南极普里兹湾夏季冰间湖沉降通量
2018-05-26韩正兵孙维萍范高晶扈传昱潘建明张海峰张海生
韩正兵,孙维萍,范高晶,扈传昱,潘建明*,赵 军,张海峰,李 栋,张海生
南极普里兹湾夏季冰间湖沉降通量
韩正兵1,2,3,孙维萍2,3,范高晶1,2,3,扈传昱2,3,潘建明2,3*,赵 军2,3,张海峰2,3,李 栋2,3,张海生1,2,3
(1.中国地质大学海洋学院,湖北 武汉 430074;2.国家海洋局,海洋生态系统与生物地球化学重点实验室,浙江 杭州 310012;3.国家海洋局第二海洋研究所,浙江 杭州 310012)
为研究南大洋的生物泵,利用2009~2010年和2014~2015年中国第26、31次中国南极科学考察期间于普里兹湾冰间湖布放的时间序列沉积物捕获器,获取了夏季沉降颗粒物通量并分析了其组成成分.结果表明: 2009~2010年和2014~2015年南极夏季普里兹湾颗粒有机碳平均通量分别为4088.13,508.99μmol/(m2·d),生物硅则分别为7358.91,2034.63μmol/(m2·d),其中生物蛋白石占夏季颗粒物通量均超过70%,表明硅藻是普里兹湾夏季的优势种和沉降通量的主要贡献者.2014~2015年有机碳沉降通量仅为2009~2010年12.5%,主要是由于缺少压舱物无法使上层有机质发生快速沉降,造成异养微生物对上层水柱中有机质降解程度更高,进一步影响有机碳的沉降效率.与非硅藻优势种的海域相比,夏季普里兹湾具有非常高的沉降通量和沉降效率,利用Martin曲线估算夏季普里兹湾的100m深度沉降通量平均为净初级生产力的8.67%,主要归因于硅藻的高沉降效率.为了预测南极海域生物泵的变化,需要重点关注南大洋浮游植物群落结构的组成与变化.
沉降通量;冰间湖;普里兹湾;生物泵
生物泵是碳从上层海洋向海洋中深层输送的重要方式,主要是通过浮游植物光合作用固定大气中的CO2并合成有机质,经过一系列复杂的生物化学作用和物理过程,最终向海洋深处输送的一个过程[1-2].极地海域是全球变化最敏感的区域之一[3],在全球变暖的背景下发生着显著的环境变化,如海冰覆盖减少、表层海水淡化、生物泵作用增强等.研究表明,南大洋生物泵效率比大多数中、低纬度海域高,这意味着南极海域具有活跃的碳循环过程[1].在海冰快速变化的条件下,生物泵在南大洋生态系统中的作用,南大洋生物泵的运转和效率受到的影响,在国际上引起了广泛的关注[4-5].
罗斯海、威德尔海和普里兹湾等南极边缘海,是南大洋初级生产的关键海域,在南极夏季海冰融化、上升流等过程的影响下,其浮游植物群落结构发生快速的演替[6-7],因而是研究南大洋生物泵变化趋势和机制的理想海域.相较于罗斯海[5,8-11]、威德尔海(含南极半岛海域)[12-16]、阿蒙森海[4,17]等南极边缘海已有的大量研究,位于南大洋印度洋扇区的普里兹湾观测相对偏少,仅有Pilskaln等[18]对普里兹湾北部深海区(~62°S)沉降通量进行的研究.
普里兹湾是东南极的最大海湾,是仅次于罗斯海和威德尔海的南极第3大边缘海,包括东南极初级生产力最高的冰间湖区域,其年均初级生产力可达103.9g C/m2,此区域的高生产力为东南极最大的阿德雷企鹅栖息地提供了关键的食物基础[19].上层海洋浮游植物通过光合作用所合成的大量有机质,一部分通过食物链的传递满足高营养级生物的物质需求,另一部分则通过沉降至海洋深处以降低大气中CO2,因此,深入研究普里兹湾沉降通量变化与生物泵过程具有重要意义.
本研究利用2009~2010年、2014~2015年夏季中国第26、31次中国南极科学考察期间于普里兹湾冰间湖获取的沉积物捕获器样品,通过对颗粒物中各生物地球化学要素沉降通量的测定,分析夏季沉降颗粒物通量及其组成成分,了解其年际变动,并与南极其他边缘海沉降通量进行对比,以期为南大洋生物泵过程以及对全球碳循环研究提供新的认识.
1 材料与方法
1.1 沉积物捕获器的布放和样品获取
沉积物捕获器是研究沉降通量时间序列的重要手段[2],本文利用2009~2010年和2014~2015年南极夏季期间,在普里兹湾冰间湖区域布放沉积物捕获器(PARFLUX Mark78H-21型,McLane, Inc., USA),获取普里兹湾夏季时间序列的沉降颗粒物样品.2009~2010年和2014~2015年沉积物捕获器布放站位分别为M26(75.53°E,68.52°S,站位水深650m,沉积物捕获器深度为480m)和M31(75.38°E,68.49°S,站位水深620m,沉积物捕获器深度为390m)(图1),沉积物捕获器的捕获面积为0.5m2,样品瓶体积为500mL,在样品瓶中注满预先原位过滤的海水,沉积物捕获器中加入氯化汞溶液达到饱和浓度(1.67%,g/g),以防止有机质由于微生物活动而发生降解,采样时间、周期等详细信息见表1.
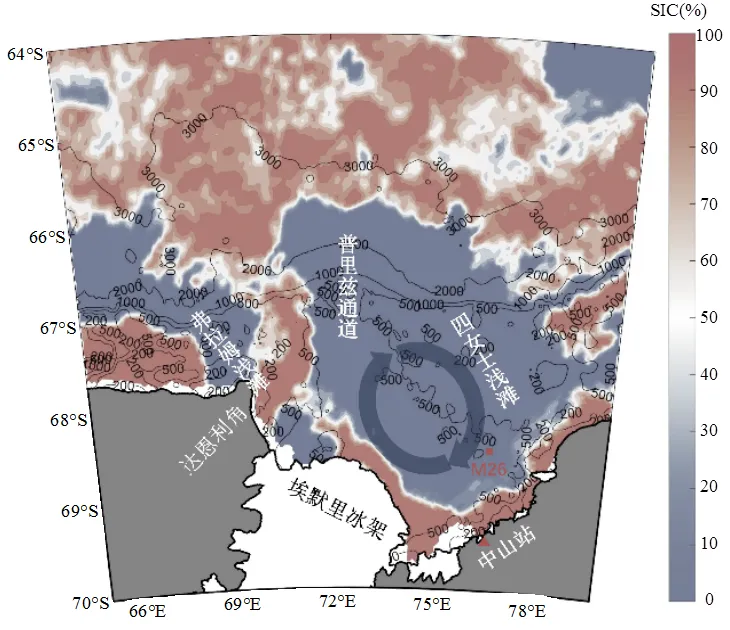
图1 2009~2010年和2014~2015南极夏季普里兹湾潜标布放站位
黑色等值线为水深等值线.填充色为2009年12月7日海冰密集度(SIC)分布,数据来自于德国不莱梅大学的Polar View of Bremen数据库.阴影部分表示普里兹湾顺时针环流(Prydz Gyre)
1.2 分析方法
收集样品后,立即于4℃进行冷藏保存.通过筛绢(孔径为1mm)以去除浮游动物,然后将小于1mm的样品采用旋转分样器(WSD-10,McLane, Inc., USA)均匀的分为10份.其中一份采用称量法获得总物质通量(TMF),样品通过预先已称量的聚碳酸酯膜(Millipore,孔径为0.4μm),用去离子水洗去盐分,在45℃干燥后称重,利用差减法计算出单份的物质重量,通过以下公式计算:
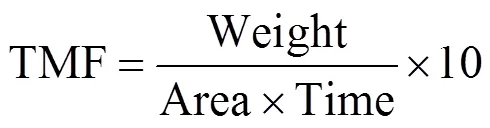
式中:Weight为单份沉降颗粒物的质量;Area为沉积物捕获器的捕获面积,0.5m2;Time为每份样品的采样天数.
颗粒有机碳(POC)和颗粒有机氮(PON)的分析:利用0.7μm孔径的玻璃纤维膜(预先用450℃灼烧5h)过滤所获取的样品,然后用盛有浓盐酸的干燥器进行酸熏处理12h,再使用超纯水洗去膜上的酸,并在45℃干燥,最后利用元素分析仪(vario MICRO cube,Elementar,German)进行分析[20].总碳则将样品过滤至0.7μm孔径的玻璃纤维膜(预先用450℃灼烧5h),直接45℃烘干并使用元素分析仪进行测定.颗粒无机碳(PIC)通过总碳和颗粒有机碳的差值法计算得出.
生物硅(BSi)的分析:采用碳酸钠提取的方法进行测定[21],利用H2O2和HCl对样品进行预处理分别去除有机质和碳酸盐,然后用2mol/L的Na2CO3溶液在85℃恒温水浴中加热提取,提取时间为5h.提取液中硅酸盐含量采用硅钼黄法,于波长380nm处采用分光光度计进行测定.并根据生物硅的含量乘以总物质通量计算其通量.蛋白石(Opal)的计算采用SiO2·0.4H2O表达[17-18].
铝(Al)、铁(Fe)和钛(Ti)的分析方法是依据美国环境保护署规范方法USEPA Method 3052[22],样品用3mL HF和7mL HNO3的混合酸经密闭微波消解系统(CEM Mars,USA)消解后,采用ICP-MS(Angilent 7000,USA)进行分析测定,相对标准偏差小于8.9%[23].
δ13C利用酸熏处理后的GF/F膜样(处理方法同POC),使用稳定同位素比值质谱仪(Thermo Delta V advantage,German)分析,标准物质为国际标准维也纳PeeDee箭石(VPDB),为确保仪分析过程中的稳定性,每10个样品加入1个IAEA- CH3标准物质(δ13C=-24.724‰)进行单点校正, IAEA-CH3标准物质的测定相对标准偏差<0.2‰.δ13C的计算公式为:

1.3 遥感叶绿素
遥感叶绿素浓度来自Aqua-MODIS产品,其空间分辨率为4km,时间分辨率为8d,选取以观测站位为中心的60km×60km范围的数据进行讨论分析.本文使用的遥感数据均已进行均值计算.
2 结果与讨论
2.1 观测站位的环境特征
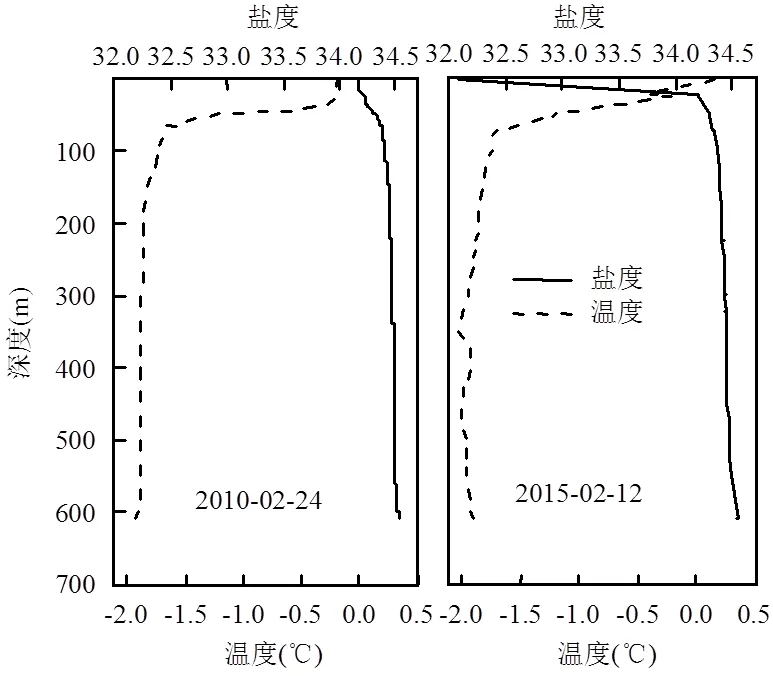
图2 2010年2月24日(左)和2015年2月12日(右)潜标站位温度、盐度垂直剖面
由于受到船时、冰情等条件的限制,不同航次仅对潜标站位进行了一次观测温盐剖面观测.2010年2月24日和2015年2月12日所观测到的温盐垂直剖面变化如图2.两年所观测的上层海水中温度、盐度具有夏季表层水(温度大于-1.8℃且小于2.1℃;盐度大于30.6且小于34.2)[24]的特征,主要受控于海冰消融和夏季太阳辐射的增强.随着深度的增加,温度跃层和盐度跃层同时出现,水团逐渐变为冬季水(温度大于-1.9℃且小于-1.5℃;盐度大于34.3且小于34.5)[24],属于典型的普里兹湾陆架水柱温盐剖面.海洋上层盐度的变化趋势主要反映了海冰消融程度的差异, 2015年2月12日观测到上层盐度较低主要是由于海冰消融造成的.
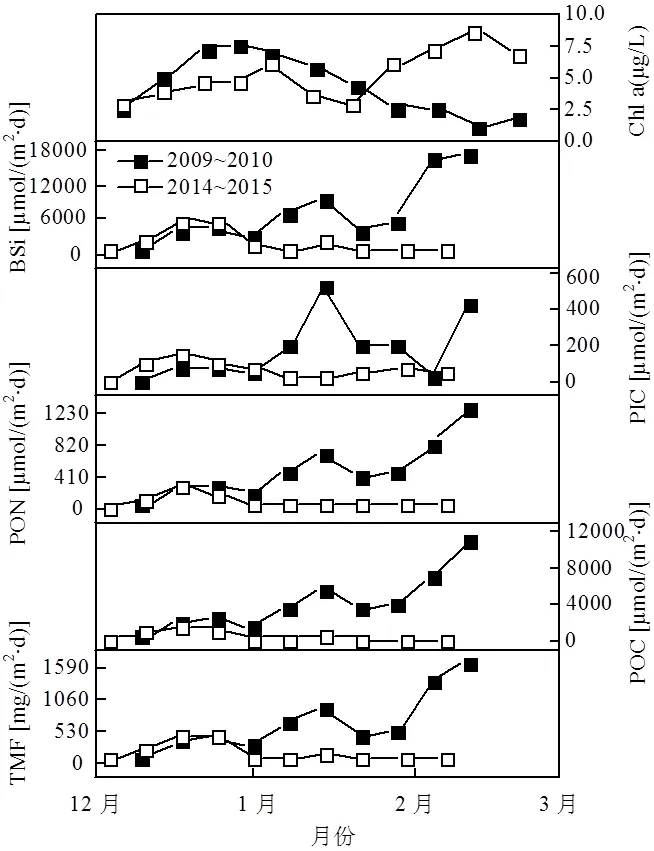
图3 2009~2010年和2014~2015年南极夏季普里兹湾各生源组分沉降通量时间序列
2.2 颗粒物沉降通量及组成
由图3和表1可见,2009~2010年和2014~ 2015年南极夏季普里兹湾TMF分别为672.61, 158.39mg/(m2·d),2014~2015年仅为2009~2010年的23.5%.两年的变化趋势也有所不同,2009~ 2010年夏季初较低,随着时间的推移,呈逐渐增高的趋势,并于次年2月份达到最高. 2014~2015年夏季初趋势同2009~2010年相似,但在次年1月和2月并未呈现明显的升高趋势.
POC是上层浮游植物初级生产的重要指标,其通量变化反映了有机质从水柱上层向下的输出变动. 2009~2010年和2014~2015年POC夏季通量均值为4088.13、508.99μmol/(m2·d)(表1), 2014~2015年仅为夏季POC通量仅为2009~2010年夏季的12.5%.整体的变化趋势与各年份的TMF的趋势相似(图3),2009~2010、2014~2015年的POC通量分别占夏季颗粒物总通量的7.3%和3.9%(图4,图5),约为普里兹湾湾外POC所占比重的1~2倍[18],反应了普里兹湾的高生产力特征.PON的变化趋势与POC完全一致,两年夏季通量分别为486.44、85.01μmol/(m2·d).
BSi是南极边缘海最重要的生源要素[2],是硅藻初级生产的重要指标.2009~2010年和2014~ 2015年夏季通量分别为7358.91、2034.63μmol/ (m2·d)(表1),折算为生物蛋白石(以SiO2·0.4H2O表达),占TMF的73.5%和86.3%,与湾外1000m深度的贡献相近(超过70%)[18],是沉降通量的主要贡献者(图4,图5).两年夏季的POC和BSi通量具有显著的正相关(2=0.896,<0.01),表明硅藻是普里兹湾夏季的主要初级生产者,与之前水柱中浮游植物[25-27]和沉积物[28]中的报道相符.
2009~2010年和2014~2015年PIC的夏季通量为179.04、65.52μmol/(m2·d)(表1),其平均通量为POC的4.4%和12.9%.以碳酸钙计,分别占TMF的2.7%和4.1%(图4,图5).2010年1月中旬和2月中旬出现高值(图3),这部分的贡献可能是来源于翼足类的文石碳酸盐壳体和浮游有孔虫等方解石碳酸盐壳体[2,8,18,29],湾外的浮游有孔虫所贡献的碳酸盐通量峰值也出现在1月中下旬[18],与湾内冰间湖的PIC通量峰值呈现了较好的同步性.2014~2015年夏季则没有明显的峰值现象,仅在夏初时期出现较小的峰值.
铝和钛是岩源物质(Litho.)的重要成分,可以反映岩源性颗粒物的来源[8],其比值可以用来指示南极海洋沉降颗粒物中岩源物质[8,18,23].2009~ 2010年Al和Ti的夏季沉降通量分别为121.67、4.78μmol/(m2·d).2014~2015年为19.64、0.76μmol/ (m2·d)(表1,图6),分别为2009~2010年的16.1%和15.9%.Al、Ti的变化趋势一致,存在显著正相关关系(图7,2>0.9,<0.01).两年夏季Al和Ti通量的质量比分别为14.3和14.6,与地壳中Al/Ti比值14.4相近[30],岩源特征明显,表明Al或Ti的通量可直接反映岩源通量的大小.以Al含量的12.15倍[10,18],估算2009~2010年和2014~2015年岩源性物质通量分别为39.91、6.44 mg/(m2·d),远高于湾外的岩源物质通量[18],推测冰筏碎屑、风尘、沉积物的再悬浮或侧向输送可能是南极陆架区域岩源物质的重要来源[8,23].
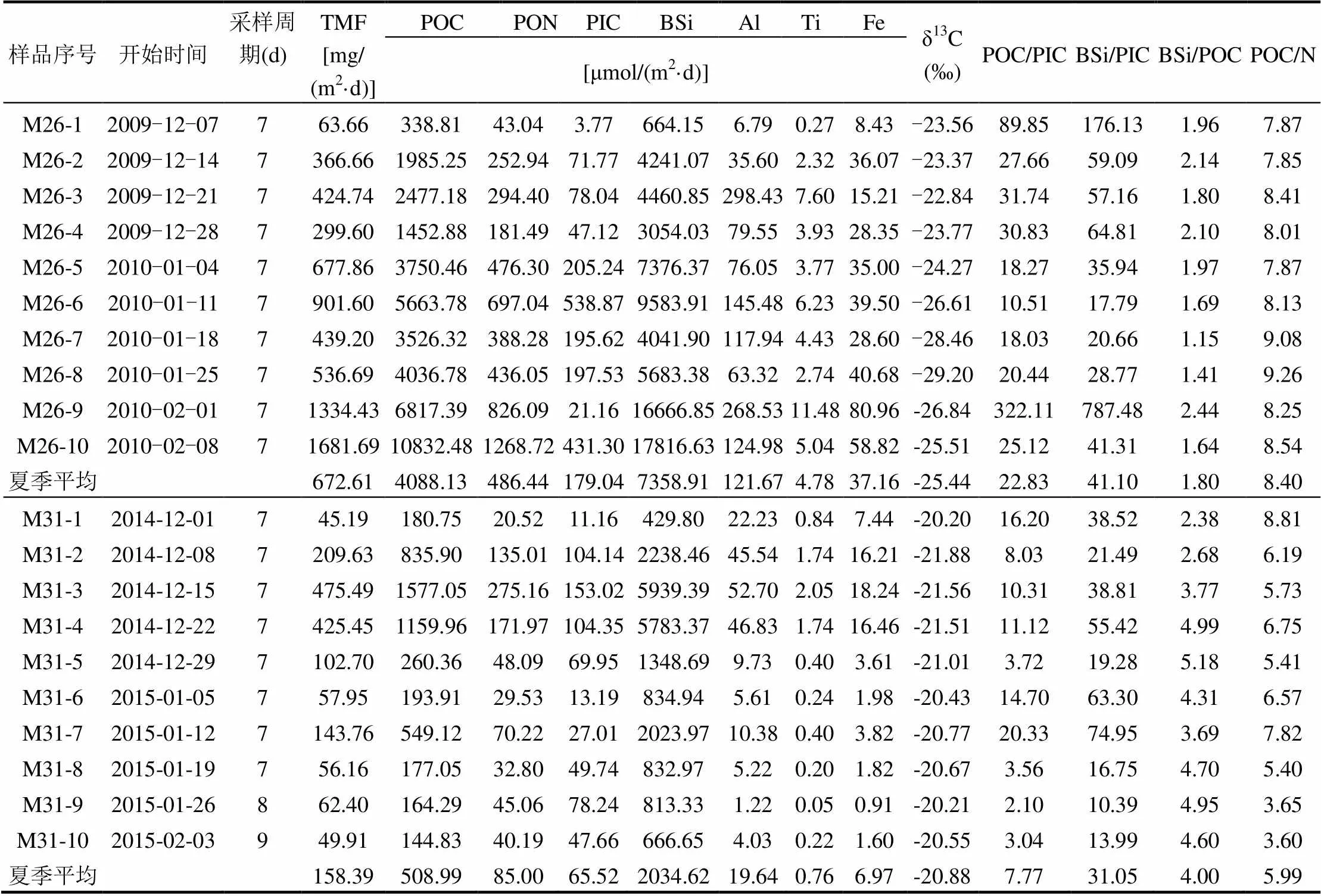
表1 南极夏季普里兹湾各生物地球化学要素沉降通量
普里兹湾冰间湖中沉降的铁被归为岩源性组分[23],但铁又是南极海洋浮游生物生长的限制元素[31],铁的溶解与清除被认为是一个复杂的物理化学过程[8].2009~2010年和2014~2015年Fe的夏季通量为37.16、6.97μmol/(m2·d)(表1).其中,2009~2010年Fe的变化趋势与Al和Ti略有差异,其第1个峰值出现早于Al、Ti,而第2个峰值则同步于Al、Ti(图6),这表明存在不同的沉降机制.该年铁的第1个峰值期间,Fe/Al(元素含量比)大于地壳中两者的比值0.68[30],可能存在一个特殊清除机制将水柱中非岩源的铁向下迁移.海冰融化后的海水中金属离子含量比极地的无冰海域高2~3个量级[32],相较于Al和Ti等非生物营养要素,海水中溶解态Fe可被浮游生物利用或吸附[33],有机质等沉降可能导致了12月中旬Fe异常高值.2014~2015年夏季Fe与Al、Ti变化趋势一致,仅在夏季初出现较小的峰值,这与海冰消融带来冰筏碎屑的输入有关.Fe和Al呈现显著正相关(2=0.999,<0.01),表明该年份Fe的来源表现为强岩源性特征,可能与该年较低的总物质通量对水体中溶解态Fe的吸附清除作用较小有关.
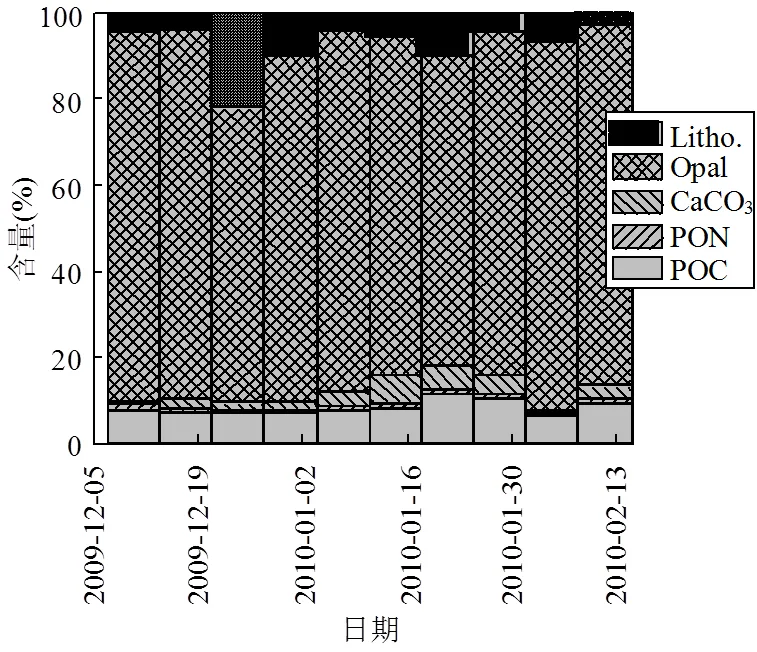
图4 2009~2010年南极夏季普里兹湾沉降通量组成时间序列分布(以5项主要成分标准化至100%)

图5 2014~2015年南极夏季普里兹湾沉降通量组成时间序列分布(以5项主要成分标准化至100%)
遥感叶绿素a(Chl a)的变化趋势与生源颗粒物(POC、BSi)显著不同.2009~2010年夏季Chl a峰值出现在12月下旬,POC和BSi的峰值相对滞后了约44d.沉降通量的滞后与沉降速率有关[2],而沉降速率则是由高密度的压舱物(蛋白石、碳酸钙和岩源物质)决定的[2].硅藻在旺发时会释放出具有絮凝作用的胞外透明聚合物(TEP),将浮游生物和压舱物形成聚集体,然后发生快速沉降形成沉降事件[34].浮游植物在12月下旬发生旺发(遥感叶绿素a出现峰值)(图3),浮游动物的摄食压力较小[35],更多的有机质在上层发生再循环,并重新供给浮游植物的光合作用.2月份岩源物质和硅藻蛋白石的增加,促使了有机质的快速沉降,并引发了该时期的沉降事件.2014~2015年夏季Chl a则出现了2次峰值,第1次出现在2015年1月上旬,第2次则出现在2月中旬,均出现在POC的峰值之后,表明浮游藻类在上层海洋未能发生快速沉降.2014~2015年夏季蛋白石和岩源颗粒物通量低于2009~2010年夏季,缺乏有效的压舱物,不利于有机质的快速沉降,导致更多的有机质在上层发生再矿化作用,致使2014~2015年夏季POC的平均通量仅为2009~2010年的23.5%.
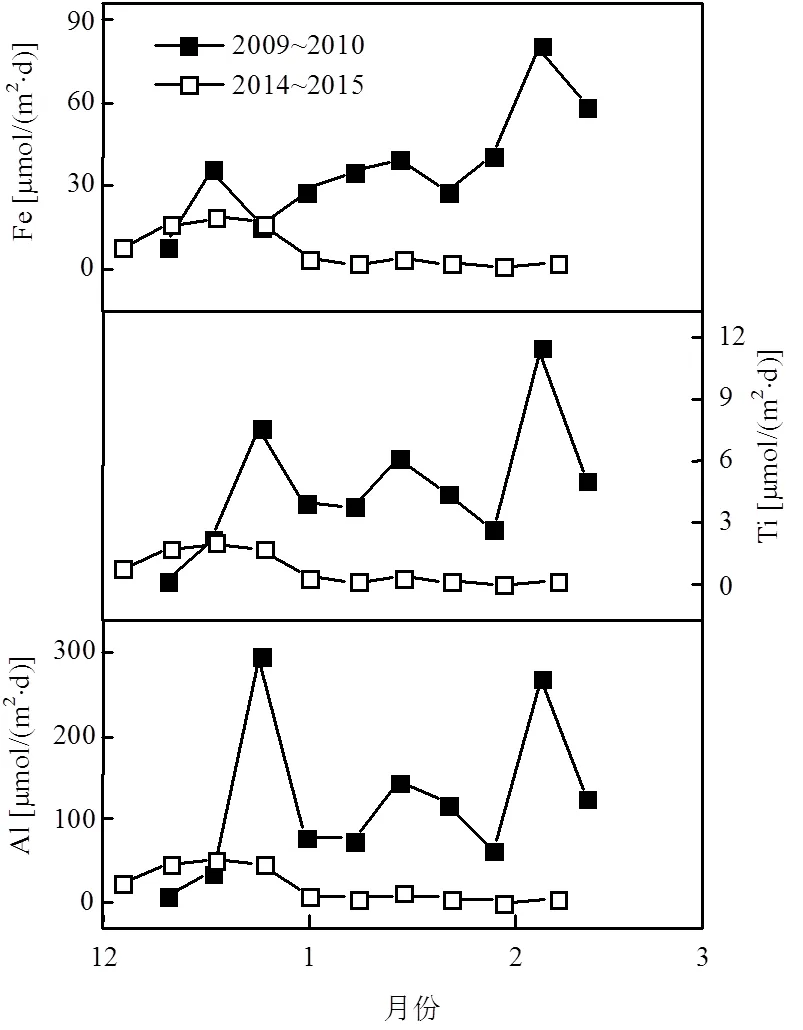
图6 南极普里兹湾岩源要素沉降通量时间序列变化

图7 沉降颗粒物中Al和Ti的相关关系
2.3 元素比和稳定同位素
2009~2010年夏季沉降通量中有机碳和氮的物质的量比值(C/N)为8.40,其变化范围为7.84~ 9.25(表1,图8),与普里兹湾湾外的C/N相近[18].南极硅藻的C/N约为6.37,略低于南极棕囊藻的7.71[36],但鉴于南极棕囊藻较低的沉降效率[37],且微生物对有机质中氨基酸和糖类等具有选择性早期降解作用[38],推测硅藻有机质在沉降过程中,细菌生产力对碳、氮降解速率的差异造成C/N偏高.2014~2015年夏季沉降通量中C/N明显低于2009~2010年夏季,其均值为5.99,并在2015年1月下旬和2月上旬的C/N出现小于4的情况,这表现出微生物C/N(3.7[39])特征,与前文的推论一致,即微生物的呼吸作用使上层大部分有机质发生了再矿化,仅有较少的有机质向下沉降.
2009~2010年和2014~2015年夏季沉降通量中生物硅和有机碳的比值(Si/C)平均分别为1.8和4.0(表1,图8),可认为普里兹湾海域为“硅海洋”(Si/C³1)[2].其变化趋势整体表现出与C/N相反的趋势.在无限制条件下,硅藻进行光合作用的Si/C为0.13[40],而在铁限制下该比值可升高2~3倍[41],但这仅适用于上层水体中硅藻生物体.沉积物捕获器样品中Si/C的比值主要由生物硅的溶解和有机质的再矿化差异造成的,而它们受到温度和微生物的共同影响[42-43].随着深度的增加,沉降颗粒物中Si/C往往呈增加趋势,直到达到沉积物中Si/C达到最大[43].较高的Si/C可能是由3个原因造成的,第一,可能是由于再悬浮沉积物的侧向输送,提高了沉积物捕获器中样品的Si/C值;第二,可能是由于上层海洋中硅藻旺发导致了海水出现了铁限制,使得硅藻的Si/C升高,进一步影响了沉降颗粒物中的Si/C;第三,异养微生物生产力升高可造成硅藻体中有机碳更大程度的降解,促使Si/C升高.考虑到两年沉积物捕获器所布放的位置及深度差异较小,沉积物再悬浮对Si/C的年际差异不予考虑. 2015年1月下旬Chl出现下降趋势可能是铁限制造成的,推测可能是Si/C升高的一个因素.此外,2014~2015年夏季压舱物(蛋白石、岩源物质)通量较低,无法引发沉降事件,导致上层海洋微生物生产力增强,可能是该年Si/C高于2009~2010年的主要因素.

图8 南极夏季普里兹湾沉降颗粒物C/N和Si/C
沉降有机质的δ13C也存在着一定的年际差异, 2009~2010年沉降有机质的δ13C平均为-25.44‰,低于2014~2015年夏季δ13C的-20.88‰(表1,图8).两个年份夏季初的δ13C较为接近,但随时间的推移,2009~2010年δ13C出现下降趋势,而2014~ 2015年则无明显变化(图8).
Lourey等[44]认为,南极夏季沉降颗粒物中偏重的δ13C可能是由于再悬浮过程将沉积物中较高的δ13C引入所致,在夏季初期由于浮游植物尚未旺发,上层有机质通量较小,因此夏季初沉降颗粒物中有机质可能受到再悬浮沉积物较大的改造,导致其偏重.而另一种可能的原因则是具有偏重的δ13C的冰藻沉降,导致夏季初的碳稳定同位素偏重.由于海冰导致冰藻的光合作用处于封闭或半封闭系统,致使碳稳定同位素产生更强的分馏,使冰藻中δ13C最高可达-8‰[45],冰藻有机质的沉降可导致夏季初沉降颗粒物中δ13C的偏高.当浮游藻类占据主导地位,上层海洋中δ13C主要受到浮游植物光合作用的强弱控制,普里兹湾1月下旬表层颗粒有机碳的δ13C最高可达-19‰[46],这是由于混合层减弱引起硅藻旺发,较强的生物初级生产引起的.考虑到有机质在沉降过程中,细菌的再矿化作用对δ13C的垂直分布是起主导作用的,一般随深度的增加,δ13C呈现整体升高的趋势[47],其差异受细菌生产力的强弱所控制.2014~ 2015年比2009~2010年夏季有机质δ13C值高约5‰,尤其是南极盛夏时期(1月和2月)差异更大.结合两年夏季的有机碳通量、C/N,推测2014~ 2015年异养细菌生产力要强于2009~2010年,从而造成该年份盛夏时期δ13C偏重,意味着该年份的有机碳沉降效率较低,前文所阐述的压舱物缺少是造成此差异的主要原因.
2.4 颗粒物通量的差异对比
与普里兹湾已有的通量结果对比发现,利用238U-234Th不平衡法所观测的输出通量高出沉积物捕获器观测结果的1~2个量级(表2),类似情况同样发生在西南极半岛海域[12].本文沉积物捕获器的通量界面深于238U-234Th不平衡法的输出界面可能是一个重要因素.根据Martin曲线[48]将有机碳沉降通量标准化至100m深度:

式中:表示深度,m;F表示深度为z的沉降碳通量;100表示100m深度的沉降碳通量;为经验系数-0.858.依据两年夏季沉降通量重新估算100m处沉降通量平均为(8.67±9.96)mmol/(m2·d),仍远小于同位素的方法所估算的通量[49-50].鉴于238U-234Th不平衡法的假定条件是234Th全部是以POC为载体迁出,忽略了其它非有机质碎屑以及浮游植物初级生产时形态和大小的改变对该模型的影响,因此,可能会对通量产生一定程度的高估.Pilskaln等[18]所报道湾外沉积物捕获器的通量根据Martin曲线折算为100m时为6.26mmol/(m2·d),与210Po-210Pb不平衡法所估算的通量值接近[51],可能是由于湾外沉降颗粒物主要是由生源物质贡献为主[18]Buesseler等[12]认为,浅层水体中沉积物捕获器的沉降通量数据存在较大的低估,这可能是由于上层水体中沉积物捕获器周围较强的侧向海流造成的捕获效率低下[52].但本文所布放的沉积物捕获器位于水体相对稳定的冰间湖,通量界面分别位于480m和390m,浅层水体变化对沉降颗粒物的影响因素可以忽略.尽管如此,两者观测方法造成的差异仍然需要更多的观测和数据去验证,尤其是年际差异、季节差异、颗粒物组成等诸多因素的影响.

表2 依据不同的方法所估算的普里兹湾沉降碳通量
注:*取12月30日至次年3月10日进行夏季通量估算.
2.5 与南极其他海域对比
表3给出了南极各边缘海基于沉积物捕获器所观测到的有机碳沉积通量.罗斯海是南极高生产力边缘海之一,其冰间湖的初级生产仅次于阿蒙森冰间湖[19],与之相比,普里兹湾的夏季沉降通量要远高于罗斯海夏季沉降通量,基于遥感估算的1月份普里兹湾冰间湖多年平均初级生产力为(100±29)mmol/(m2·d),略低于罗斯海冰间湖的(122±23)mmol/(m2·d)[19],而沉降通量的显著差异表明普里兹湾冰间湖具有更高的沉降效率.Collier等[8]在1996~1998年期间观测到了罗斯海最高的沉降通量,POC通量的主要峰值出现在秋季(4~6月),均在海冰覆盖之后,并且上层(~200m)的POC通量仅为下层(~480m)的1/3,因此沉积物再悬浮可能是其高沉降通量的原因[8].表层沉积物中Si/Al通常低于水柱中颗粒物,所以低Si/Al被认为是沉积物再悬浮的一个指示[8].普里兹湾POC和BSi通量出现峰值时Si/Al也较高,因此沉积物再悬浮对本研究中POC的峰值贡献较小.尽管罗斯海的生产力更高,但南极棕囊藻是罗斯海的春季浮游植物旺发的优势种[36,53],其具有非常低的沉降效率[37],对沉降通量贡献较低[4].利用遥感初级生产力的数据[19],通过Martin曲线估算夏季普里兹湾的100m深度沉降通量约平均为净初级生产力的8.67%,,与南极半岛海域170m观测的多年结果(~5%)相近[13].同南极半岛海域相同,普里兹湾冰间湖主要以硅藻为优势种[25-27],硅藻聚集体的快速沉降是高沉降效率的主要原因.
南极半岛海域通常被认为是具有高效生物泵效率的海域[12],但Weston等[16]通过沉积物捕获器和营养盐亏损的2种方法对南极半岛海域的Marguerite湾进行了有机碳通量和效率估算,发现200m深度输出效率仅为1%,认为该区域具有高循环低输出的特点.Ducklow等[4]认为,该海湾环流结构复杂,沉降通量可能是由于平流输送导致偏低,表现出低输出的特征,因此无法代表半岛海域的生物泵效率.
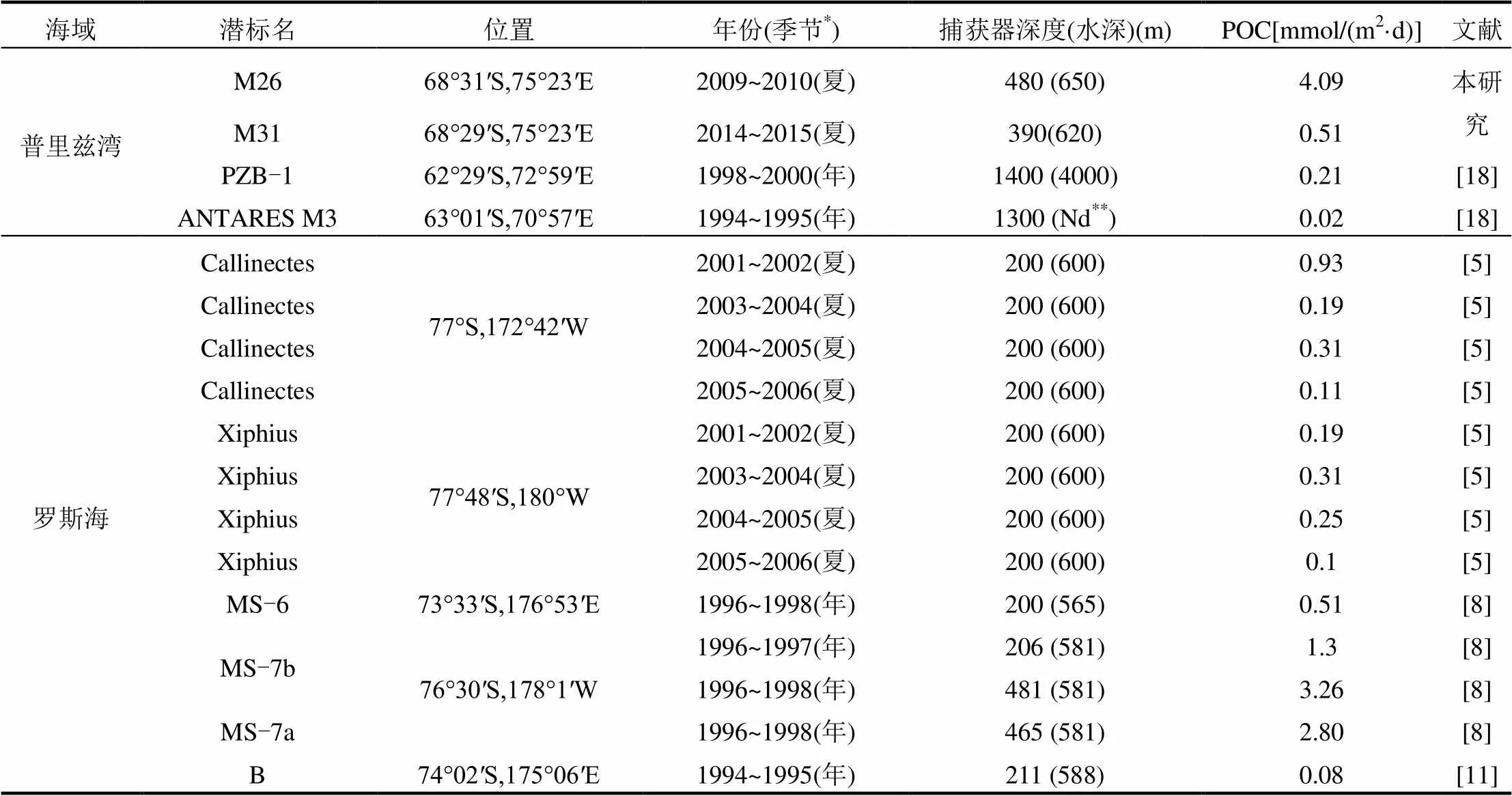
表3 南极各边缘海有机碳沉降通量比较

续表3
注:*表示通量数据所覆盖的时间范围:“夏”代表夏季平均通量;“年”代表年平均通量;**表示数据未能获取;#表示1993~2006年(除2004年外)平均通量和标准偏差.
阿蒙森冰间湖是南极初级生产力最高的冰间湖,其年际平均生产力达(160.7±36.9) g C/ (m2·a)[19],但2011~2012年深度为350m处的平均通量仅为0.86μmol/(m2·d)[13].以往的研究认为极地海域具有更高的生物泵效率[1,12],阿蒙森冰间湖春季的优势藻种是南极棕囊藻,这导致了生物泵效率要低于硅藻为优势种的海域[4],与罗斯海较为相似[5].某种程度上,浮游植物群落结构的差异是影响南极海域的生物泵效率的主要因素,硅藻和南极棕囊藻则分别控制各自优势海域的输出效率.在气候变化的大背景下,南极各个边缘海将呈现出不同的变化趋势,浮游植物群落结构可能将出现明显差异[54],因此,这就需要更高时间分辨率和更综合的观测来深入理解生物泵的效率机制[4].
3 结论
3.1 南极普里兹湾2009~2010年和2014~2015年颗粒有机碳夏季平均通量分别为4088.13、508.99μmol/ (m2·d),生物硅夏季平均通量分别为7358.91、2034.63μmol/(m2·d).生物硅占夏季颗粒物通量均超过70%,表明硅藻是普里兹湾夏季的优势种和沉降通量的主要贡献者.
3.2 缺少压舱物易导致上层有机质无法快速沉降,令异养微生物对上层水柱中有机质降解程度更高,造成2014~2015年夏季的有机碳沉降通量仅为2009~2010年12.5%.
3.3 沉积物捕获器所观测的结果要略低于之前普里兹湾238U-234Th不平衡法的观测结果,但考虑到年际差异等因素,仍具有一定的可对比性.夏季普里兹湾具有非常高的沉降通量和沉降效率, 通过Martin曲线估算夏季普里兹湾的100m深度沉降通量平均为净初级生产力的8.67%,这种高生物泵运转效率归因于硅藻在水柱中的优势地位.
[1] Ducklow H, Steinberg D, Buesseler K. Upper Ocean Carbon Export and the Biological Pump [J]. Oceanography, 2001,14(4): 50-58.
[2] Honjo S, Manganini S J, Krishfield R A, et al. Particulate organic carbon fluxes to the ocean interior and factors controlling the biological pump: A synthesis of global sediment trap programs since 1983 [J]. Progress in Oceanography, 2008,76(3):217-285.
[3] Takahashi T, Sutherland S C, Wanninkhof R, et al. Climatological mean and decadal change in surface oceanCO2, and net sea–air CO2flux over the global oceans [J]. Deep-Sea Research Part II: Topical Studies in Oceanography, 2009,56(8-10):554-577.
[4] Ducklow H W, Wilson S E, Post A F, et al. Particle flux on the continental shelf in the Amundsen Sea Polynya and Western Antarctic Peninsula [J]. Elementa-Science of the Anthropocene, 2015,3:46.
[5] Smith W O, Shields A R, Dreyer J C, et al. Interannual variability in vertical export in the Ross Sea: Magnitude, composition, and environmental correlates [J]. Deep-Sea Research Part I: Oceanographic Research Papers, 2011,58(2):147-159.
[6] Anderson R F, Carr M E. Paleoclimate. Uncorking the Southern Ocean's vintage CO2[J]. Science, 2010,328(5982):1117-1118.
[7] Ducklow H W, Baker K, Martinson D G, et al. Marine pelagic ecosystems: the west Antarctic Peninsula [J]. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences, 2007,362(1477):67-94.
[8] Collier R, Dymond J, Honjo S, et al. The vertical flux of biogenic and lithogenic material in the Ross Sea: moored sediment trap observations 1996–1998 [J]. Deep-Sea Research Part II: Topical Studies in Oceanography, 2000,47(15/16):3491-3520.
[9] Honjo S. Particle export and the biological pump in the Southern Ocean [J]. Antarctic Science, 2004,16(4):501-516.
[10] Honjo S, Francois R, Manganini S, et al. Particle fluxes to the interior of the Southern Ocean in the Western Pacific sector along 170°W [J]. Deep-Sea Research Part II: Topical Studies in Oceanography, 2000,47(15/16):3521-3548.
[11] Langone L, Frignani M, Ravaioli M, et al. Particle fluxes and biogeochemical processes in an area influenced by seasonal retreat of the ice margin (northwestern Ross Sea, Antarctica) [J]. Journal of Marine Systems, 2000,27(1-3):221-234.
[12] Buesseler K O, McDonnell A M P, Schofield O M E, et al. High particle export over the continental shelf of the west Antarctic Peninsula [J]. Geophysical Research Letters, 2010,37(22):208- 217.
[13] Ducklow H W, Erickson M, Kelly J, et al. Particle export from the upper ocean over the continental shelf of the west Antarctic Peninsula: A long-term record, 1992~2007 [J]. Deep-Sea Research Part II: Topical Studies in Oceanography, 2008,55(18): 2118-2131.
[14] Kim D Y, Kim D Y, Kim Y J, et al. Downward fluxes of biogenic material in Bransfield Strait, Antarctica [J]. Antarctic Science, 2003,16(3):227-237.
[15] Wefer G, Fischer G. Annual primary production and export flux in the Southern Ocean from sediment trap data [J]. Marine Chemistry, 1991,35(1):597-613.
[16] Weston K, Jickells T D, Carson D S, et al. Primary production export flux in Marguerite Bay (Antarctic Peninsula): Linking upper water-column production to sediment trap flux [J]. Deep-Sea Research Part I: Oceanographic Research Papers, 2013, 75(5):52-66.
[17] Kim M, Hwang J, Kim H J, et al. Sinking particle flux in the sea ice zone of the Amundsen Shelf, Antarctica [J]. Deep-Sea Research Part I: Oceanographic Research Papers, 2015,101: 110-117.
[18] Pilskaln C H, Manganini S J, Trull T W, et al. Geochemical particle fluxes in the Southern Indian Ocean seasonal ice zone: Prydz Bay region, East Antarctica [J]. Deep-Sea Research Part I: Oceanographic Research Papers, 2004,51(2):307-332.
[19] Arrigo K R. Phytoplankton dynamics within 37 Antarctic coastal polynya systems [J]. Journal of Geophysical Research, 2003, 108(C8):3271.
[20] Hedges J I, Stern J H. Carbon and nitrogen determinations of carbonate-containing solids [J]. Limnology & Oceanography, 1984,29(3):657-663.
[21] Mortlock R A, Froelich P N. A simple method for the rapid determination of biogenic opal in pelagic marine sediments [J]. Deep-Sea Research Part A: Oceanographic Research Papers, 1989,36(9):1415-1426.
[22] US EPA. Method3052. Microwave assisted acid digestion of siliceous and or fanically based matrices [EB/OL]. http://www. epa.gov/osw/hazard/testmethods/sw846/pdfs/3052.pdf.1996-12.
[23] Sun W P, Han Z B, Hu C Y, et al. Source composition and seasonal variation of particulate trace element fluxes in Prydz Bay, East Antarctica [J]. Chemosphere, 2016,147:318-27.
[24] Wong A P S, Bindoff N L, Forbes A. Ocean-Ice shelf interaction and possible bottom water formation in Prydz Bay, Antarctica [C]. Washington: American Geophysical Union, 1998:173-187.
[25] 刘诚刚,宁修仁,孙 军,等.2002年夏季南极普里兹湾及其邻近海域浮游植物现存量,初级生产力粒级结构和新生产力研究 [J]. 海洋学报, 2004,26(6):107-117.
[26] 孙 军,刘东艳,宁修仁,等.2001/2002年夏季南极普里兹湾及其邻近海域的浮游植物 [J]. 海洋与湖沼, 2003,34(5):519-532.
[27] 朱根海,宁修仁.1991/1992年夏季南极普里兹湾邻近海域网采浮游植物的聚类分析 [J]. 南极研究, 1995,7(2):38-44.
[28] Taylor F, McMinn A, Franklin D. Distribution of diatoms in surface sediments of Prydz Bay, Antarctica [J]. Marine Micropaleontology, 1997,32(3):209-229.
[29] Wilks J V, Rigual-Hernandez A S, Trull T W, et al. Biogeochemical flux and phytoplankton succession: A year-long sediment trap record in the Australian sector of the Subantarctic Zone [J]. Deep-Sea Research Part I: Oceanographic Research Papers, 2017,121:143-159.
[30] Taylor S R. Abundance of elements in the crust: A new table [J]. Geochimica and Cosmochimica Acta, 1964,28:1273-1285.
[31] Martin J H, Fitzwater S E, Gordon R M. Iron deficiency limits phytoplankton growth in Antarctic waters [J]. Global Biogeochemical Cycles, 1990,4(1):5-12.
[32] Lannuzel D, van der Merwe P C, Townsend A T, et al. Size fractionation of iron, manganese and aluminium in Antarctic fast ice reveals a lithogenic origin and low iron solubility [J]. Marine Chemistry, 2014,161:47-56.
[33] Riedel T, Zak D, Biester H, et al. Iron traps terrestrially derived dissolved organic matter at redox interfaces [J]. Proceedings of the National Academy of Sciences, 2013,110(25):10101-10105.
[34] Passow U, Alldredge A L. Aggregation of a diatom bloom in a mesocosm: The role of transparent exopolymer particles (TEP) [J]. Deep-Sea Research Part II: Topical Studies in Oceanography, 1995,42(1):99-109.
[35] Yang G, Li C L, Sun S, et al. Feeding of dominant zooplankton in Prydz Bay, Antarctica, during austral spring/summer: food availability and species responses [J]. Polar Biology, 2013,36(11): 1701-1707.
[36] Arrigo K R, Robinson D H, Worthen D L, et al. Phytoplankton community structure and the drawdown of nutrients and CO2in the southern ocean [J]. Science, 1999,283(5400):365-367.
[37] Wolf C, Iversen M, Klaas C, et al. Limited sinking of Phaeocystis during a 12 days sediment trap study [J]. Molecular Ecology, 2016,25(14):3428-3435.
[38] Meyers P A. Organic geochemical proxies of paleoceanographic, paleolimnologic, and paleoclimatic processes [J]. Organic Geochemistry, 1997,27(5/6):213-250.
[39] Lee S, Fuhrman J. Relationships between biovolume and biomass of naturally derived marine bacterioplankton. [J]. Applied and environmental microbiology, 1987,53(6):1298–1303.
[40] Brzezinski M A. The Si: C: N ratio of marine diatoms: Interspecific variability and the effect of some environmental variables [J]. Journal of Phycology, 1985,21(3):347-357.
[41] Takeda S. Influence of iron availability on nutrient consumption ratio of diatoms in oceanic waters [J]. Nature, 1998,393(6687): 774-777.
[42] Bidle K D, Manganelli M, Azam F. Regulation of oceanic silicon and carbon preservation by temperature control on bacteria [J]. Science, 2002,298(5600):1980-1984.
[43] Nelson D M, Anderson R F, Barber R T, et al. Vertical budgets for organic carbon and biogenic silica in the Pacific sector of the Southern Ocean, 1996~1998 [J]. Deep-Sea Research Part II: Topical Studies in Oceanography, 2002,49(9/10):1645-1674.
[44] Lourey M J, Trull T W, Tilbrook B. Sensitivity of δ13C of Southern Ocean suspended and sinking organic matter to temperature, nutrient utilization, and atmospheric CO2[J]. Deep-Sea Research Part I: Oceanographic Research Papers, 2004, 51(2):281-305.
[45] Thomas D N, Dieckmann G S. Sea ice: An introduction to its physics, chemistry, biology and geology [M]. Berlin: John Wiley & Sons, 2003.
[46] Zhang R, Zheng M F, Chen M, et al. An isotopic perspective on the correlation of surface ocean carbon dynamics and sea ice melting in Prydz Bay (Antarctica) during austral summer [J]. Deep-Sea Research Part I: Oceanographic Research Papers, 2014, 83:24-33.
[47] 任春燕,陈 敏,高众勇,等.南极普里兹湾及其邻近海域悬浮颗粒有机物的碳同位素组成及其影响因素 [J]. 海洋学报, 2015, 37(12):74-84.
[48] Martin J H, Knauer G A, Karl D M, et al. VERTEX: carbon cycling in the northeast Pacific [J]. Deep-Sea Research Part A: Oceanographic Research Papers, 1987,34:267-285.
[49] Ma H, Zeng Z, He J, et al.234Th-derived particulate organic carbon export in the Prydz Bay, Antarctica [J]. Journal of Radioanalytical and Nuclear Chemistry, 2013,299(1):621-630.
[50] He J H, Ma H, Chen L Q, et al. The investigation on particulate organic carbon fluxes with disequilibria between thorium-234 and uranium-238 in the Prydz Bay, the Southern Ocean [J]. Acta Oceanologica Sinica, 2008,27(2):21-29.
[51] 杨伟锋,黄奕普,陈 敏,等.基于210Po/210Pb不平衡的南大洋与南海颗粒有机物的输出与再矿化 [J]. 科学通报, 2009,(11): 1579-1584.
[52] Buesseler K O, Antia A N, Chen M, et al. An assessment of the use of sediment traps for estimating upper ocean particle fluxes [J]. Journal of Marine Research, 2007,65(3):345-416.
[53] Smith Jr W O, Ainley D G, Arrigo K R, et al. The oceanography and ecology of the Ross Sea [J]. Annual Review of Marine Science, 2014,6(1):469-487.
[54] Deppeler S L, Davidson A T. Southern Ocean phytoplankton in a changing climate [J]. Frontiers in Marine Science, 2017,4:40.
致谢:作者在此感谢雪龙船在潜标的布放和回收过程中给予的帮助,感谢陈倩娜在稳定同位素分析中给予的帮助.
Sinking particle fluxes during austral summer in the Prydz Bay polynya, Antarctica.
HAN Zheng-bing1,2,3, SUN Wei-ping2,3, FAN Gao-jing1,2,3, HU Chuan-yu2,3, PAN Jian-ming2,3*, ZHAO Jun2,3, ZHANG Hai-feng2,3, LI Dong2,3, ZHANG Hai-sheng1,2,3
(1.College of Marine Science and Technology, China University of Geosciences, Wuhan 430074, China;2.Key Laboratory of Marine Ecosystem and Biogeochemistry, State Oceanic Administration, Hangzhou 310012, China;3.Second Institute of Oceanography, State Oceanic Administration, Hangzhou 310012, China)., 2018,38(5):1923~1934
To understand the biological pump of the Antarctic Ocean, two sets of time-series sediment trap was deployed and recovered in the polynya of the Prydz Bay during the 26thand 31stChinese Antarctic scientific expeditions in the austral summer of 2009~2010 and 2014~2015. The particle fluxes were analyzed. The results showed that the summer fluxes of particulate organic carbon (POC) were 4088.13 and 508.99μmol/(m2·d) for the year of 2009~2010 and 2014~2015, respectively, while biogenic silica (BSi) were 7358.91 and 2034.63 μmol/(m2·d), respectively. The biogenic opal contributed more than 70% of total mass flux in two years, which indicated diatoms should be the dominated species in the column and the main carriers of sinking POC during austral summer. The POC flux in 2014~2015 was only 12.5% of that in 2009~2010. It was mainly due to the lack of ballast matters that prevented the rapid settling of organic matter in the upper layer, resulting in more degrading organic matter in the upper water column caused by heterotrophic microorganisms. Compared with other Antarctic marginal seas where the blooms were not dominated by diatoms, the Prydz Bay polynya is characterized of higher sinking fluxes and efficiency. Estimated by the Martin curve, the 100m POC flux in summer in Prydz Bay averages 8.67% of the net primary productivity. It mainly owed to the diatom-dominated plankton community in the Prydz Bay polynya, therefore the plankton community in polar oceans should attract more attention when predicting the variation of biological pump.
particle fluxes;polynya;Prydz Bay;biological pump
X55
A
1000-6923(2018)05-1923-12
2018-02-07
国家自然科学青年基金(41406219,41576186,41506223,41376193);中央级公益性科研院所基本科研业务费专项资金(JT1405);南北极环境综合考察与评估极地专项(CHINARE01-04,CHINARE04-01,CHINARE04-04)
* 责任作者, 研究员, jmpan@sio.org.cn
韩正兵(1986-),男,江苏连云港人,助理研究员,在读博士生,主要从事极地海洋生物地球化学研究.发表论文20余篇.
