Morphometric study con firms the presence of only Vatica mangachapoi on Hainan Island,China
2018-05-19JunjieGuoShuaibinShangJieZeng
Junjie Guo•Shuaibin Shang•Jie Zeng
Introduction
Vatica mangachapoiis a dipterocarp species famous for its timber and non-wood byproducts.Its timber is important commercially in international tropical timber markets(Appanah and Turnbull 1998)and its leaves and stems contain a number of secondary metabolites that are used in perfumes and medicines(Qin et al.2011).This species is widely distributed in Southeast Asia(Blanco 1837),and the populations of the genusVaticaLinn.(Dipterocarpaceae)on Hainan Island of China were first identi fied asV.mangachapoi(Qu 1956).Later,Wang(1985)treated the Hainan populations as a new species,V.hainanensisHT Chang et LC Wang,and subdivided it into two varieties,V.hainanensisvar.parvifoliaHT Chang apud LC Wang andV.hainanensisvar.glandipetalaLC Wang.Subsequently Yang et al.(2002)considered all Hainan populations as a subspecies ofV.mangachapoi(V.mangachapoispp.hainanensis).However,all these treatments were not accepted by Chen(2005)inFlora of Guangdongor by Li et al.(2007)inFlora of China,and onlyV.mangachapoiwas considered to be native on Hainan Island.Nevertheless,a new variety,V.mangachapoispp.hainanensisvar.wanningensisFu and Yang(2008),was described for populations growing on coastal sandy soils.These taxonomic studies did not quantify morphological variations at population level.The objectives of the present study were to quantify the variations of morphological characters within and among the populations of the taxa mentioned above and to determine their taxonomic validity.
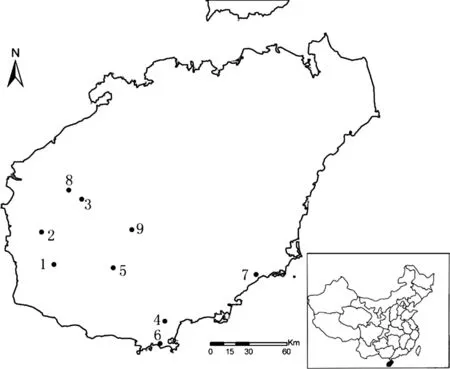
Fig.1 Localities of nine natural populations of Vatica spp.on Hainan Island,China
Materials and methods
Sample collection
Nine populations of the genusVaticawere sampled on Hainan Island between 2011 and 2013(Fig.1;Table 1).Some were populations from which the type specimens of the new species,subspecies and varieties described above were collected.Six to thirty-four mature trees were randomly sampled in each population with the distances greater than 100 m between sampled trees.In total 133 individuals were sampled.For all sampled trees,morphological traits of leaf and fruit were measured(Table 2).Voucher specimens were deposited in Research Institute of Tropical Forestry(RITF),Chinese Academy of Forestry.Eighty-three herbarium specimens of the taxa mentioned above in South China Agricultural University Herbarium(CANT),Sun Yet-sen University Herbarium(SYS)and Chinese Academy of Forestry Herbarium(CAF)were also checked,and 11 specimens with fully-developed leaves and fruits were measured.
Morphological measurement
Species ofVaticaare generally identi fied by their fruit sepal size,lamina size,lateral vein number,base form,and petiole length(Li et al.2007).Twelve morphological characters of leaf and fruit were measured for each sampled tree and for specimens deposited in herbaria(Table 2).Thirty fruits and 15 completely developed leaves were measured for each sampled tree.On herbarium specimens,all fruits and fully developed leaves were measured.Themeasurement accuracy was about 0.01 mm for length and width and 0.1°for angle of lamina base(Table 2).Ratios of the longest fruit sepal length to width,the shortest fruit sepal length to width,fruit length to diameter(width),and lamina length to width were calculated to roughly depict shapes of these organs(Frampton and Ward 1990).

Table 1 Nine natural populations of Vatica spp.on Hainan Island,China
Statistical analysis
Coefficients of variation(CV)for each trait were calculated within each population and among all nine populations to assess variability.The CV among populations was calculated based on the mean value of each population.Box and whisker plots were drawn for fruit and leaf measurements of each population to assess whether morphological variation was continuous.Software NTSYS version 2.1 was applied to calculate Nei’s(1972)distances between individuals and generate a dendrogram of all sampled individuals and specimens by the neighbor joining method.FigTree version 1.3.1 was used to draw annular dendrogram.
Results
Of the 16 leaf and fruit traits measured,the angle of the lamina base(ALB)and height of maximum lamina width point(HMLW)showed in general the highest coefficients of variation(CV)within populations,and the ratio of fruit height to width(RFHW)the lowest(Table 3).Lower variances within populations were observed for fruit traits than for leaf traits.CV values were>20%for lamina length(LL),lamina width(LW)and HMLW in P5 and ALB in P9,while they were<6%for RFHW in all populations.The variances among populations for fruit traits were alsolower than those for leaf traits:the CV of RFHW was<2%.As a whole,four ratio traits were less variable than size traits within as well as among populations.
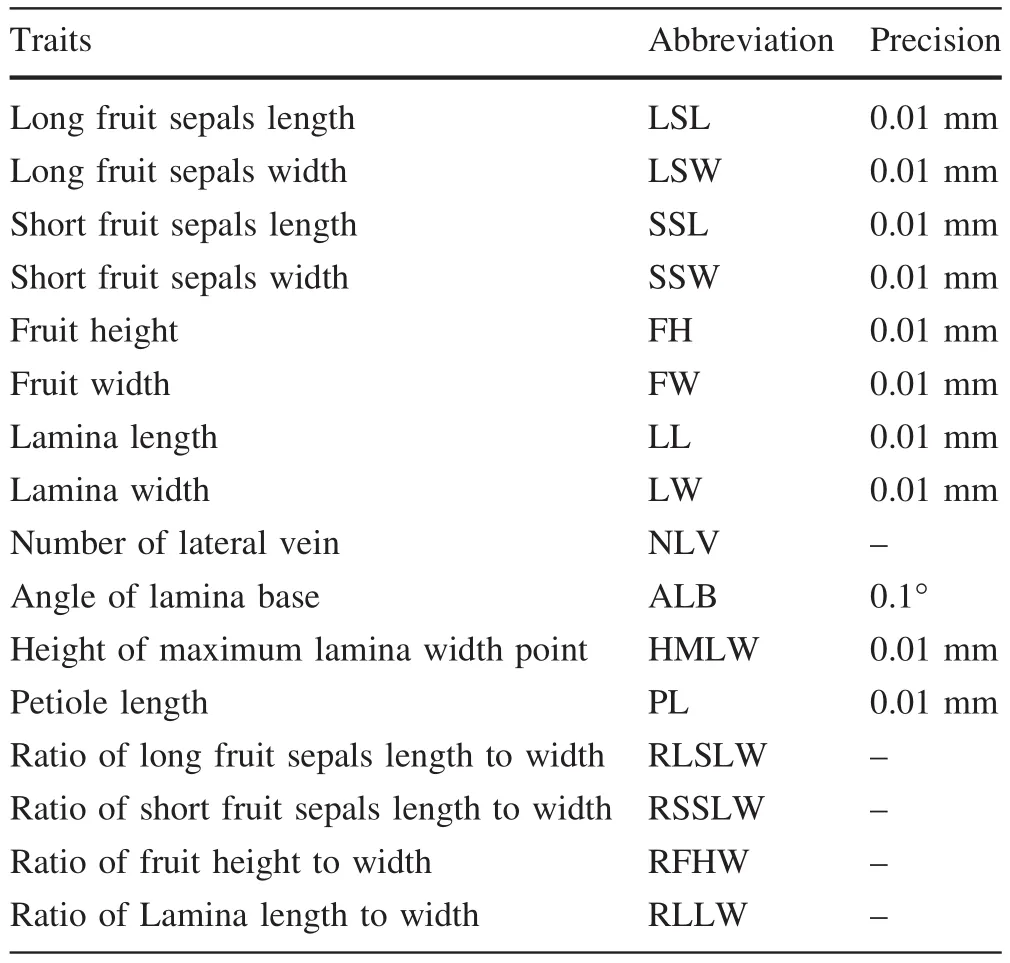
Table 2 Traits measured in the present study
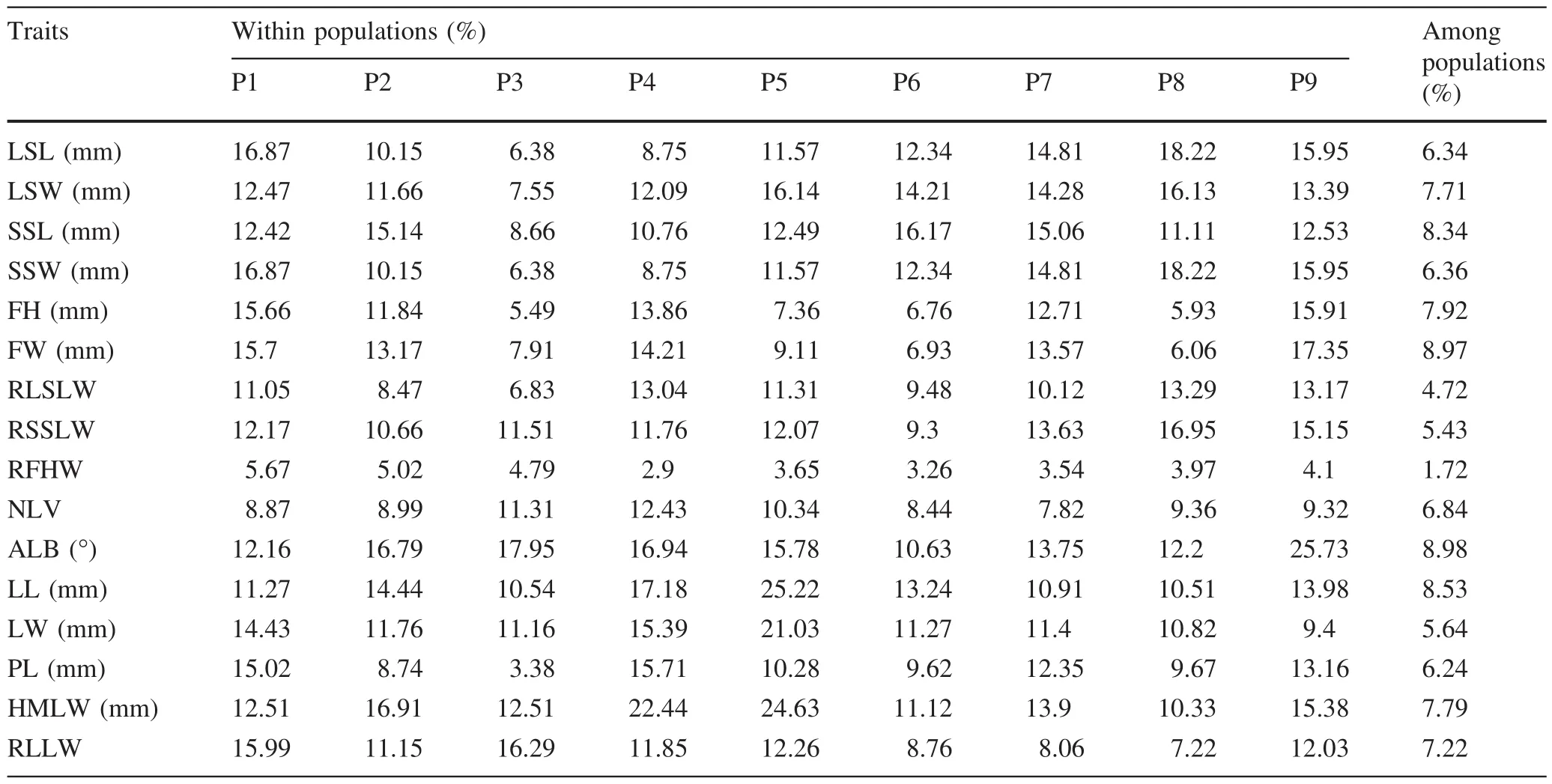
Table 3 Coefficients of variation for leaf and fruit traits in nine natural populations of Vatica spp.on Hainan Island,China
For all samples of the nine populations,the variation ranges of LL,LW and HMLW were the greatest of the 16 traits and their maxima were above five times the minimum(Table 4).Meanwhile,RFHW varied least with its ratio of maximum to minimum being approximately 1.7.The number of lateral veins(NLV)was the least variable of 12 directly-measured traits.All nine populations overlapped considerably for each of 16 morphological traits.Variation of each morphological trait was continuous among individuals and populations(Fig.2).
Cluster analysis shows the relationships between individuals among and within populations.Individuals of any population were not clearly distinguished from those of other populations and specimens of each variety did not cluster(Fig.3;Table 5).From this we conclude that the published subspeci fic taxa possessed no morphological characters that distinguish them fromV.mangachapoi.

Fig.2 Box and whisker plots showing the range of variation in fruit and leaf traits in the nine populations of Vatica spp.on Hainan Island,China.The edges of the box are 25%and 75%;the vertical lines are drawn from the box to the most extreme point within 1.5 interquartile ranges;and the transverse lines in box are the medians.See Table 1 for abbreviations of traits
Discussion
We quanti fied differences among individuals and populations for 16 morphological traits.Our results demonstrate that there are morphological variations among populations of the species.These might be caused by genetic variability,micro-scale adaptation or environmentally-induced differences(Schluter 2001;Alan 2002;Shi et al.2009).We also documented by field survey thatVatica spp.showed abundant variation of leaf and fruit morphology at a range of sites from coastal sand to montane rain forest on Hainan Island.
To determine whether a species is a recognizable taxon,it is necessary to quantify its patterns of morphological variation and compare those to patterns displayed by its closest relatives(Hsu 1998).Nooteboom(1992)concluded that it is necessary to conduct field surveys and to study enough specimens ideally representing the entire geographic range of the subject taxon.Chang and Wang(Wang 1985)identi fied populations ofVaticaon Hainan Island as a new endemic species,V.hainanensis,rather than part of the variation within the widespreadV.mangachapoi.They considered that the species ofVaticaon Hainan Island was characterized as follows:NLE,10–13;PL,8–15 mm;LSL,40–55 mm;LSW,12–20 mm;SSL,12–20 mm;SSW,5–8 mm(Wang 1985).These measurements substantially differed from those ofV.mangachapoi(NLE,7–10;PL,15–20 mm;LSL,30–40 mm;LSW,10–15 mm;SSL,8 mm;SSW,4 mm;Blanco 1837).In the present study on morphological variations of leaf and fruit traits of 133 individuals from nine populations,we found that the variation ranges of these traits were much wider(NLE,8–21;PL,5–21 mm;LSL,22–78 mm;LSW,6–22 mm;SSL,10–32 mm;SSW,3–9 mm).These measurements indicated that only SSL of the species on Hainan Island was obviously greater than that ofV.mangachapoi,the ranges of NLE,PL,LSL,LSW and SSW found in the present study overlapped those described in the literature with both taxa identi fied.Since the study at population level yields a more accurate estimate of the variance within a taxon and is much more reliable than conclusions based on a small number of samples in herbaria(Nooteboom 1992;Hsu 1998),the two taxa could not be separated from each other.Our results suggest that all native populations ofVaticaon Hainan Island are referable toV.mangachapoiwithout occurrence of subspecies because they are not distinguishable from the species described inFlora de Filipinas(Blanco 1837).
Our study also showed that each morphological trait showed continuous variation with a wide range on Hainan Island(Table 4;Fig.2).The range of variation overlapped the reported measurements for var.parvifolia(LL:40–60 mm;LW:15–20 mm;LSL:30–40 mm;and LSW:8–10 mm)and var.glandipetala(LSL:22–50 mm;LSW:5–12 mm;and NLV:6–9)published by Wang(1985)and Yang et al.(2002),and var.wanningensis(NLV:8–9;and LSL:45–52 mm)reported by Fu and Yang(2008)for each natural population.It is notable that the type specimen of varietyglandipetalawas collected from trees in Guangdong Academy of Forestry which were introduced from Bawangling,Hainan Island.Additionally,the presence of glands on the inner side of petal bases(Wang 1985)and indumentum differences on small branches(Fu and Yang 2008)are hardly suf ficient differences for identifying varietyglandipetalaandwanningensis,respectively.Cluster analysis revealed that individuals in any population did not cluster,and specimens of a described taxon were assigned to the other taxa.This made it impossible to assign a given individual to any population based on these morphological traits.In other words,the varieties were poorly separated by cluster analysis based on Nei’s(1972)distances.The three varieties,parvifolia,glandipetala,andwanningensis,could thus not be distinguished from one another or from the type specimens ofV.mangachapoi.
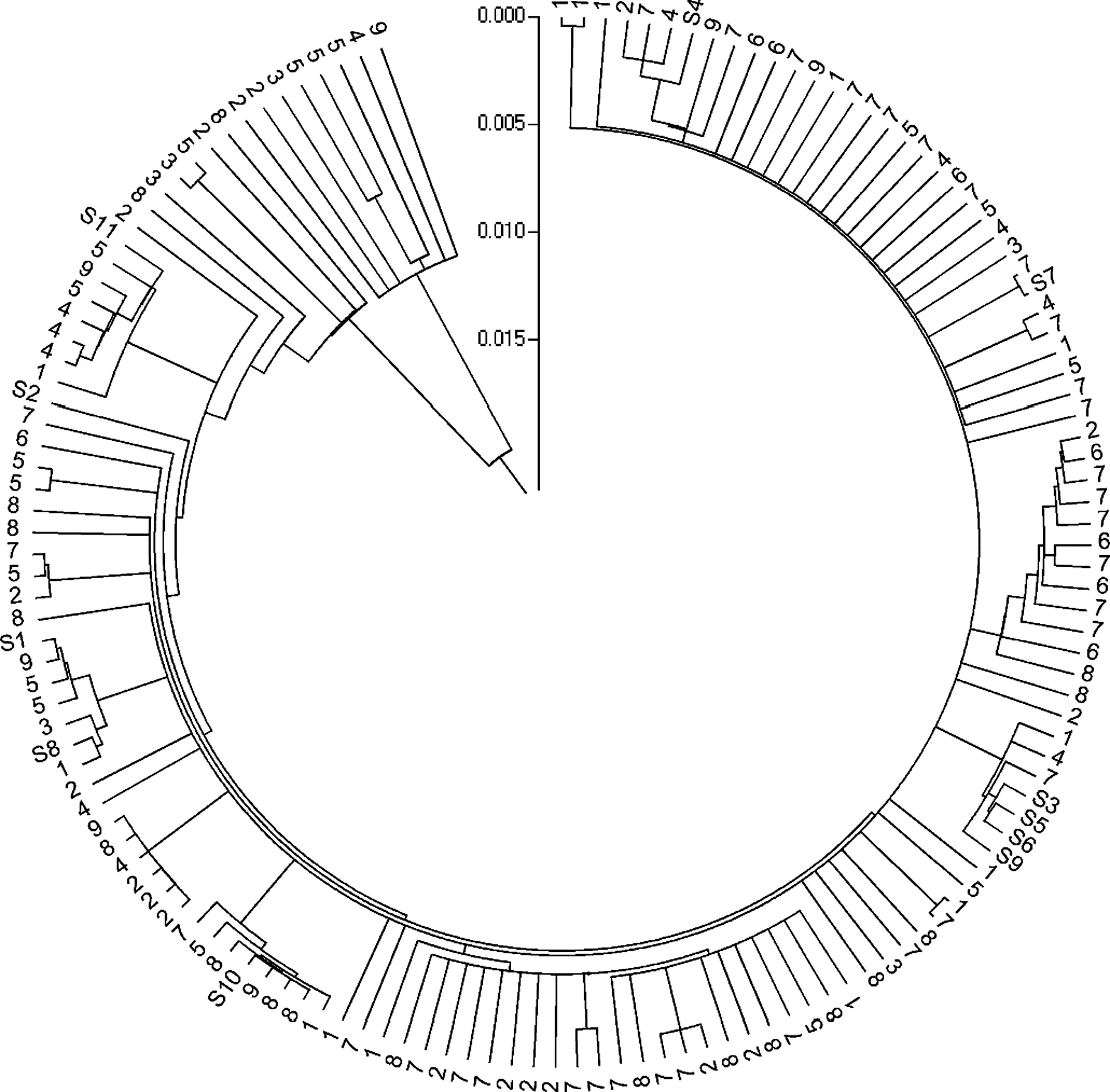
Fig.3 Dendrogram at individual level generated from neighbor joining analysis based on Nei’s(1972)distances for Vatica spp.on Hainan Island,China.Note 1–9,codes of the populations listed in Fig.1,the same number means individuals from the same population;S1–S11,codes of specimens of taxa published;and particularly,S6–S11 were collected from trees in Guangdong Academy of Forestry,Guangzhou City(GAF)which were introduced from Bawangling,Changjiang County in 1960.See Table 5 in detail

Table 5 Detailed description of Fig.3
Conclusion
The present study showed that all populations ofVaticaon Hainan Island,China should be referred toVatica mangachapoiwith no subspeci fic categories being recognized.This conclusion supports the taxonomic treatment of the genusVaticafrom Hainan Island as reported inFlora of GuangdongandFlora of China.
Taxonomic treatment
Vatica mangachapoiBlanco,Flora de Filipinas,ed.1,401,1837.
=Vatica hainanensisHT Chang et LC Wang,Acta Scientiarum Naturalium Universitatis Sunyatseni,24(3):94,1985;Vatica mangachapoiBlanco ssp.hainanensis(HT Chang et LC Wang)YK Yang et JK Wu,Chinese Wild Plant Resources 21(5):1,2002.
=Vatica hainanensisHT Chang et LC Wang var.parvifoliaH.T.Chang,Acta Scientiarum Naturalium Universitatis Sunyatseni,24(3):96,1985;Vatica mangachapoiBlanco var.parvifolia(HT Chang)YK Yang et JK Wu,Chinese Wild Plant Resources 21(5):2,2002.
=Vatica hainanensisH.T.Chang et L.C.Wang var.glandipetalaL.C.Wang Acta Scientiarum Naturalium Universitatis Sunyatseni,24(3):96,1985;Vatica mangachapoiBlanco var.glandipetala(LC Wang)YK Yang et JK Wu,Chinese Wild Plant Resources 21(5):1,2002.
=Vatica mangachapoiBlanco var.wanningensisGA Fu et YK Yang,Bulletin of Botanical Research 28(3):259,2008,syn.nov.
AcknowledgementsWe should like to thank the herbaria CANT,SYS and CFA for permission to study their specimens.We are also grateful to thank Dr.Hugh A.McAllister(Institute of Integrative Biology,University of Liverpool),Professor Shi-Liang Zhou and Dr.Qiang Wang(Institute of Botany,the Chinese Academy of Sciences)for their constructive suggestions and English correction.
References
Alan AB(2002)Population:a central concept for ecology?Oikos 97(3):439–442
Appanah S,Turnbull JM(1998)A review of dipterocarps:taxonomy,ecology and silviculture.Center for International Forestry Research,Bogor,p 195
Blanco M(1837)Flora de Filipinas:Segun el sistema sexual de Linneo.ed.1,Miguel Sanchez,Manila:Candido Lopez,the Philippines,401
Chen WQ(2005)Dipterocarpaceae.In:South China Botanical Garden,Chinese Academy of Sciences(ed)Flora of Guangdong,Vol.3:hernandiaceae through begoniaceae.Guangdong Science and Technology Press,Guangzhou,pp 158–159
Frampton C,Ward J(1990)The use of ratio variables in systematics.Taxon 39(4):586–592
Fu GA,Yang YK(2008)A new variety ofVaticaL.(Dipterocarpaceae)and a new combination ofcanthium(Rubiaceae)from Hainan Island.Bull Bot Res 28(3):259–260
Hsu PS(1998)The species problem in plant taxonomy in China.Acta Phytotaxon Sin 36(5):470–480
Li XW,Li J,Ashton PS(2007)Dipterocarpaceae.In:Wu ZY,Raven PH (eds),Flora of China,vol 13:Alangiaceae through Violaceae.Science Press,Beijing and Missouri Botanical Garden Press,St.Louis,pp.48–54
Nei M(1972)Genetic distance between populations.Am Nat 106:283–292
Nooteboom HP(1992)A point of view on the species concept.Taxon 41:318–320
Qin YH,Zhang J,Cui JT,Guo ZK,Jiang N,Tan RX,Ge HM(2011)Oligostilbenes fromVatica mangachapoiwith xanthine oxidase and acetylcholinesterase inhibitory activities. RSC Adv 1(1):135–141
Qu ZX(1956)The discovery ofVatica mangachapoion Hainan Island.Sci Silvae Sin 2(3):279–281
Schluter D(2001)Ecology and the origin of species.Trends Ecol Evol 16(7):372–380
Shi W,Pan B,Gaskin JF,Kang X(2009)Morphological variation and chromosome studies inCalligonum mongolicumandC.pumilum(Polygonaceae)suggests the presence of only one species.Nord J Bot 27(2):81–85
Wang LZ(1985)A study onVaticaplants growing in Hainan of Guangdong.Acta Sci Nat Univ Sunyatseni 24(3):94–97
Yang YK,Wu JK,Gong ZN(2002)A taxonomic research of the Dipterocarpaceae in China.Chin Wild Plant Resour 21(5):1–7
杂志排行

Journal of Forestry Research的其它文章
- In vitro propagation of conifers using mature shoots
- ‘Relationships between relationships’in forest stands:intercepts and exponents analyses
- Effects of application date and rate of foliar-applied glyphosate on pine seedlings in Turkey
- Assaying the allelopathic effects of Eucalyptus camaldulensis in a nursery bed incorporated with leaf litter
- Effects of soil compaction on growth variables in Cappadocian maple(Acer cappadocicum)seedlings
- Variation and selection analysis of Pinus koraiensis clones in northeast China