龙,等.森林生态系统土壤呼吸时空异质性及影响因子研究进展[J].生态环境学报,2013,22(4):689-704.
[30] García-Palacios P, Vandegehuchte M L, Shaw E A, et al. Are there links between responses of soil microbes and ecosystem functioning to elevated CO2, N deposition and warming: A global perspective[J]. Global Change Biology, 2015,21(4):1590-1600.
