低盐虾塘浮游生物群落及摄食研究
2018-05-29曾祥波黄永春
曾祥波,黄永春
(1.集美大学水产学院,福建 厦门 361021;2.鳗鲡现代产业技术教育部工程研究中心,福建 厦门 361021)
0 引言
浮游植物是虾塘水体浮游生物的主体,对维持虾塘水质和生态系统的稳定起着关键的作用,浮游植物种类组成及优势种群的数量往往决定了虾池环境的质量,是虾塘水环境调控的主要对象。浮游动物与虾塘水质、生态系统内的物质循环与能量流动关系密切,其对浮游植物的摄食压力同样会影响对虾的生长和系统的稳定。对虾塘浮游生物群落的研究,以往多侧重于浮游植物[1-4]或者浮游动物群落结构[5-6],而对整个浮游生物群落的研究仅见于个别报道[7-9]。
近年,应用稀释法研究微型浮游动物的摄食已从海洋[10-11]扩展至湖泊[12-13]、虾塘[14-15]等各种水生生态系统,而在低盐虾塘中有关微型浮游动物的摄食研究还未见报道。本文旨在通过对不同养殖阶段的虾塘浮游生物优势种类组成的调查以及微型浮游动物对浮游植物的摄食压力研究,为利用浮游生物调控虾塘水体环境以及虾塘生态系统的能流与物流研究提供基本资料和参数。
1 材料与方法
1.1 实验虾塘的基本情况
虾塘位于杏林浦林村,水深约 1.5 m,每亩虾塘2~3亩,养殖对象为凡纳滨对虾(Litopenaeusvannamei)。虾塘中、后期视水质变化适当换水,日常管理主要是定期泼洒益生菌等水质和底质改良剂。1号虾池为4月16日新引水培育的虾池,处于养殖的早期,3号虾池已捕大留小处于养殖的后期,4号和7号虾池分别养殖1个月和2个月左右,处于养殖的中期。2017年5月5日,分别采集水样用于后续实验研究。
1.2 浮游生物的采集与计数
在每个养殖塘四角及中央各取表层0.5 m处水样1份,混匀后,取1 L水样用酸性鲁哥氏液固定,终浓度为2%(体积分数),室内静置沉淀48 h以上,用底端附有10 μm筛绢的吸管逐步虹吸浓缩至30 mL,甲醛保存。将样品摇匀,取0.1 mL置于计数框中,在显微镜下进行种类的鉴定与计数。
1.3 虾塘微型浮游动物摄食压力实验
微型浮游动物的摄食压力通过稀释法[10]进行,实验主要步骤为:如上取各虾塘表层0.5 m处的养殖水,用孔径为 0.45 μm的Whatman GF/F 玻璃纤维滤膜过滤,制备12 L稀释液。然后将稀释液按0 ∶1、1 ∶3、1 ∶1、3 ∶1等4个比例分别与200 μm 筛绢过滤的虾塘养殖水现场混合,分装于1.5 L 聚碳酸酯培养瓶中,每个比例设两个平行样,置于虾塘水面下0.5 m处,培养 24 h。取采集培养前后的水样各100 mL,用0.45 μm 的 Whatman GF/F滤膜过滤,滤膜对折,加10 mL 90%(体积分数) 丙酮-20 ℃萃取24 h,采用荧光光度法测定水样的叶绿素a浓度。
根据稀释法原理,用线形回归模型求得浮游植物的生长率k(d-1)和微型浮游动物的摄食率g(d-1),微型浮游动物对浮游植物现存量及初级生产力的摄食压力(分别用Pi和Pp表示)用下列公式计算[16]:
Pi(%)=1- e-gt×100,
PP(%)=(ekt- e(k-g)t)/(ekt-1)×100,
其中Pt是t时浮游植物的浓度,Po为起始时浮游植物的浓度,k是浮游植物的生长率,g是浮游动物的摄食率,d为稀释度。
2 结果
2.1 浮游生物种类组成
虾塘浮游植物优势类群主要为绿藻和蓝藻,其次为硅藻和裸藻,总丰度为1.056×107~1.512×108个/L(见表1)。各塘浮游植物构成及优势种类有较大的差异:1号池种类较多,包括多种绿藻、硅藻,优势蓝藻只有一种;4号池虽然有多个门的藻类,但各门藻类优势种类减少,丰度及优势度却明显增加,如绿藻主要为二形栅藻(Scenedesmudimorphus),而蓝藻主要为鞘丝藻(Planktolyngbyasp.)及微小平裂藻(Merismopediatenuissima);3号池则以多种蓝藻为优势类群,主要为伪鱼腥藻(Pseudanabaenasp.)、鞘丝藻(Planktolyngbyasp.)、弯形尖头藻(Raphidiopsiscurvata)和弱细颤藻(Oscillatoriatenuis);7号池由于中途蓝藻爆发,施以杀藻剂,重新调控培育后则以绿藻为优势类群。整体上,随着养殖的进行,浮游植物群落从优势类群多样,以绿藻和硅藻为主,逐渐转变为以蓝藻尤其是丝状蓝藻为主的类型(见表 1)。
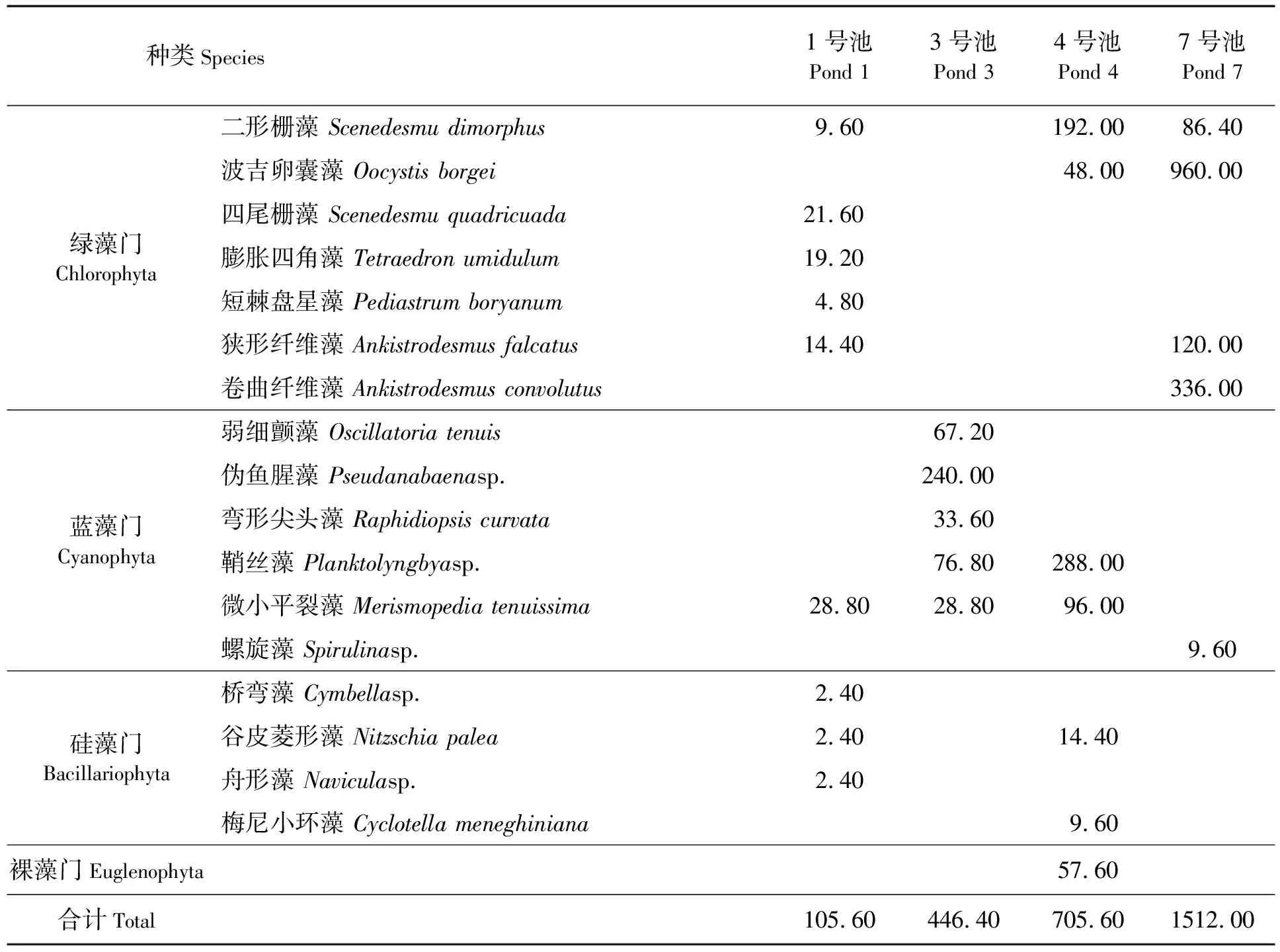
表1 浮游植物优势种类组成及丰度
微型浮游动物总丰度为1.02×105~7.20×105个/L,主要为原生动物和轮虫。其中:3号池主要为轮虫,而其他池均以原生动物为主,旋急游虫(Strombidiumspiralis)和单环栉毛虫(Didiniumbalbianii)为普遍的优势种类;1号池、4号池以及7号池则还分别以膜袋虫(Cyclidiumsp.)、游仆虫(Euplotessp.)、筒壳虫(Tintinnidiumsp.)为优势种类,轮虫主要为前额犀轮虫(Rhinoglenafrontalis)和壶状臂尾轮虫(Brachionusurceus)(见表2)。
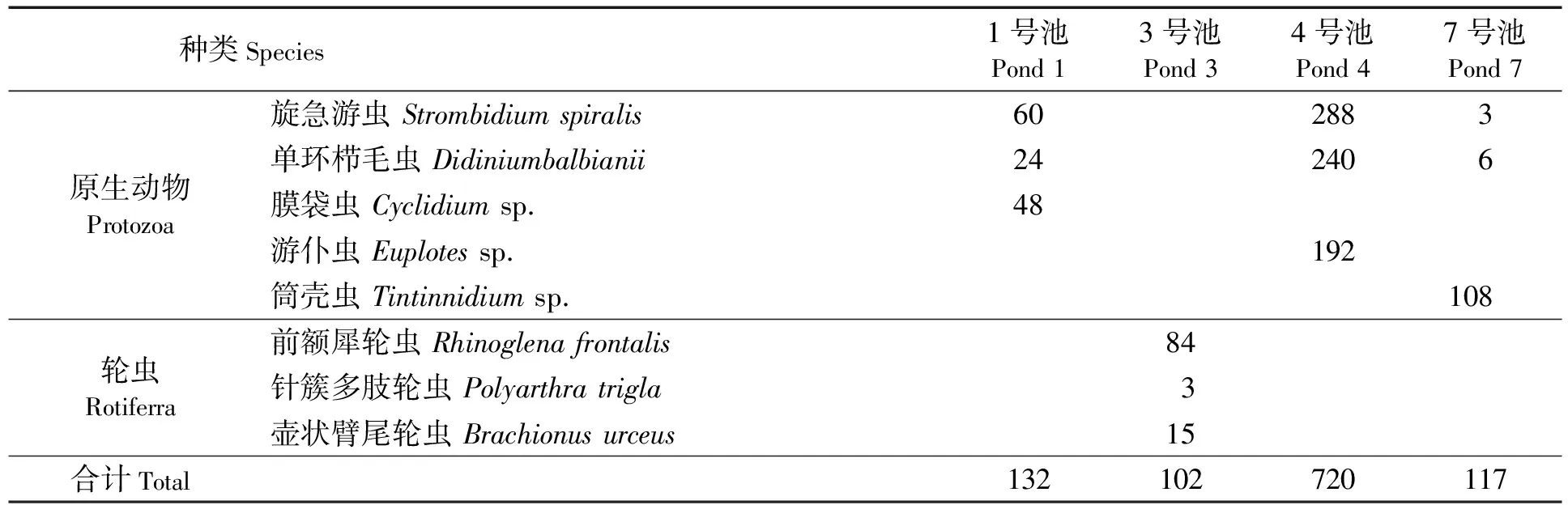
表2 微型浮游动物优势种类组成及丰度
2.2 摄食压力实验结果
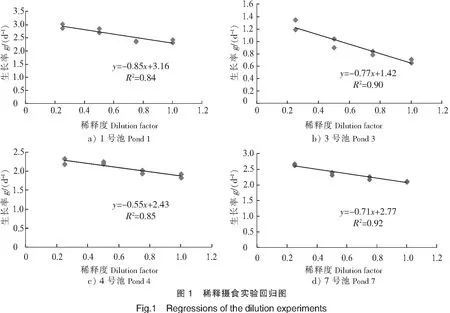
实验结果如表3与图1所示。微型浮游动物对浮游植物的摄食率为0.55~0.85/d,浮游植物的生长率为1.42~3.16/d,微型浮游动物对浮游植物现存量的摄食压力为42.31%~57.26%,对浮游植物初级生产力的摄食压力为46.39%~59.80%。
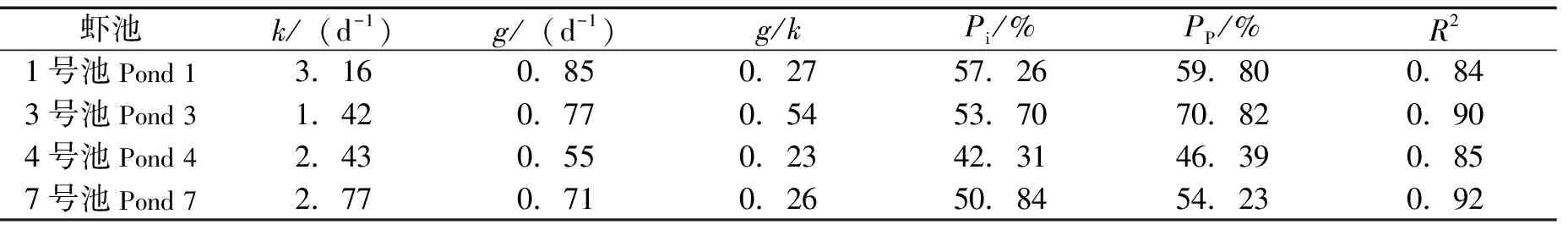
表3 微型浮游动物对浮游植物的摄食压力
3 讨论
3.1 虾塘浮游生物群落结构
查广才等[7]对广东淡化虾塘微型浮游生物群落研究表明,对虾淡化低盐养殖水体中的浮游植物,基本为淡水种类,主要以绿藻和蓝藻为主,且种类组成与对虾海水养殖或沿海海域种类组成不同,绿藻主要为栅藻和小球藻(Chlorellavulgaris)等,蓝藻主要为螺旋藻(Spirulinasp.)、伪鱼腥藻(Pseudoanabaenasp.)、弯形尖头藻(Raphidiopsiscurvata)等。陈辉煌[8]在浙江省凡纳滨对虾低盐综合养殖池塘浮游生物群落结构的研究中也有类似的结果,主要优势种为螺旋藻(Spirulinasp.)、湖泊鞘丝藻(Planktolyngbyalimnetica)和湖生卵囊藻(Oocystislacustris)等。本研究的结果表明,优势种类并没有与前两者完全一致,而更倾向是两地优势种类的综合结果(见表1),且处于不同养殖阶段的虾塘其优势种类之间表现出较大的差异。这进一步说明了,虾塘中的浮游植物种类除主要受到盐度的影响以外,还与虾塘底质、所处水温和pH等环境因素紧密相关[17]。
低盐虾塘中的微型浮游动物主要为原生动物[7]或者原生动物和轮虫[8].本研究结果表明,不同养殖阶段、不同虾塘,微型浮游动物的种类及组成差别较大。在养殖过程中由于投饵导致营养盐不断累积,使浮游植物大量增殖,如以浮游藻类或有机碎屑为食的纤毛虫等原生动物得以大量繁殖,使其在种类和数量上占据优势。但研究[7]表明,对虾的养殖密度、放苗时间、饵料的投喂量、施肥及杀菌等消毒药物的应用以及天气变化等各种因素都可影响到水体中微型浮游动物种类组成及变化,因此,不同虾池或同一虾池不同养殖时间,其微型浮游动物种类和个体数量都有所差异,有时差异比较大。本次研究结果则与此相一致。
3.2 虾塘微型浮游动物对浮游植物的摄食
根据不同研究水域微型浮游动物的摄食结果来看,浮游植物生长率为-0.63~3.30/d,微型浮游动物摄食率为 0~3.86/d,微型浮游动物对浮游植物现存量的摄食压力为11%~811.69%,对初级生产力的摄食压力为0.20%~467.88%[13]。本研究与其相比,虾塘微型浮游动物摄食结果均处于这些范围之内;但浮游植物生长率处于较高的水平,尤其1号池和7号池,这与其处于养殖培育早期,水体营养盐丰富,水质条件良好,浮游植物大量繁殖的实际相一致。
张立通等[13]对虾塘养殖中后期微型浮游动物的摄食压力的研究表明,≤20 μm微型浮游动物的摄食率、微型浮游动物对浮游植物现存量和初级生产力的摄食压力,占微型浮游动物(≤200 μm) 摄食率及相关摄食压力的比例分别为73.85%~97.69%、76.67%~97.91%和78.87%~98.59%,说明≤20 μm微型浮游动物在虾塘生态系统中起到了绝对重要的作用,而这主要是因为该虾塘的浮游植物主要以≤20 μm微型浮游植物为主,其生物量占微型浮游植物生物量(≤200 μm)的90%以上。而从微型浮游动物的组成及其食性来看,除了单环栉毛虫是肉食者,其他原生动物和轮虫主要属于食藻者和食菌的碎屑者[7]。本次实验期间,各虾塘的浮游藻类也基本以微藻为主,根据粒经谱理论,这些优势微藻应不易被≤20 μm微型浮游动物所摄食,微型浮游动物(≤200 μm)才是其主要的摄食者,但由于虾塘饵料非常丰富,浮游藻类生长率高,微型浮游动物摄食率对浮游植物生长率比值(g∶k)介于0.23~0.54之间,微型浮游动物对浮游植物的摄食不足以控制其种群的增长,这也是虾塘易暴发藻华的原因之一。
通过本次研究表明,虾塘由于受人为调控的影响,浮游生物群落优势类群变化快,浮游植物生长率高,浮游植物与微型浮游动物关系密切。弄清虾池生态系统的物质循环和能量流动的过程与机理还需进一步的深入研究,类似张立通等[15]、陈默等[13]对浮游生物分粒级研究的方法是可以发展的一个方向。
[]
[1]曾建刚,蒋霞敏.对虾养殖塘浮游植物的动态变化[J].海洋湖沼通报,2010(1):71-81.
[2]彭聪聪,李卓佳,曹煜成,等.凡纳滨对虾半集约化养殖池塘浮游微藻优势种变动规律及其对养殖环境的影响[J].海洋环境科学,2011,30(2):193-198.
[3]查广才,麦雄伟,周昌清,等.凡纳滨对虾低盐度养殖池浮游藻类群落研究[J].海洋水产研究,2006,27(1):1-7.
[4]李由明,黄翔鹄,李晓梅.凡纳滨对虾养殖水体中浮游植物群落的组成分析[J].琼州学院学报,2012,19(5):42-45.
[5]晏军,张玉平,孙振中,等.鱼、虾和蟹塘浮游甲壳动物群落结构分析与讨论[J].水产科技情报,2015,42(4):169-174.
[6]吴礼广,王伟洪,宋飞彪,等.南美白对虾大棚养殖池浮游动物的种群动态与调控研究[J].农村经济与科技,2013,24(4):165-166.
[7]查广才,周昌清,黄建荣,等.凡纳对虾淡化养殖虾池微型浮游生物群落及多样性[J].生态学报,2004,2(8):1752-1759.
[8]陈辉煌.凡纳滨对虾低盐综合养殖池塘浮游生物群落结构的研究[D].宁波:宁波大学,2013.
[9]何京,陈晨,王一农,等.凡纳滨对虾设施养殖池塘浮游生物群落结构及多样性研究[J].生物学杂志,2015,32(3):58-62.
[10]LANDRY M R,HASSETT R P.Estimating the grazing impact of marine microzooplankton[J].Mar Biol,1982,67:283- 288.
[11]CALBET A,LANDRY M R.Phytoplankton growth,microzooplankton grazing,and carbon cycling in marine systems[J].Limnology and Oceanography ,2004,49:51-57.
[12]段翠兰,陈 静,张永江,等.大型浅水湖泊太湖中微型浮游动物的摄食生态研究[J].安徽科技学院学报,2016,30(3):44 -49.
[13]陈默,高光,朱丽萍,等.太湖水体中微型原生动物对细菌的捕食作用[J].应用生态学报,2007,18(10):2384-2388.
[14]沈锦兰,林元烧,杨圣云,等.厦门杏林虾池夏冬季微型浮游动物对浮游植物的摄食压力[J].台湾海峡,2002(1):31-36.
[15]张立通,孙耀,赵从明,等.虾塘养殖中后期微型浮游动物的摄食压力[J].生态学报,2011,31(7):2046-2052.
[16]VERITY P G,STOECKER D K,SIERACKI M E ,et al.Grazing,growth and mortality of microzooplankton during the 1989 North Atlantic spring bloom at 47 °N,18 °W[J].Deep Sea Res I,1993,40:1793-1814.
[17]彭聪聪,李卓佳,曹煜成,等.虾池浮游微藻与养殖水环境调控的研究概况[J].南方水产,2010(5):74-80.
