不同栽培模式下冬小麦灌浆过程中旗叶蔗糖代谢和籽粒淀粉积累特性
2018-06-06李红兵邓西平宝鸡文理学院地理与环境学院陕西省灾害监测与机理模拟重点实验室陕西宝鸡71013中国科学院水利部水土保持研究所黄土高原土壤侵蚀与旱地农业国家重点实验室陕西杨凌71100
陈 炜,李红兵,邓西平(1.宝鸡文理学院 地理与环境学院,陕西省灾害监测与机理模拟重点实验室,陕西宝鸡 71013;.中国科学院 水利部水土保持研究所,黄土高原土壤侵蚀与旱地农业国家重点实验室,陕西杨凌 71100)
淀粉是小麦籽粒的主要储藏物质,占籽粒干质量的65%~70%,其组成和质量分数直接关系着小麦的加工品质[12]。小麦籽粒淀粉的形成与营养器官的生长状况密切相关。在籽粒淀粉合成的过程中,淀粉合成酶和淀粉分支酶均起重要作用[13],淀粉合成酶又分为可溶性淀粉合成酶(Soluble starch synthase,SSS)和淀粉粒结合态淀粉合成酶(Granule-Bound starch synthase,GBSS),前者主要催化合成支链淀粉,后者主要催化合成直链淀粉,而且GBSS需在淀粉粒形成一定的晶体结构以后才能起催化作用[14]。
国内外学者进行大量关于灌浆过程中旗叶蔗糖向籽粒淀粉合成转化的研究,但多是控制性或者单一因素试验,张艳艳等[15]分析不同土层深度测墒补灌对小麦旗叶蔗糖和籽粒合成的影响,表明0~40 cm的补灌深度可以显著提高旗叶磷酸蔗糖合成酶的活力和蔗糖质量分数,张礼军等[16]研究不同施肥和覆盖模式对旱地冬小麦糖代谢的影响,表明化肥和有机肥的共同使用可延缓灌浆前期旗叶蔗糖下降的速度,于淙超等[17]分析秸秆还田深度对小麦蔗糖转化及产量形成的影响,表明秸秆还田可以增加灌浆期间旗叶蔗糖的质量分数,张嫚等[18]分析不同温光型小麦品种花后旗叶生理与籽粒淀粉积累特性,Yang等[19]的研究结果表明,通过调控蔗糖向淀粉转化的酶的活性,增加灌浆速率,最终可以提高产量,同时蔗糖向淀粉转化相关酶的活性受到脱落酸(ABA)和赤霉素(GA)浓度的影响,曹雯梅等[20]研究表明,通过增加旗叶中蔗糖的输出,可以提高收获指数和氮肥生产效率。然而在小麦的实际生产中,往往受到多种因素的共同影响。生产实践中作物高产和高效是环境、栽培、耕作、管理等多因素综合作用的结果。因此,本试验根据长武地区生态条件,综合优化不同耕作、栽培等因素,研究不同栽培模式下冬小麦灌浆过程中叶片(源端)蔗糖代谢与转运和籽粒(库端)蔗糖-淀粉代谢相关酶活性等方面的特性,旨在探讨不同栽培模式对冬小麦灌浆过程中源器官(旗叶)和库器官(籽粒)中,蔗糖代谢和淀粉合成的生理机制差异,从作物生理学上解释不同栽培模式间籽粒淀粉积累的差异。
1 材料与方法
1.1 供试材料与试验设计
于2008-2009年在中国科学院长武农业生态试验站布置田间试验(107°40′E,35°12′N)。以当地小麦品种‘长武134’(2000年至今主要推广的品种)和‘长旱58’(新育成的品种,2004年审定至今)为试验材料,于2008年10月3日播种,2009年4月6日追肥,2009年6月21日收获,采用随机区组设计,随机区组排列,小区面积为136 m2,行距20 cm,4次重复。整个生育期无灌溉。2008年7月-2009年6月试验区总降雨量为494.4 mm,然而约63%的降雨发生在夏闲期(图1)。试验田土质为黑垆土,土壤密度平均为1.35 g·cm-3,耕层pH为8.4,有机质、全氮、速效磷和速效钾质量分数分别为11.8 g·kg-1、0.87 g·kg-1、14.4 mg·kg-1和144.6 mg·kg-1。试验采用4种不同的栽培模式,当地传统栽培模式(CM1):供试验品种为‘长武134’,氮肥水平为150 kg·hm-2,不施有机肥;优化栽培模式(CM2):供试验品种为‘长旱58’,氮肥水平为120 kg·hm-2,不施有机肥,并在拔节期追肥为75 kg·hm-2,进行保护性耕作处理;高产栽培模式(CM3):供试验品种‘长旱58’,氮肥水平为120 kg·hm-2,施有机肥(牛粪)45 000 kg·hm-2,拔节期追施氮肥为75 kg·hm-2,进行保护性耕作,其中有机肥碳、氮、钾和磷的质量分数分别为378.6、27.8、9.8和10.1 g·kg-1;高效栽培模式(CM4):供试验品种为‘长旱58’,氮肥水平为150 kg·hm-2,以农民正常耕作为主。上述栽培模式中磷肥水平均为120 kg·hm-2。选用上述4种栽培模式的原因是由于该地区降雨分布不均,通过适当的覆盖措施,使夏闲期的降雨存留在土壤中,施用有机肥从本质上改善土壤的养分状况和理化性质。
1.2 测定项目与方法
1.2.1 小麦旗叶可溶性糖、蔗糖质量分数及籽粒质量、淀粉质量分数 在开花期选取同一天开花、长势一致的单茎挂牌标记,每小标记250株主茎。自开花之日起,每隔6 d取 1 次标记植株样本的30株主茎,人工剥离籽粒,其中将20株主茎于105 ℃杀青30 min后,再在80 ℃烘干至恒量,旗叶粉碎后过孔径为0.25 mm的筛,用于可溶性糖、蔗糖和淀粉的测定。籽粒质量用百分之一天平测量,用于计算淀粉累积速率;另外10株经液氮速冻后置于-80 ℃冰箱,用于旗叶蔗糖代谢和籽粒淀粉积累相关酶活性的测定。可溶性糖质量分数的测定采用硫酸蒽酮法[21];淀粉质量分数测定参照蒽酮比色法[21];蔗糖质量分数的测定采用间苯二酚法[22]。
1.2.2 籽粒淀粉累积速率的计算 淀粉积累量=淀粉质量分数×粒质量,用Rn=(Cn+6-Cn-6)/12计算单粒淀粉积累速率,其中n为开花后天数,Rn为第n天的单粒淀粉积累速率,Cn+6为第(n+6)天的单粒淀粉积累量,Cn-6为第(n-6)天的单粒淀粉积累量。
1.2.3 旗叶蔗糖代谢相关酶和籽粒淀粉合成相关酶的活性 旗叶SPS和SS的活性测定:酶液的提取和测定参照汤章城等[23]的方法。
籽粒SSS和GBSS的活性测定:参照Douglas等[24]的方法。
1.3 数据分析
使用Microsoft Excel 2003处理数据并作图,采用SPSS 20.0统计软件进行差异显著性检验。
2 结果与分析
2.1 旗叶可溶性糖和蔗糖代谢
旗叶中可溶性糖(图2-A)和蔗糖(图2-B)质量分数的测定结果表明,灌浆过程中在不同栽培模式下,小麦旗叶中可溶性糖和蔗糖质量分数呈现先升高后逐渐降低的趋势。在开花后6~30 d旗叶中可溶性糖和蔗糖的质量分数逐渐上升,在花后24~30 d上升的速率最快,并于花后30 d达到高峰值后开始迅速下降。在小麦旗叶中,蔗糖的合成代谢比较活跃,并被运输至籽粒降解为淀粉合成前体,最终为合成淀粉奠定物质基础。不同栽培模式并没有改变试验条件下小麦花后旗叶中可溶性糖和蔗糖质量分数的变化趋势,‘长旱58’旗叶中可溶性糖和蔗糖的质量分数高于‘长武134’,表明在籽粒淀粉积累的旺盛期,‘长旱58’旗叶能合成更多的蔗糖以利于籽粒淀粉的合成,而在CM3中,由于有机肥和秸秆还田,提高土壤中的有机质,提高灌浆期可溶性糖和蔗糖的积累和运输能力,促进糖分向籽粒库运输,从而为籽粒灌浆提供充足的营养物质。
2.2 旗叶SPS和 SS活性
灌浆过程中,旗叶SPS(图3-A)和SS(图3-B)测定结果表明,灌浆期旗叶SPS和SS的活性随着灌浆过程的推移均呈现单峰曲线变化,各栽培模式均于花后24 d达到最大值,随后下降。SPS是叶片中蔗糖合成的关键酶,SPS活性的高低代表旗叶光合产物转化为蔗糖的能力,在籽粒灌浆中后期,‘长旱58’旗叶SPS活性显著高于‘长武134’,表明在籽粒淀粉积累的旺盛期,‘长旱58’蔗糖的供应能力强于‘长武134’。SS催化输送至小麦籽粒中的蔗糖分解为UDPG和果糖,是淀粉合成反应的第一步。同旗叶中SPS变化趋势一样,‘长旱58’旗叶SS的活力高于‘长武134’,表明在籽粒淀粉积累的旺盛期,‘长旱58’旗叶能合成更多的蔗糖以利于籽粒淀粉的合成。在‘长旱58’中,CM3中旗叶SPS和SS的活性最高,表明该栽培模式有利于提高旗叶SPS和SS的活性,说明通过相应的栽培措施维持其较高的酶活性,有利于后期光合产物向蔗糖和淀粉的转换。
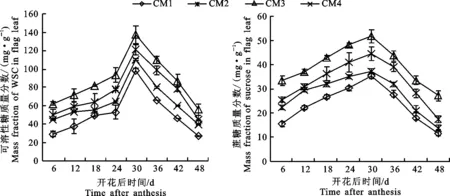
图2 不同栽培模式对灌浆过程中旗叶可溶性糖(A)和蔗糖(B)质量分数的影响
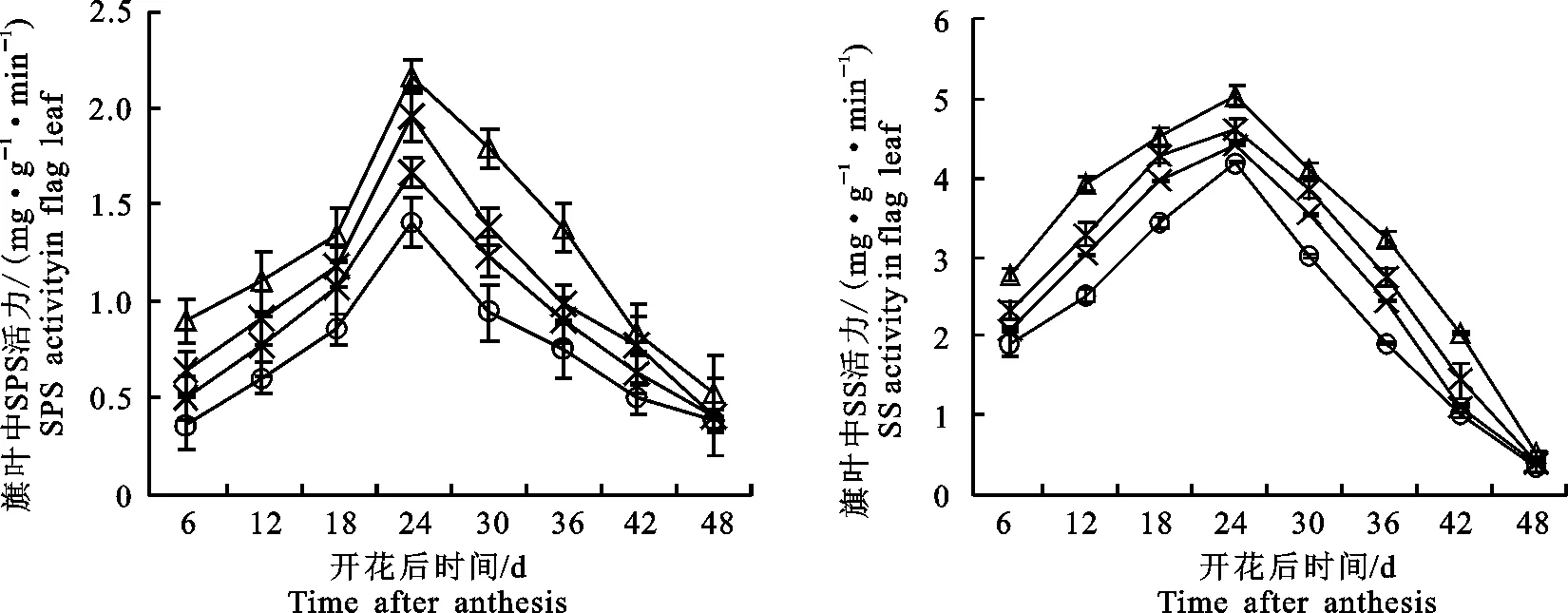
图3 不同栽培模式对灌浆过程中旗叶磷酸蔗糖合成酶(A)和蔗糖合成酶(B)活力的影响
2.3 籽粒淀粉积累量及其积累速率变化
灌浆过程中,每粒籽粒的淀粉累积量(图4)测定结果表明,淀粉可占小麦粒质量的70%左右,因此淀粉积累速率的快慢在很大程度上决定粒质量的高低。在籽粒的发育过程中,不同栽培模式下2个小麦品种籽粒淀粉积累量均呈逐渐升高的趋势。花后成熟时期籽粒淀粉积累量达到最大值。不同栽培模式对籽粒淀粉累积的影响表现为,‘长旱58’的淀粉最终积累量高于‘长武134’。在‘长旱58’中,CM3籽粒淀粉的累积量最大,其次为CM2,而CM4最低。在开花后前36 d,CM2和CM4淀粉累积量的差异较大,但其后无显著差异,说明CM2在淀粉合成高峰期提高淀粉累积量,而灌浆模式对淀粉累积量无显著影响;但CM3对淀粉累积量的影响持续整个灌浆过程。

图4 不同栽培模式对灌浆过程中籽粒淀粉积累量的影响
灌浆过程中每粒籽粒的总淀粉累积速率(图5)测定结果表明,随着灌浆过程的进行,籽粒总淀粉积累速率呈现出“慢-快-慢”的单峰变化趋势,灌浆初期,淀粉合成速率较低,可能是因为用于籽粒淀粉合成的“原料”供应不足,灌浆中期达到最大值,之后下降。在灌浆中期,源器官的光合作用较强,将光合同化产物以蔗糖的形式运输到籽粒中,而在灌浆后期,由于叶片衰老,旗叶中氮素的降解,影响光合作用,进而影响籽粒中淀粉的积累速率。不同栽培模式下,小麦总淀粉的积累变化趋势一致,均呈现出“低-高-低”的变化趋势,花后24 d,籽粒总淀粉积累速率达到最大。在不同栽培模式间,‘长旱58’籽粒总淀粉积累速率大于‘长武134’,CM3显著提高整个灌浆过程中籽粒总淀粉积累速率,而在花后18 d之前,CM2和CM4差异不显著,在花后18 d之后,差异逐渐显著。
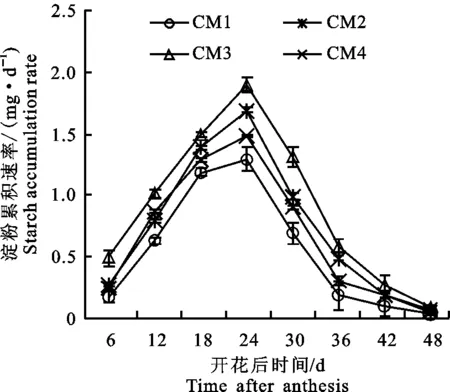
图5 不同栽培模式对灌浆过程中籽粒淀粉累积速率的影响
2.4 籽粒SSS和GBSS的活性
SSS主要存在造粉体中,主要催化支链淀粉的合成。在籽粒灌浆过程中,不同栽培模式下小麦籽粒SSS活性均呈单峰曲线,峰值出现在花后24 d(图6-A)。其中以CM3籽粒SSS的活性最高,表明该栽培模式促进小麦灌浆籽粒SSS活性的提高。‘长武134’籽粒SSS的活性低于‘长旱58’。GBSS也主要存在于造粉体中,它的活性必须在淀粉内部起作用,主要催化直链淀粉的合成。在籽粒灌浆过程中,不同栽培模式下籽粒GBSS活性变化与SSS活性相同,峰值也出现在花后24 d(图6-B)。
由表1可知,不同栽培模式显著影响冬小麦灌浆过程中旗叶蔗糖的代谢和籽粒淀粉的合成。‘长旱58’旗叶蔗糖合成和籽粒淀粉积累的能力显著高于‘长武134’。灌浆后期,不同栽培模式中籽粒淀粉累积速率差异不显著,而在灌浆中期,即旗叶蔗糖合成酶活力最大时,差异显著。
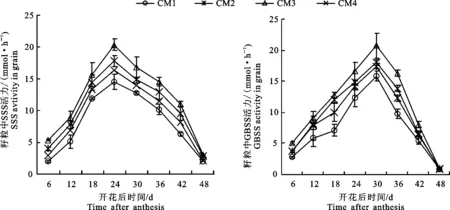
图6 不同栽培模式对灌浆过程中SSS(A)和GBSS(B)活性的影响
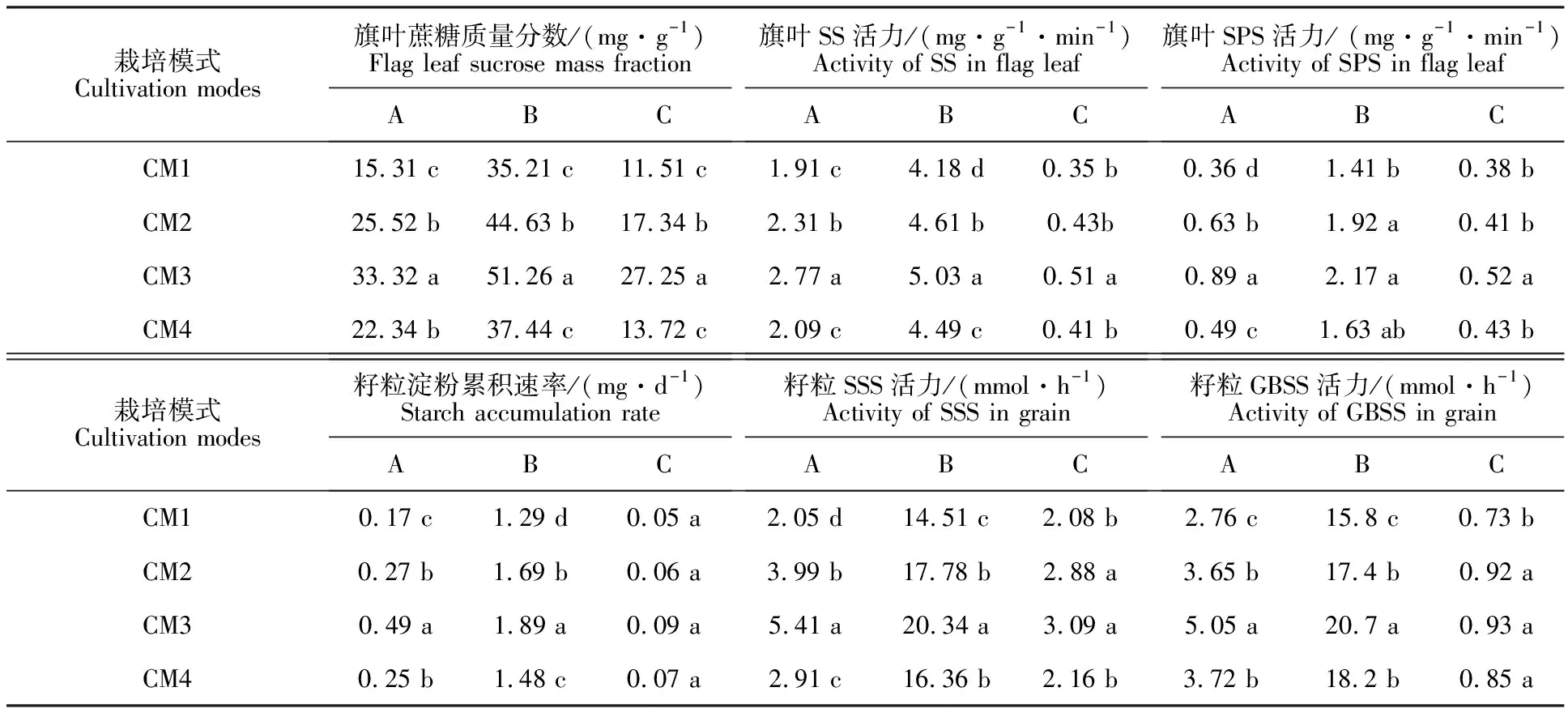
表1 旗叶蔗糖代谢和籽粒淀粉积累
注:A表示灌浆起始阶段,B表示最大值,C表示灌浆结束阶段。同列不同字母表示在0.05水平上差异显著。
Note:A. Initial grain filling stage; B. Maximum value during grain filling; C. The end of grain filling stage. Different letters within the same column indicate significant difference among the cultivation modes at 0.05 level.
3 讨 论
3.1 不同栽培模式对小麦灌浆过程中旗叶可溶性糖和蔗糖代谢的影响
花后光合产物主要以可溶性碳水化合物的形式存在于植物体内,以往研究表明,花后旗叶可溶性总糖的变化均为单峰曲线[25],本试验中,小麦花后旗叶可溶性糖质量分数也呈单峰曲线变化,花后0~6 d,可溶性碳水化合物和蔗糖的质量分数较低,这可能与此时穗部发育需要较多的光合产物,叶片可溶性碳水化合物和蔗糖的大量输出有关,同时与该阶段旗叶的光合作用较弱有关。花后6 d后开始回升,小麦开花后,花后光合产物是籽粒灌浆的主要物质来源,其中旗叶是主要光合器官,光合产物以蔗糖的形式向外输出,在供大于求的情况下引起旗叶蔗糖质量分数的升高。不同栽培模式下2 个小麦品种均于花后30 d达到高峰,随后迅速降低。可能是由于随着灌浆过程的持续,一方面籽粒合成淀粉对蔗糖的需求增加,另一方面由于叶片衰老,光合能力降低,导致旗叶中蔗糖质量分数降低,光合产物以蔗糖的形式向外输出。在CM3中旗叶蔗糖的质量分数和蔗糖合成的能力是最高的,可能是由于有机肥的使用,改善土壤的理化性质,增加土壤的有机质,增加源器官固定同化物的能力,同时延缓叶片的衰老,维持叶片的光合能力,从而提高整个灌浆过程中蔗糖合成的能力。因此延迟小麦光合器官的衰老,维持叶片足够的生理活性对于贮存光合产物向籽粒运输是至关重要的。SPS和SS是控制叶片中蔗糖合成的关键酶[26]。研究表明SPS在小麦叶片光合产物向蔗糖的转化过程中起关键性的调节作用[27]。同时SS在光合细胞蔗糖合成中也起重要作用[25]。CM3通过提高SPS和SS的活性进而提高旗叶中蔗糖的供应,即增加光合器官中光合产物的供应,这为获得高产提供物质基础。研究表明,增施氮肥,可以增加旗叶中蔗糖质量分数及旗叶蔗糖合成的能力[4]。本试验中,‘长旱58’的氮肥使用量为CM3>CM2>CM4,使蔗糖代谢关键酶的活性也存在CM3>CM2>CM4的规律,这与之前的研究结果一致[3,12,20]。
3.2 不同栽培模式对小麦灌浆过程中籽粒淀粉合成的影响
淀粉是小麦籽粒的主要贮藏物质,其合成与积累决定粒质量的高低[4],而籽粒淀粉是源器官制造的光合同化物以蔗糖的形式运输到籽粒后,在一系列酶的作用下形成的[28],在籽粒中与淀粉合成相关的酶包括可溶性淀粉合成酶(SSS)和结晶态淀粉合成酶(GBSS)[29]。淀粉的累积量与累积速率与源器官的供应能力和籽粒的库容密切相关。本试验中,不同栽培模式对淀粉合成关键酶的影响与对蔗糖代谢关键酶的影响一致,都是随施氮量的增加,小麦籽粒中淀粉合成关键酶活性均有所提高,这与前人的研究结果一致[30]。不同栽培模式没有影响上述酶活性在灌浆过程中的变化趋势,都是随着灌浆过程的进行,酶的活力呈现单峰变化的趋势,并且出现酶活性最大值的时间是一致的,在花后30 d。这可能与长武当地的气候有密切关系(高温和干旱)。由于灌浆过程中降雨较少,导致灌浆期提前结束,在灌浆结束前与蔗糖代谢和淀粉合成的相关酶活性迅速升高,其目的可能是使茎秆中的营养物质转运至籽粒以便快速形成淀粉粒。
小麦籽粒的充实过程,主要是淀粉的合成和积累过程。小麦淀粉积累速率主要由库强决定。库强取决于库容量和库活性2个方面,库容量一般包括胚乳细胞数目和大小等,而库活性则包括碳代谢相关酶的活性及其他生理指标[31]。灌浆前、中期籽粒中较高的蔗糖质量分数与较高的蔗糖合成酶活力有关,而灌浆后期较高的蔗糖质量分数由蔗糖转化为淀粉能力降低有关,本试验中,在整个灌浆过程中CM3不仅提高旗叶蔗糖合成酶活力而且增加蔗糖向淀粉转换的能力,可能是由于在CM3中有机肥的施用,增加土壤含水量,缓解由于土壤干旱对光合器官光合产物积累和淀粉合成关键酶的抑制作用,在一定程度上消除由于干旱对蔗糖转变为淀粉过程的抑制[32]。在本试验中,不同栽培模式主要通过影响“蔗糖-淀粉”代谢途径中关键酶的活性,实现对籽粒灌浆过程中源器官光合产物的供应能力和库器官淀粉合成能力进行调控,CM3中通过相应的农艺措施,能同时提高小麦源端同化物的供应能力和库端同化物的合成能力。
参考文献Reference:
[1] EVANS L J,DUNSTONE R L.Some physiological aspects of evolution of wheat[J].AustralianJournalofBiologicalScience,1970,23(4):725-742.
[2] STITT M,QUICK W P.Photosynthetic carbon partitioning:its regulation and possibilities for manipulation[J].PhysiologiaPlantarum,1989,77(4):633-641.
[3] 史 金,茹园园,谭金芳,等.施氮对冬小麦旗叶蔗糖含量及籽粒淀粉合成的影响[J].麦类作物学报,2007,27(3):497-502.
SHI J,RU Y Y,TAN J F,etal.Effects of nitrogen fertilizer application on sucrose content of flag leaf and starch synthesis in the grain of different winter wheat varieties[J].JournalofTriticeaeCrops,2007,27(3):497-502.
[4] 姜 东,于振文,李永庚,等.施氮水平对高产小麦蔗糖含量和光合产物分配及籽粒淀粉积累的影响[J].中国农业科学,2002,35(2):157-162.
JIANG D,YU ZH W,LI Y G,etal.Effects of different nitrogen application levels on changes of sucrose content in leaf,culm,grain and photosynthate distribution and grain starch accumulation of winter wheat[J].ScientiaAgriculturaSinica,2002,35(2):157-162.
[5] 潘庆民,韩兴国,白永飞,等.植物非结构性贮藏碳水化合物的生理生态学研究进展[J].植物学通报,2002,19(1):30-38.
PAN Q M,HAN X G,BAI Y F,etal.Advances in physiology and ecology studies on stored non-structure carbohydrates in plants[J].ChineseBulletinofBotany,2002,19(1):30-38.
[6] JIANG D,CAO W X,DAI T B,etal.Activities of key enzymes for starch synthesis in relation to growth of superior and inferior grains on winter wheat (TriticumaestivumL.) spike[J].PlantGrowthRegulation,2003,41(3):247-257.
[7] 李永庚,于振文,姜 东,等.冬小麦旗叶蔗糖和籽粒淀粉合成动态及其有关的酶活性的研究[J].作物学报,2001,27(5):658-664.
LI Y G,YU ZH W,JIANG D,etal.Studies on the dynamic changes of the synthesis of sucrose in the flag leaf and starch in the grain and related enzymes of high yielding wheat[J].ActaAgronomicaSinica,2001,27(5):658-664.
[8] ZHANG J J,XUY J,CHEN W,etal.A wheat 1-FEH w3 variant underlies enzyme activity for stem WSC remobilization to grain under drought[J].NewPhytologist,2015,205(1):293-305.
[9] CHARLES L G,JOAN L A H,STEVEN C H.Sucrosephosphate synthase and sucrose accumulation at low temperature[J].PlantPhysiology,1992,100(1):502-508.
[10] KEELING P L,WOOD J R,TYSON R H,etal.Starch biosynthesis in developing wheat grain[J].PlantPhysiology,1988,87(2):311-319.
[11] 李 咏,冶 婷,李卫华,等.小麦淀粉粒结合蛋白与面粉蛋白质和淀粉品质的关系[J].麦类作物学报,2016,36(5):603-610.
LI Y,YE T,LI W H,etal.Relationship between wheat starch granule protein with flour protein and starch qualities[J].JournalofTriticeaeCrops,2016,36(5):603-610.
[12] 潘庆民,于振文,王月福.小麦开花后旗叶中蔗糖合成与籽粒中蔗糖降解[J].植物生理与分子生物学学报,2003,28(3):235-240.
PAN Q M,YU ZH W,WANG Y F.Sucrose synthesis in flag leaves and sucrose degradation in grains after anthesis of wheat[J].JournalofPlantPhysiologyandMolecularBiology,2003,28(3):235-240.
[13] OU-LEE T M,SETTER T L.Effect of increased temperature in apical regions of maize ears on starchsynthesis enzymes and accumulation of sugar and starch[J].PlantPhysiology,1985,79(3):852-855.
[14] 王月福,于振文,李尚霞,等.小麦籽粒灌浆过程中有关淀粉合成酶的活性及其效应[J].作物学报,2003,29(1):75-81.
WANG Y F,YU ZH W,LI SH X,etal.Activity of enzymes related to starch synthesis and their effect during filling of winter wheat[J].ActaAgronomicaSinica,2003,29(1):75-81.
[15] 张艳艳,张永丽,石 玉,等.测墒补灌深度对小麦旗叶光合作用和产量的影响[J].麦类作物学报,2014,34(9):1233-1238.
ZHANG Y Y,ZHANG Y L,SHI Y,etal.Effects of supplemental irrigation based on measurement of soil moisture content on the photosynthesis and grain yield of winter wheat[J].JournalofTriticeaeCrops,2014,34(9):1233-1238.
[16] 张礼军,鲁清林,白玉龙,等.施肥和覆盖模式对旱地冬小麦花后干物质转移、糖代谢及其籽粒产量的影响[J].草业学报,2017,26(3):149-160.
ZHANG L J,LU Q L,BAI Y L,etal.Effects of different patterns of fertilization and mulching on the post-anthesis dry matter remobilization,sugar metabolism and grain yield of winter wheat in dry lands[J].ActaPrataculturaeSinica,2017,26(3):149-160.
[17] 于淙超,刘义国,林 琪.秸秆还田深度对小麦蔗糖转化及产量的影响[J].中国农学通报,2014,30(30):11-14.
YU C CH,LIU Y G,LIN Q.Effects of depth of straw return on sucrose content and yield of wheat[J].ChineseAgriculturalScienceBulletin,2014,30(30):11-14.
[18] 张 嫚,周苏玫,张甲元,等.不同温光型专用小麦品种花后旗叶生理与籽粒淀粉积累特性[J].麦类作物学报,2017,37(4):520-527.
ZHANG M,ZHOU S M,ZHANG J Y,etal.Physiological characteristics of flag leaf and starch accumulation in grains at post-anthesis of termo-photo sensitive wheat (TriticumaestivumL.) cultivars for different end uses[J].JournalofTriticeaeCrop,2017,37(4):520-527.
[19] YANG J C,ZHANG J H,WANG Z Q,etal.Activities of key enzymes in sucrose-to-starch conversion in wheat grains subjected to water deficit during grain filling[J].PlantPhysiology,2004,135(3):1621-1629.
[20] 曹雯梅,刘述忠,郑贝贝,等.不同高产栽培模式对小麦产量形成及灌浆期糖质量分数的影响[J].西北农业学报,2015,24(10):35-41.
CAO W M,LIU SH ZH,ZHENG B B,etal.Effects of high-yield cultivation patterns on yield and sugar mass fraction at grain-filling stage of winter wheat[J].ActaAgriculturaeBoreali-occidentalisSinica,2015,24(10):35-41.
[21] 高俊凤.植物生理学实验指导[M].北京:高等教育出版社,2006.
GAO J F.Guidance of Plant Physiology Experiment [M].Beijing:Higher Education Press,2006.
[22] 张志良,瞿伟菁.植物生理学实验指导 [M].北京:高等教育出版社,2003.
ZHANG ZH L,QU W J.Guidance of plant physiology experiment [M].Beijing:Higher Education Press,2003.
[23] 汤章城,魏家绵,陈 因,等.现代植物生理学实验指南 [M].北京:科学出版社,1999.
TANG ZH CH,WEI J M,CHEN Y,etal.Guide of Modern Plant Physiology Experiment [M].Beijing:Science Press,1999.
[24] DOUGLAS C D,TSUNG M K,FREDERICK C F.Enzymes of sucrose and hexose metabolism in developing kernels of two inbreds of maize[J].PlantPhysiology,1988,86(4):1013-1019.
[25] 姜 东,于振文,李永庚,等.冬小麦叶茎粒可溶性糖含量变化及其与籽粒淀粉积累的关系[J].麦类作物学报,2001,21(3):38-41.
JIANG D,YU ZH W,LI Y G,etal.Changes of soluble sugar content in leaf,stem and grain in winter wheat and its relationship with grain starch accumulation[J].JournalofTriticeaeCrops,2001,21(3):38-41.
[26] THOMASW R,PHIILLP S K,STEVEN C H.Characterization of diurnal changes in activities of involved in sucrose biosynthesis[J].PlantPhysiology,1983,73(2):428-433.
[27] 韩俊杰,王昊龙,李 咏,等.小麦淀粉合成酶基因SSIIa、SBEIIa和SBEIIb表达序列多态性及对淀粉质量分数的影响[J].西北农业学报,2016,25(8):1150-1157.
HAN J J,WANG H L,LI Y,etal.Correlation analysis between expression sequence polymorphisms of the genes encoding wheat starch synthesisSSIIa,SBEIIaandSBEIIb,and starch mass fraction[J].ActaAgriculturaeBoreali-occidentalisSinica,2016,25(8):1150-1157.
[28] 王文静,潘一展.不同类型小麦品种灌浆期蔗糖代谢关键酶的活性变化[J].华北农学报,2008,23(2):21-24.
WANG W J,PAN Y ZH.Dynamic changes of activities of key enzymes involved in sucrose metabolism during grain filling of wheat with different qualities[J].ActaAgriculturaeBorealiSinica,2008,23(2):21-24.
[29] NAKAMURA Y,YUKI K.Changes in enzyme activities associated with carbohydrate metabolism during development of rice endosperm[J].PlantScience,1992,82(1):15-20.
[30] WANG Z B,LI W H,QI J C,etal.Starch accumulation,activities of key enzyme and gene expression in starch synthesis of wheat endosperm with different starch content[J].JournalofFoodScienceandTechnology,2014,51(3):419-429.
[31] 王蔚华,郭文善,方明奎,等.小麦籽粒胚乳细胞增殖及物质充实动态[J].作物学报,2003,29(5):779-784.
WANG W H,GUO W SH,FANG M K,etal.Endosperm cell proliferating and grain filling dynamics in wheat[J].ActaAgronomicaSinica,2003,29(5):779-784.
[32] LINGLE S E,CHEVALIER P.Movement and metabolism of sucrose indeveloping barley kernels[J].CropScience,1984,24(2):315-319.
