黄土丘陵区植物群落功能多样性/物种多样性与产流产沙响应关系
2018-06-15温仲明朱朵菊曾鸿文
陶 宇, 温仲明, 朱朵菊, 曾鸿文, 张 静
(1.西北农林科技大学, 陕西 杨凌 712100; 2.中国科学院 水利部 水土保持保持研究所, 陕西 杨凌 712100)
植被是控制水土流失的关键因素,长期以来,恢复植被都被看作是控制土壤侵蚀、改善生态环境的重要途径,经过长期的植被恢复措施,黄土高原的水土流失状况已有了很大改善,但局部地区水土流失严重的现象仍然存在。可能的原因是这些局部地区的人工植被恢复往往存在物种单一、结构较差的问题[1-2]。这些植被可能具有较高的植被盖度,但因较差的植被结构,难以控制水土流失。黄健熙等[3]的研究表明,若仅以覆盖度作为植被因子指标进行C因子的计算,会导致土壤侵蚀评价结果出现较大误差。高光耀等[4]分析了植被斑块、坡面覆被格局对水土流失的影响,认为植被类型、层次结构和形态特征是植被斑块尺度上影响径流泥沙的关键因素;雷婉宁等[5]在前人研究的基础上,耦合各植被结构层,提出了结构化植被指数的概念与计算方法,但该指数在大尺度提取上存在困难,对一些常绿植被效果较差,探寻合适的植被指数仍是植被与土壤侵蚀关系中的难点之一。
随着功能生态学研究的深入,基于性状的方法越来越多地被用来解释群落组成和生态系统过程之间的关系[6-7],一些新概念或方法为研究群落特性与侵蚀产沙提供了新的借鉴。在理论上,植物形态特征、根直径和根的拉伸强度,对土壤侵蚀的程度有着显著的影响[8-11]。Zhu等[12]的研究发现,群落功能分异度对土壤侵蚀有很强的负效应,而且这种效应在不同的降雨事件之间表现出很好的稳定性;一些研究则采用物种多样性评估植被减少侵蚀的效益,因为物种多样性会显著影响植物群落覆盖度、枯落物凋落量、个体密度和高度等群落结构的形成[13],从而对侵蚀产生影响;王震洪等[14]的研究发现物种多样性高的生态系统相较于物种多样性低的生态系统,具有更好的水土保持功能。
但物种功能多样性与物种多样性在土壤侵蚀研究中,哪个指标能更好地反映植被的水土保持效益则少有研究涉及。鉴于此,本研究试图采用植物功能多样性与物种多样性的指标,以黄土丘陵区的坡面草地群落为对象,构建具有不同物种与功能性状结构的径流小区,通过人工降雨试验,研究群落功能性状多样性、物种多样性与侵蚀产沙的响应关系,评估物种多样性、功能多样性与群落减水减沙能力关系的敏感性,为选择合适的植被指数提供依据。
1 材料和方法
1.1 研究区概况
研究区域设在安塞县纸坊沟流域的沿河湾镇瓦树塌村(北纬36°44′43″;东经109°14′45″),属黄土丘陵沟壑区。年均气温8.8℃;年平均降水量为549.1 mm,降水年际变率大,枯水年只有300 mm左右,丰水年可700 mm以上。研究对象为坡面自然恢复的草本群落,主要物种有达乌里胡枝子(Lespedezadavurica)、长芒草(StipabungeanaTrin)、铁杆蒿(Artemisiavestita)、蒙古蒿(Artemisiamongolica)等。土壤为黄土母质上发育而成的黄绵土,流域内除部分塌地外,具有坡度陡,土壤侵蚀强烈,岩石裸露等特点,土壤侵蚀模数为14 000 t/hm2。
1.2 试验方法
1.2.1 试验设计 在试验区内选择9个草本群落建立径流小区(同一坡面,地形条件、气候与土壤条件一致),小区规格设为2 m×5 m;为便于根据试验需求对小区物种进行调控,本研究选择的植物群落既要是自然恢复的群落,也要求物种不能过多。在选定的小区中,群落构成主要有达乌里胡枝子、铁杆蒿、长芒草、蒙古蒿4个物种,物种较少,便于调控。在选择的小区内,采用物种剔除法,设置了物种组成不同但植被盖度大致相同的9个小区(表1),在各群落进行物种调查。小区处理完毕后,先进行降雨试验,降雨试验结束后,对各小区内的物种进行性状测定,然后计算各小区物种多样性与功能多样性的相关指数,并分析两类多样性指标与侵蚀产沙的响应关系。
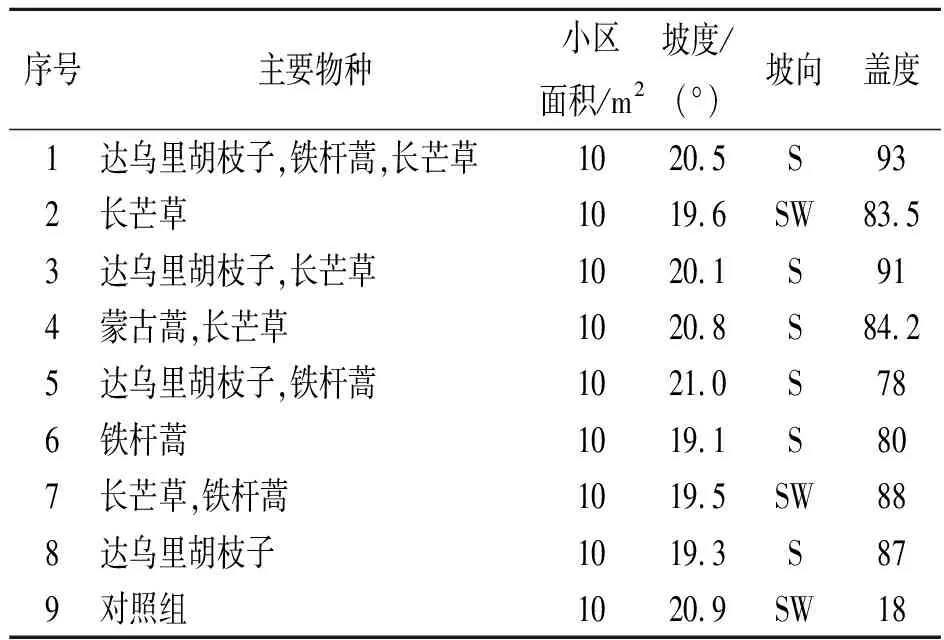
表1 径流小区植被状况
1.2.2 人工降雨试验 降雨试验在2016年8月至9月进行。设备采用顶喷式降雨器,雨强变化范围20~300 mm/h,降雨均匀度大于0.7。首先在处理后的径流小区四周,用厚5 mm、宽35 cm的铁板垂直嵌入地表15 cm,以避免径流侧流集水槽置于小区底部,并在集水槽下部放置径流桶用以接取径流泥沙样品。小区搭建好之后,用PVC布对小区进行遮盖,进行雨强的率定,待雨强稳定在120 mm/h左右时,撤去PVC布,开始降雨,待雨滴到达地面后开始计时,记录下初始产流时间,在产流后的前3 min每1 min接1次径流泥沙全样,之后每3 min接1次径流且只取中间的1 min所产生的径流。在产流30 min后,径流基本趋于稳定,停止降雨,并记录下结束产流时间。在整个降雨过程的前期,中期和后期分别测定3次雨强。降雨结束后对降雨中接取的径流泥沙混合样进行称重,静置一段时间后,滤去上清液,带回实验室,烘干24 h称重。降雨结束后,对小区进行调查。包括植物名称、盖度、株树(丛数)、高度、株幅、多度,并记录每个物种取样比例。
1.2.3 植物功能性状测定 本研究测定的植物功能性状主要有每个物种的比叶面积、叶组织密度、叶体积、比根长、根组织密度及全氮含量6个性状指标。在每个植株采集直径小于2 mm的细根,并沿东西南北4个方向采集10~15片光照条件良好、完全伸展且没有病虫害的叶片[15]。对采集的全部物种用电子游标卡尺(精度0.01)测量每片叶子的厚度,测量时避开叶脉。在叶片前、中、末端分别测量叶片厚度,然后取其平均值,即为叶片厚度。用扫描仪对平展的叶片进行扫描,然后用图像分析软件IPP(image pro-plus)根据扫描图片的像元数计算叶片面积。将采集的细根(直径<2 mm)用清水洗掉附着的泥土并去除根毛,用精度为0.01 mm的电子游标卡尺测量10条细根的长度,将新鲜洗净的细根完全侵入盛水的量筒约5 s,读取量筒中水增加的体积即为根体积(排水替代法)。然后将叶片和细根样品放入80℃的烘箱内烘干48~72 h,用精度为万分之一的天平称取叶片和细根的干重。最后用所测得的数据计算各植物功能性状,比叶面积(SLA)=叶面积/叶干重(mm2/mg);叶组织密度(LTD)=叶干重/叶体积(mg/mm3),其中,叶体积(LV)=叶面积×叶厚度;比根长(SRL)=根长/根干重(m/g),根组织密度(RTD)=根干重/根体积(mg/mm3)。将烘干的叶片和细根样品粉碎,用H2SO4—H2O2法消解,采用凯氏定氮法分别测定植物叶片全氮含量LN(g/kg)和细根的全氮含量RN(g/kg)[15]。
1.2.4 群落多样性指数计算 研究共选择6个指数来表征群落功能多样性和物种多样性(表2),用以了解坡面产流产沙与不同多样性指数的响应关系,评估物种多样性、功能多样性与群落减水减沙能力关系的敏感性,为选择合适的植被指数提供依据。各指标计算方法如下[16]:
(1) 功能丰富度(Functional richness):
(1)
式中:FRic为群落i中特征c的功能丰富度;SFic为群落内物种所占据的生态位;Rc为绝对特征值范围。
(2) 功能均匀度指数(Functional evenness)[16]:
(2)
式中:S为物种数;Pi为i的相对丰富度。
(3) 功能离散度指数(Functional divergence)[16]:
(3)
式中:FDiv为功能离散度指数;Ci为第i项功能特征的数值;Ai为第i项功能特征的丰度比例;lnx为物种特征值的自然对数;N为群落中的物种数。
(4) Simpson指数的表达式为:
(4)
式中:N为全部种的个体总数;Ni是i中的个体数.
(5) Shannon-Wiener指数其表达式如下:
(5)
式中:S为物种数;Pi为第i种的个体的比例.
(6) Pielou指数表达式为:
(6)
式中:H是实际观察的物种多样性指数;S是物种总数。

表2 功能多样性指数和物种多样性指数简介
1.2.5 数据处理 试验数据处理利用SPSS 22.0,R和Excel完成。功能多样指数和物种多样性指数使用R中FD包和vegan包计算完成。初始产流和结束产流时间与功能多样性指数之间的相关性和不同指数与产流总量产沙总量之间的相关性检验均采用SPSS 22.0中双变量相关性方法进行分析。不同小区之间产流总量和产沙总量的差异性采用单样本T检验进行分析。不同指数与产流产沙总量之间的关系采用一元线性回归方法分析。
2 结果与分析
2.1 径流小区群落功能多样性指数
各径流小区群落多样性指数差异是分析小区植被与径流产沙响应差异的基础。在对各小区植物性状测定的基础上,利用R中的FD包计算各小区群落的功能多样性指数包括功能丰富度指数、功能均匀度指数、功能离散度指数等(表3)。
经单样本检验,功能多样性指数与物种多样性指数在各小区间差异显著(p<0.05),达乌里胡枝子、铁杆蒿、长芒草群落具有较高的功能丰富度指数,而只有铁杆蒿群落的功能丰富度指数最低,具有较多物种的群落其功能丰富度指数普遍较高。功能均匀度指数表现出了与功能丰富度指数相反的情况,铁杆蒿群落功能均匀度指数最高,达乌里胡子子群落的功能均匀度指数最低。功能离散度指数在有长芒草的群落中都较高,并且达乌里胡枝子群落也具有较高的功能离散度指数。物种多样性Shannon-Wiener指数在组合物种群落中较高,单物种群落其Shannon-Wiener指数较低,并且Simpson指数和Shannon-Wiener指数是显著正相关的,所以组合物种群落中Simpson指数也比较高。Pielou均匀度指数在不同的群落之间差异比较大。
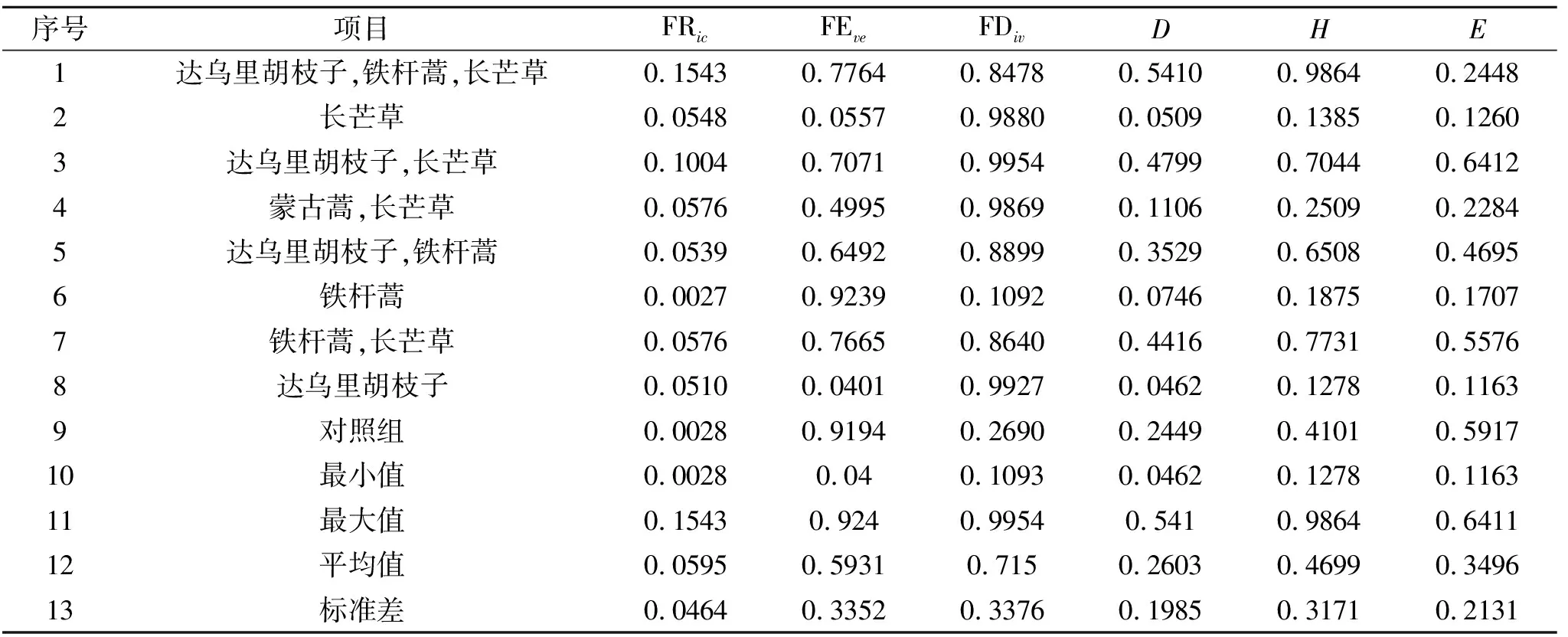
表3 多样性指数计算结果
2.2 不同群落下产流产沙过程与特征
(1) 产流时间变化特征。以降雨开始时间为起点,到薄层径流汇集到小区出口为终点,这期间的所用时长我们称之为产流起始时间。产流结束时间是指降雨结束到小区出口无径流流出所需的时间(表4)。这2个指标可以反映小区内植被状况的延缓径流和持蓄降水的能力。从结果中可以看出大部分组合物种群落中的初始产流时间在2.5 s左右。但是达乌里胡枝子、长芒草群落,蒙古蒿、长芒草群落的初始产流时间显著的低于2.5 s。结束产流时间各群落相差不大。并且通过相关性检验可以看出,FRic即群落功能丰富度指数与结束产流时间在0.05水平上有显著的负相关关系(表5)。
(2) 不同群落小区累计径流量/产沙量与径流时间的关系。不同植物群落不仅具有不同的产流时间变化特征,产流产沙过程也各不相同。对不同群落径流小区坡面累计径流量、累计产沙量和降雨历时间进行回归分析(图1—9),表明累计径流量与降雨历时之间有较好的线性关系(R2>0.9),径流量的增量为一个较为平稳的状态。累计产沙量和降雨历时之间的关系为二次函数关系,说明在降雨后期产沙量在逐渐降低,产沙速率减慢。同时通过累计产沙量与降雨历时的关系,可以看出多数情况下,在有铁杆蒿或蒙古蒿存在的小区中,其累计产沙量从开始都一直处在一个较为平稳的增长过程,而其他不存在这两个物种的小区,累计产沙量在开始阶段都有一个较快的增长过程。出现这种情况可能的原因是蒿类植物由于其植株硬度比较大,吸收雨滴冲击力的能力比较强,所以在雨滴接触到其植株后,降雨动能经历了比较大的衰减,从而使得雨水冲刷地表的能力遭到了削弱。而没有这两种植物的小区,由于达乌里胡枝子、长芒草等植物地上部分是比较柔软的,无法有效削减降雨动能,所以其产沙量在开始的阶段存在急速增长的现象。
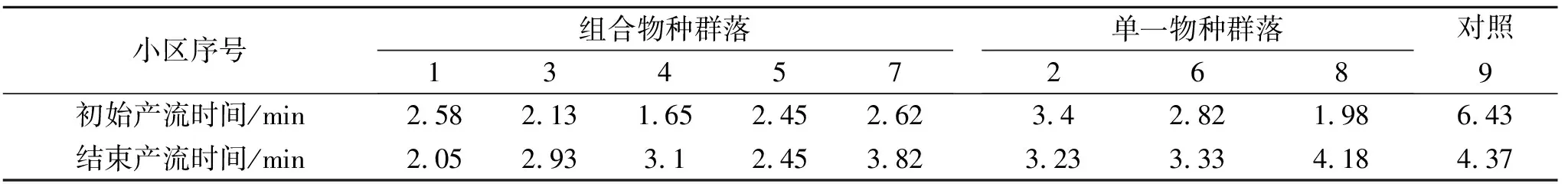
表4 不同群落初始产流和结束产流时间
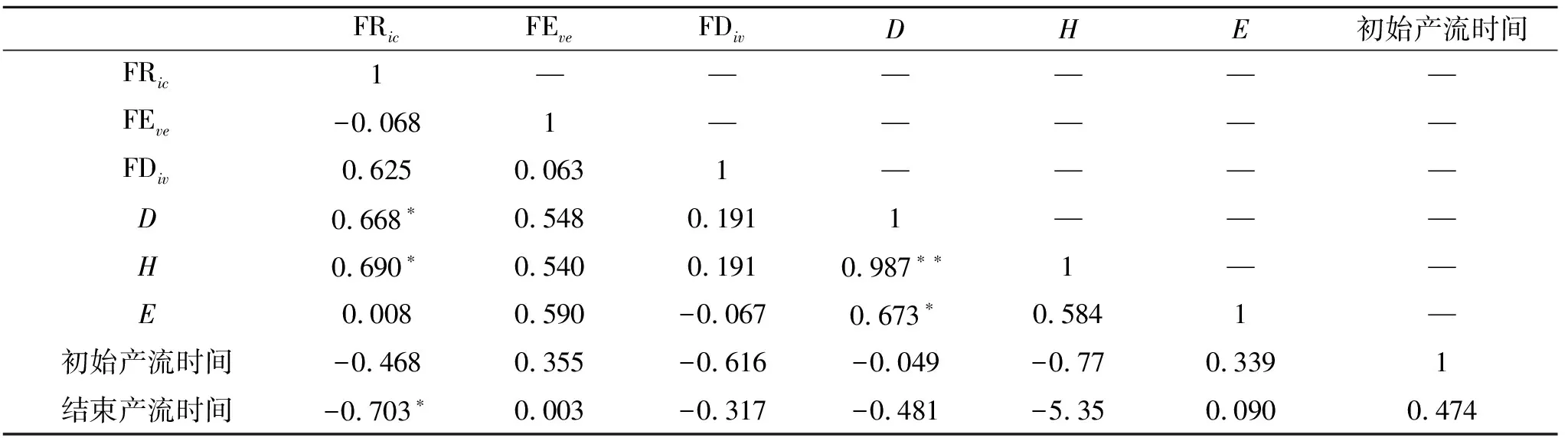
表5 初始产流和结束产流时间与功能多样性指数相关性检验结果
注:*表示p<0.05,**表示p<0.01,下表同。
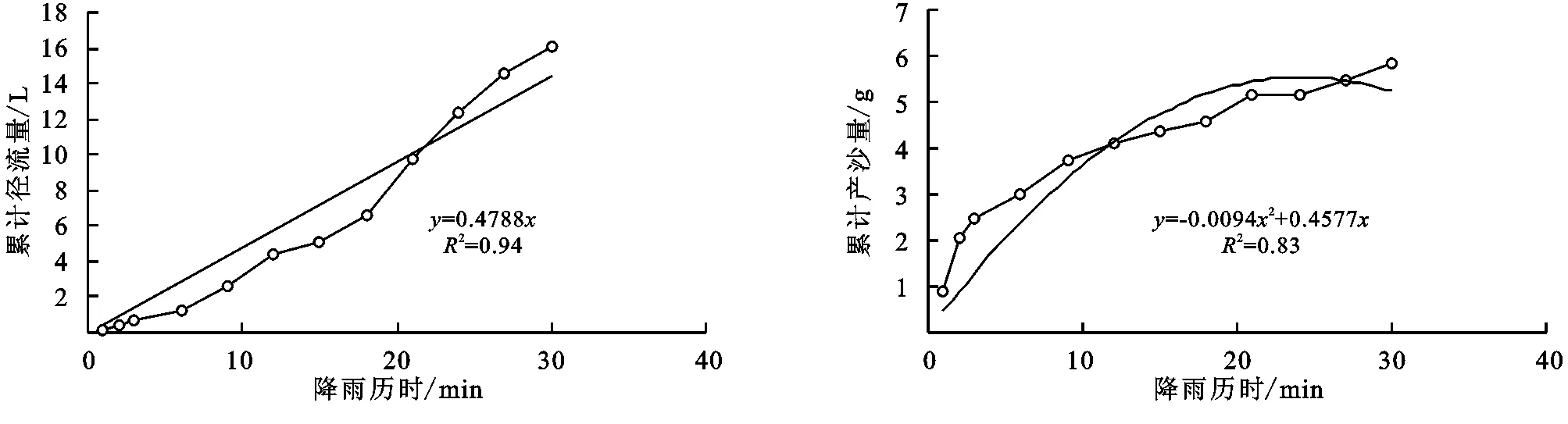
图1 达乌里胡枝子+铁杆蒿+长芒草径流小区累计径流量与累计产沙量随时间变化关系
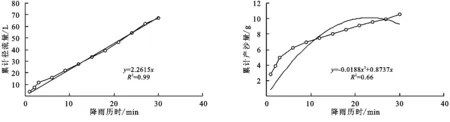
图2 长芒草径流小区累计径流量与累计产沙量随时间变化关系
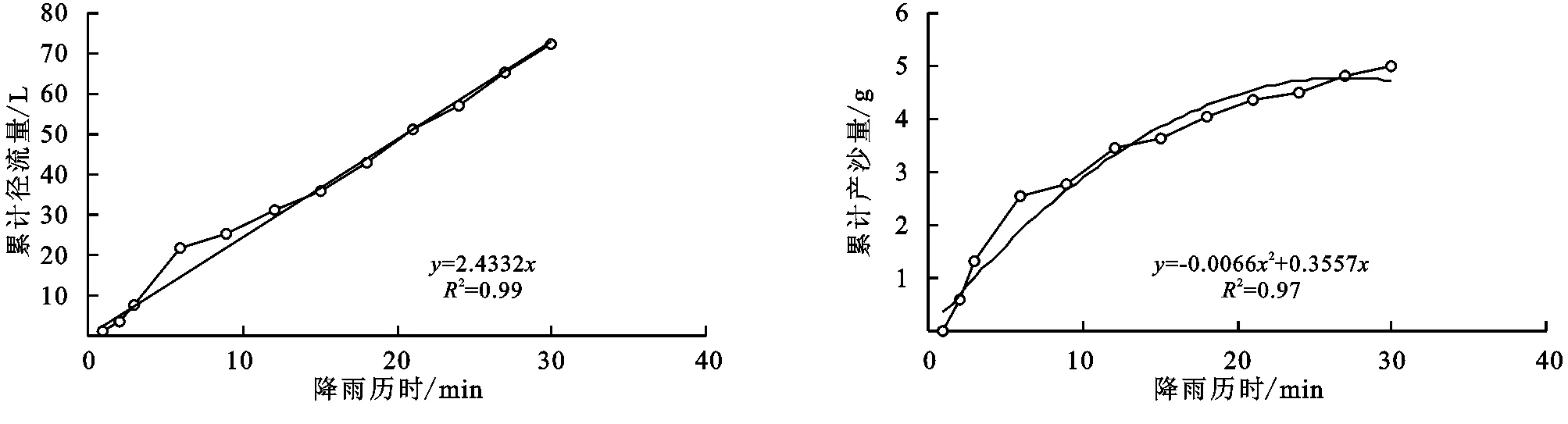
图3 达乌里胡枝子+长芒草径流小区累计径流量与累计产沙量随时间变化关系
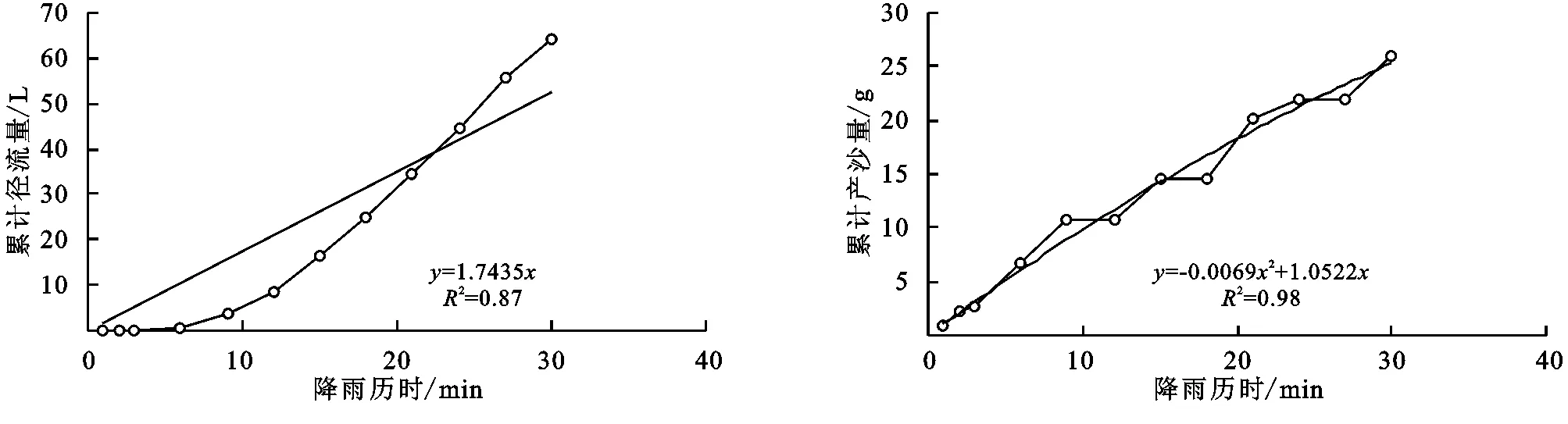
图4 蒙古蒿+长芒草径流小区累计径流量与累计产沙量随时间变化关系
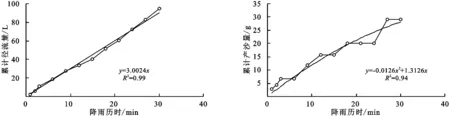
图5 达乌里胡枝子+铁杆蒿径流小区累计径流量与累计产沙量随时间变化关系
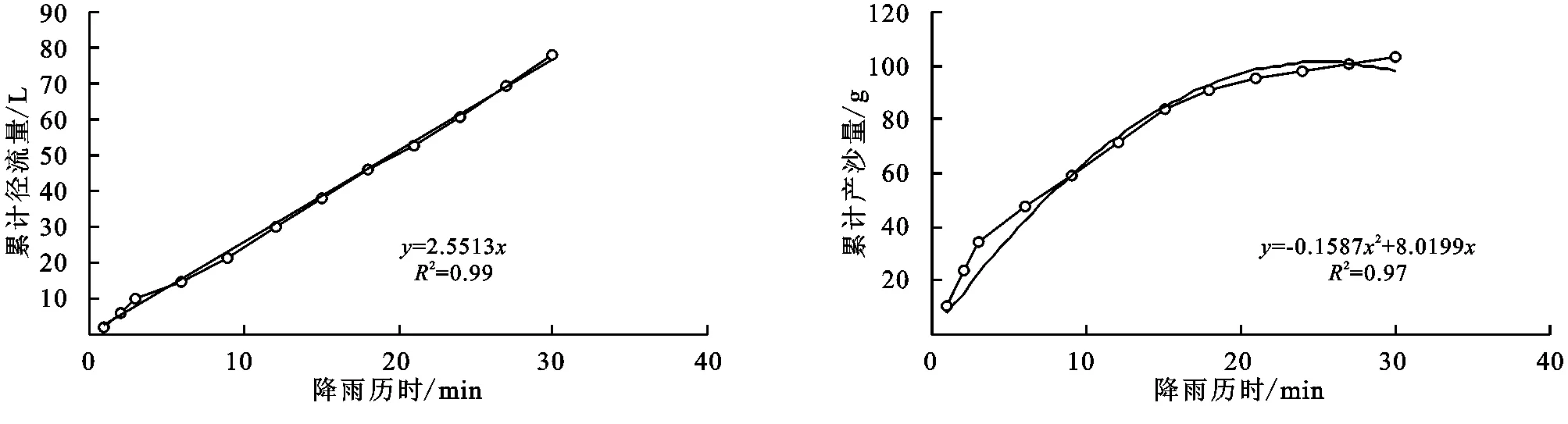
图6 铁杆蒿径流小区累计径流量与累计产沙量随时间变化关系
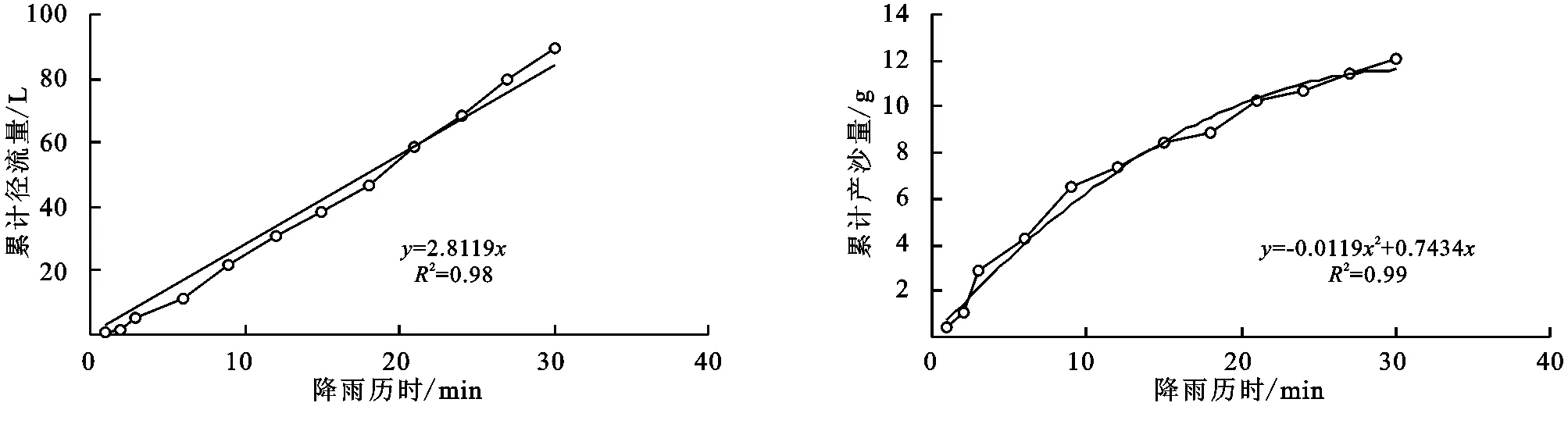
图7 铁杆蒿+长芒草径流小区累计径流量与累计产沙量随时间变化关系
2.3 群落生物多样性指数与坡面产流产沙关系
通过对不同径流小区的产流总量和产沙总量进行单样本T检验(表6),可以看出径流总量在各小区之间具有显著性差异,但是产沙总量差异不明显。
将产沙总量和产流总量与生物多样性指数(物种多样性、功能多样指数)进行相关性检验(表7)。结果表明,产流总量与功能多样性指数中只有群落功能丰富度(FRic)在0.05水平上显著负相关,产沙总量与各指数并无显著相关性。同时,其他功能多样性指数与产流、产沙总量无显著相关性,物种多样性指数与径流和产沙总量之间不存在显著的相关性,但Shannon-Wiener指数与FRic功能丰富度指数在0.05水平上显著正相关。
进一步进行回归分析发现(图10),群落FRic指数与径流总量存在线性相关关系,和其他指数之间并不存在明显的线性相关关系,和物种多样性指数也不存在显著地相关关系。

图8 达乌里胡枝子径流小区累计径流量与累计产沙量随时间变化关系
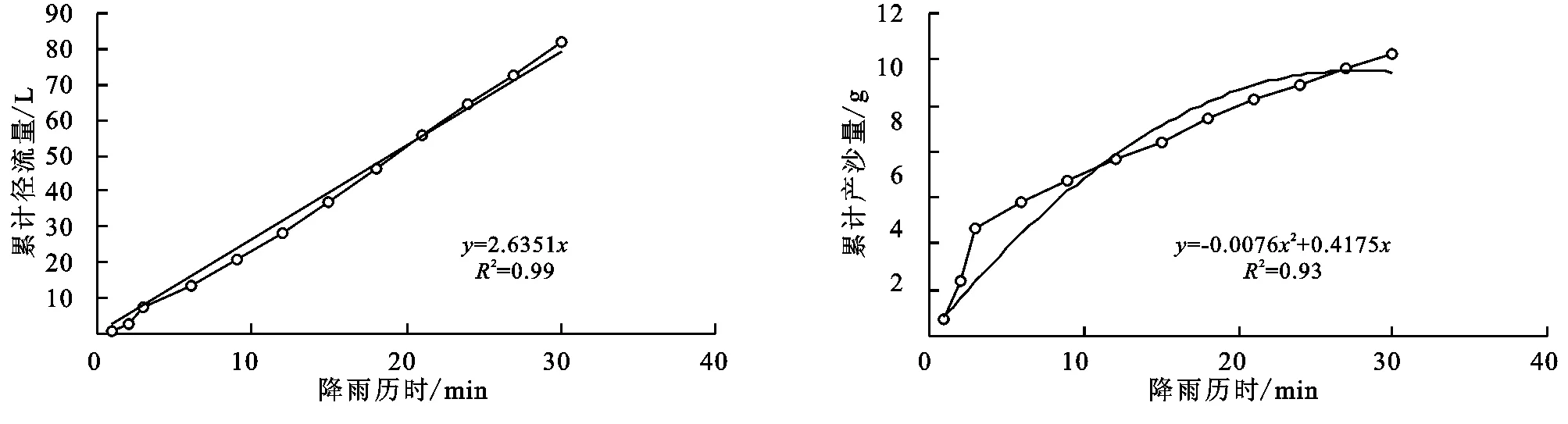
图9 杂草径流小区累计径流量与累计产沙量随时间变化关系

项目t显著性平均值差值径流总量9.330.00071155.38产沙总量2.070.07219.96
3 讨论与结论
3.1 讨 论
(1) 不同群落初始产流时间与结束产流时间差异。不同群落的初始产流时间之间是存在较大差异的。通过对比群落间物种组成的差异,可以看出存在长芒草、达乌里胡枝子等物种的小区初始产流时间普遍较短。那么可能的原因就是长芒草、达乌里胡枝子这两个物种能够为雨水的汇集提供有效路径,从而降低初始产流时间。结束产流时间方面,在长有铁杆蒿的群落中,其结束产流时间普遍在3 s以上,可能的原因是铁杆蒿植株高度较高,成簇生长,其间枝杈较多,其拦蓄降水能力比较强,在降雨结束后,叶片上的雨水沿着茎干留下来,导致其结束产流时间延长。同时FRic即群落功能丰富度指数与结束产流时间有显著负相关关系,即群落中物种占有的生态位空间越大,结束产流的时间就越短,原因在于群落占有的生态位空间越大,其植被生产力就越高,对降雨的截流作用越大;同时较高的植被生产力,可增加土壤入渗率和表面糙度,还可以有效拦蓄降水,从而缩短了结束产流时间。
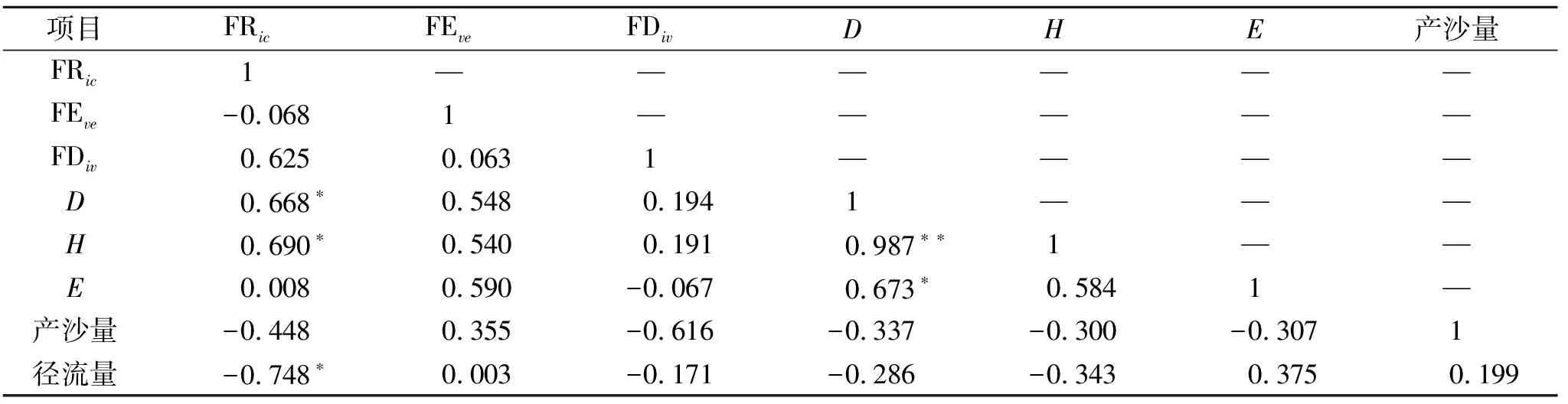
表7 产沙量和产流量与功能多样指数相关性检验结果
(2) 多样性指数与产沙量和产流量之间响应关系。在试验设计中,共选择了6个指数分别对群落的物种的功能多样性和物种多样性进行了描述,并且在处理小区时尽可能保证各小区盖度大致相同,以排除植被盖度的影响。从结果看,3个功能多样性指数中,FEve和FDiv对产流产沙没有显著影响,而FRic与径流总量呈显著负相关。物种多样性指数,虽然Shannon-Wiener指数与FRic具有显著的相关性,但是Shannon-Wiener指数在降雨事件中并没有出现于FRic一样的负效应。Simpson指数和Pielou指数对径流总量没有显著影响。
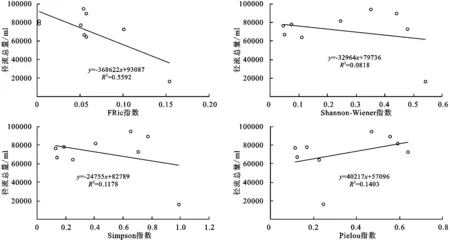
图10 径流小区FRic指数和物种多样性指数与径流总量的回归分析
Shannon-Wiener指数与FRic指数有显著正相关性,物种多样性下降或物种丧失将引起有些功能性状丢失或性状空间维度减小,导致功能多样性的降低[17]。但Shannon-Wiener指数并没有像FRic指数一样,表现出与产流产沙过程的显著相关性,表明物种丰富度只能在一定程度上反映群落的功能丰富度,但在植被盖度相近且物种多样性差异较小时,却无法提供进一步揭示植物群落结构与产流产沙过程的响应关系。出现这个现象可能与植物群落构建过程有关。植物群落的构建主要受制于环境筛选与生物过滤两种生态过程,环境筛选引起物种性状趋同[18]。竞争导致性状分离而产生性状趋异[19]。在植物群落演化过程中,物种数目可能并没有增多,但其性状距离其演化初期已经有了较大的差距,不论是趋同还是趋异,性状值都会随环境变化及生物间作用发生变化,从而影响到系统功能。而物种多样性多基于分类学信息计算,不能反映植物功能性状的种内变异。而性状的种内变异,对植物的环境适应能力及生态系统过程具有更大的影响[20]。因此,功能多样性指数具有比物种多样性更高的敏感性,这或许是造成功能多样性指数与产流产沙过程相关而物种多样性指数没有相关性的原因。这也表明,在植被恢复实践中,我们不仅要关注物种多样性组成,更要关注群落系统的功能性状结构变化,以制定更符合预期的生态调控措施与方案。
3.2 结 论
(1) 结束产流时间随着群落功能丰富度指数(FRic)的增大而减小,FRic指数和其他的指数均未与初始产流时间表现出相关性;
(2) 3个群落功能多样性指数中,只有群落功能丰富度指数(FRic)对坡面产流总量呈线性响应关系,回归结果的R2>0.5,其他指数结果的R2均在0.01左右,表明基本不存在响应关系;
(3) 通过与Simpson指数、Shannon-Wiener指数、Pielou指数比较,功能多样性指数比物种多样性指数响应敏感,能更好地反映群落水土保持效益。但植物群落性状结构是非常复杂的,单一的功能多样性指标是无法完整诠释性状结构与侵蚀产沙之间关系的,因此探索指标之间的组合模型应是未来植物群落与产流产沙响应关系研究的一个重要方向。
参考文献:
[1] Cao S, Chen L, Yu X. Impact of China′s Grain for Green Project on the landscape of vulnerable arid and semi-arid agricultural regions: a case study in northern Shaanxi Province[J]. Journal of Applied Ecology, 2009,46(3):536-543.
[2] Cao S. Impact of China′s large-scale ecological restoration program on the environment and society in arid and semiarid areas of China: achievements, problems, synthesis, and applications[J]. Critical Reviews in Environmental Science and Technology, 2011,41(4):317-335.
[3] 黄健熙,吴炳方,曾源,等.水平和垂直尺度乔、灌、草覆盖度遥感提取研究进展[J].地球科学进展,2005,20(8):871-881.
[4] 高光耀,傅伯杰,吕一河,等.干旱半干旱区坡面覆被格局的水土流失效应研究进展[J].生态学报,2013,33(1):12-22.
[5] 雷婉宁,温仲明.基于TM遥感影像的陕北黄土区结构化植被因子指数提取[J].应用生态学报,2009,20(11):2736-2742.
[6] Garnier E, Cortez J, Billès G, et al. Plant functional markers capture ecosystem properties during secondary succession[J]. Ecology, 2004,85(9):2630-2637.
[7] Mokany K, Ash J, Roxburgh S. Functional identity is more important than diversity in influencing ecosystem processes in a temperate native grassland[J]. Journal of Ecology, 2008,96(5):884-893.
[8] Baets S D, Poesen J, Gyssels G, et al. Effects of grass roots on the erodibility of topsoils during concentrated flow[J]. Geomorphology, 2006,76(1/2):54-67.
[9] De Baets S, Poesen J, Gyssels G, et al. Effects of grass roots on the erodibility of topsoils during concentrated flow[J]. Geomorphology, 2006,76(1):54-67.
[10] Pohl M, Alig D, Körner C, et al. Higher plant diversity enhances soil stability in disturbed alpine ecosystems[J]. Plant and Soil, 2009,324(1):91-102.
[11] Burylo M, Rey F, Bochet E, et al. Plant functional traits and species ability for sediment retention during concentrated flow erosion[J]. Plant and Soil, 2012,353(1):135-144.
[12] Zhu H, Fu B, Wang S, et al. Reducing soil erosion by improving community functional diversity in semi-arid grasslands[J]. Journal of Applied Ecology, 2015,52(4):1063-1072.
[13] Lavorel S, Garnier E. Predicting changes in community composition and ecosystem functioning from plant traits: revisiting the Holy Grail[J]. Functional Ecology, 2002,16(5):545-556.
[14] 王震洪,段昌群,侯永平,等.植物多样性与生态系统土壤保持功能关系及其生态学意义[J].植物生态学报,2006,30(3):392-403.
[15] 郑颖,温仲明,宋光,等.延河流域森林草原区不同植物功能型适应策略及功能型物种数量随退耕年限的变化[J].生态学报,2015,35(17):5834-5845.
[16] Mason N W H, Mouillot D, Lee W G, et al. Functional richness, functional evenness and functional divergence: the primary components of functional diversity[J]. Oikos,2005,111(1):112-118.
[17] Micheli F, Halpern B S. Low functional redundancy in coastal marine assemblages[J]. Ecology Letters, 2005,8(8):391-400.
[18] Pillar V D, Duarte L S, Sosinski E E, et al. Discriminating trait-convergence and trait-divergence assembly patterns in ecological community gradients[J]. Journal of Vegetation Science, 2009,20(2):334-348.
[19] Grime J P. Trait convergence and trait divergence in herbaceous plant communities: mechanisms and consequences[J]. Journal of Vegetation Science, 2006,17(2):255-260.
[20] Cadotte M W, Carscadden K, Mirotchnick N. Beyond species: functional diversity and the maintenance of ecological processes and services[J]. Journal of Applied Ecology, 2011,48(5):1079-1087.
