4种不同外源物对干旱胁迫下欧报春种子萌发的影响
2018-05-10郑云凤张晓曼赵家豪
郑云凤, 张晓曼, 赵家豪
(河北农业大学,河北保定 071000)
欧报春(Primulavularis)是报春花属中品种相对最多、花色最为丰富、应用最为广泛的一种植物,经过园艺学家长期选育,已成为重要的观赏植物之一[1]。欧报春作为世界早春园林多色草花,其应用价值十分广泛,市场潜力非常大。欧报春喜温凉湿润的环境,高温、干旱、严寒是制约欧报春在北方园林应用的三大因素,研究欧报春在干旱胁迫过程中的生理反应,对欧报春在园林上的应用有着重要的意义。目前,外源物质调节植物抗旱性和相关应用技术的研究相对较多,但有关欧报春受干旱环境胁迫的研究报道相对较少,尤其是关于外源物质缓解欧报春干旱胁迫伤害的研究相对更少。
欧报春一般通过种子繁殖,欧报春种子能否在干旱条件下萌发成苗,是其能否在北方成功引种的前提和关键条件。本试验以欧报春种子为材料,用5%聚乙二醇6000(PEG-6000)溶液模拟干旱胁迫,研究甜菜碱(GB)、赤霉素(GA)、氯化钙(CaCl2)、脱落酸(ABA)这4种外源物质不同浓度浸种处理对干旱胁迫下欧报春种子萌发的影响,了解欧报春种子萌发的抗旱性特征和4种外源物缓解干旱胁迫的最适宜浓度,为欧报春在实际生产栽培过程中遇到干旱胁迫问题提供解决方法,也为欧报春在北方地区的种植提供理论依据,更为我国野生报春花引种过程中提高其抗旱性提供参考和指导。
1 材料与方法
1.1 试验材料
欧报春种子,由四川德阳东方花卉有限公司提供;聚乙二醇6000、甜菜碱、赤霉素、氯化钙、脱落酸,市购。
1.2 试验方法
1.2.1 浸种 本试验于2016年8月在河北农业大学园林遗传育种实验室进行,将颗粒饱满、大小一致的欧报春种子用70%乙醇消毒,冲洗干净。将欧报春种子分别置于含有浓度为100、200、300、400、500 mg/L甜菜碱,10、50、100、150、200 mg/L 赤霉素,10、50、100、150、200 mg/L氯化钙,1、2、4、6、8 mg/L脱落酸4种不同外源物的培养皿中浸泡,以双蒸水、5% PEG-6000浸泡分别作为对照1(CK1)、对照2(CK2)。将浸种过的种子置于培养箱中培养24 h,温度 20 ℃,光照12 h/d;用蒸馏水冲洗3次,备用。
1.2.2 干旱胁迫萌发试验 在培养皿中底部铺1层纱布、2层滤纸作为种子发芽床,用配好的5% PEG-6000溶液浸透培养皿里的纱布和2层滤纸;将经过浸泡处理的种子均匀摆放在培养皿中,每个培养皿放50粒种子,重复3次;放入温度(20±1)℃、光照度6 000 lx、光照时间12 h/d的培养箱中培养,早晚各观察1次,以称质量法补充培养皿中的蒸馏水来维持溶液渗透势的稳定。
1.3 测定项目与方法
按照《国际种子检验规程》中以胚根突出种皮的长度为种子长度一半的种子视为发芽种子,从种子置于培养箱中第2天开始,到种子发芽后第20天结束,每天同一时间记录种子的发芽情况,测量萌发种子的胚根、胚轴长度,统计萌发率(G)、发芽指数(Gi)、生长速率(Rv)、活力指数(Iv)、发芽势(GE),计算公式分别为:G=n/N×100%;Gi=∑(Gt/Dt);Rv=L/∑[Ni×(D20-Di+0.5)];Iv=s×Gi;GE=n13/N×100%。式中:n为萌发种子数,粒;N为供试种子数,粒;Gt为第t天的萌发种子数,粒;Dt为相应的萌发时间,d;L指每皿中全部萌发种子的胚根、胚轴长度的总和,mm;Ni指第i天的萌发种子数,粒;D20指试验持续的时间,本试验为20 d;Di指第i天;s为种子胚根长,mm;n13为种子萌发相对比较集中、第13天的正常种子发芽数,粒。
1.4 数据分析
采用Excel软件对试验数据进行统计,应用SPSS 17.0软件进行显著性分析。
2 结果与分析
2.1 GB浸种处理对干旱胁迫下欧报春种子萌发的影响
由表1可知,与双蒸水处理的正常种子(CK1)相比,5% PEG-6000干旱胁迫处理(CK2)的欧报春种子萌发率有显著降低(P<0.05),下降了51.33百分点,表明PEG-6000处理显著抑制了欧报春种子的正常萌发;GB不同浓度浸种处理都能明显提高欧报春种子在干旱胁迫下的萌发率,提高了欧报春种子的抗旱能力,其中,GB浓度为400 mg/L时种子萌发率相对最高,较CK2高出31.94百分点,差异显著(P<0.05);GB浓度为100~400 mg/L时,随GB浓度的增大,欧报春种子萌发率呈逐渐上升趋势,且相互间差异不显著(P>0.05)。
发芽指数、活力指数比单纯的发芽率更能全面反映种子在实际条件下的萌发速度、整齐度及种子萌发数目和幼苗健壮生长的潜势[2-3]。由表1可见,CK2处理的发芽指数、活力指数分别比CK1下降1.18、160.39,5% PEG-6000处理使欧报春种子的发芽指数、活力指数有显著降低(P<0.05);CK2处理的发芽指数、活力指数明显低于GB不同浓度处理,但相互间差异不显著(P>0.05);GB浓度为300 mg/L时,欧报春种子的发芽指数、活力指数均达到最高值,分别高出CK20.46、26.70;GB浓度为100~500 mg/L时,随GB浓度的增大,欧报春种子发芽指数与活力指数呈先上升后下降趋势。因此可见,GB对干旱胁迫下欧报春种子的发芽指数和活力指数有促进作用,当GB浓度为300 mg/L时对种子萌发数、速度及整齐度作用相对最大。
种子萌发后,胚根的延伸可以反映植物定居成苗的特性[4]。由表1可见,GB处理对干旱胁迫下欧报春种子生长速率有明显影响;种子经不同浓度GB处理后,随GB浓度的增大,欧报春种子生长速率呈先增加后降低趋势,在GB浓度为 300 mg/L 时达到最大值,为2.71 mm/d,比CK2高出 0.42 mm/d,表明GB能提高干旱胁迫下欧报春种子的生长速率,加快种子胚根的生长。
发芽势能反映种子的发芽速度、发芽整齐度及种子生活力的强弱程度[5]。由表1可见,不同浓度GB处理对干旱胁迫下欧报春种子发芽势有明显影响,均高于CK2,说明GB能提高干旱胁迫下欧报春种子的发芽势,能有效提高欧报春种子的发芽速度和生活力,提高发芽整齐度;随GB浓度的增大,欧报春种子发芽势呈先升后降再升趋势,浓度为300 mg/L时,欧报春种子的发芽势达到最大值,为29.33%,比CK2高12.00百分点。
综上,添加GB浓度为300 mg/L时,发芽指数、生长速率、活力指数及发芽势都达到最大值,该条件下种子发芽率为52.00%,仅次于浓度为400 mg/L时的最高发芽率53.94%,因此添加浓度为300 mg/L的GB对干旱胁迫下的欧报春种子萌发有较好的缓解作用。
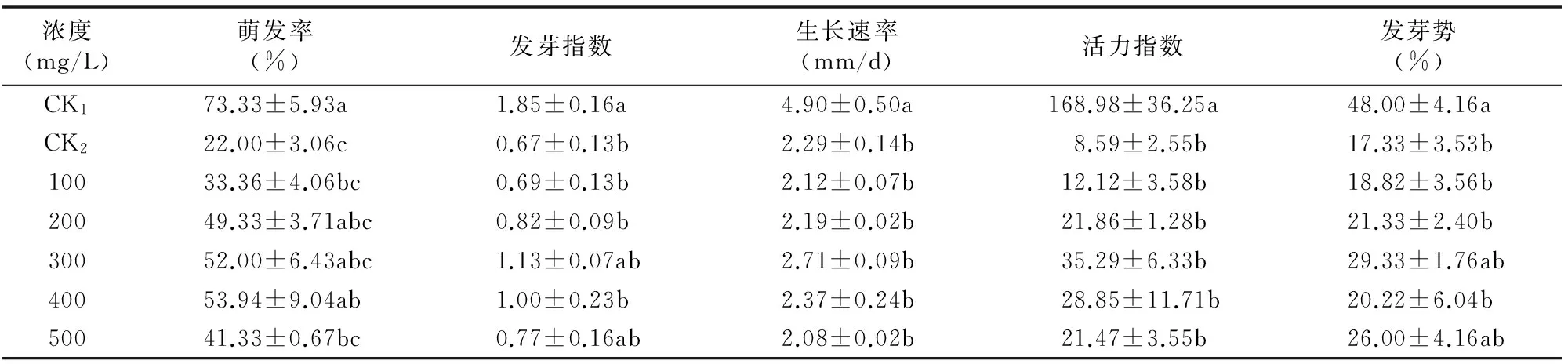
表1GB对干旱胁迫下欧报春种子萌发指标的影响
注:表中数据为“平均值±标准误”;同列数据后不同小写字母表示处理间差异显著(P<0.05)。下表同。
2.2 GA浸种处理对干旱胁迫下欧报春种子萌发的影响
由表2可知,不同浓度GA处理对干旱胁迫下欧报春种子萌发有明显影响。随GA浓度的增大,干旱胁迫下欧报春种子的萌发率、发芽指数、活力指数、发芽势均呈逐渐降低趋势;GA浓度为10 mg/L时,干旱胁迫下欧报春种子的萌发率、发芽指数、生长速率、活力指数、发芽势相对最高,分别为 65.33%、1.97、3.51 mm/d、134.10、51.33%,分别比CK2高43.33百分点、1.30、1.22 mm/d、125.51、34.00百分点,而与CK1相比,其发芽指数和发芽势分别比CK1高0.12和3.33百分点,相互间差异不显著(P>0.05),说明低浓度GA能明显提高干旱胁迫下欧报春种子的萌发率、发芽势和生长速率,加快种子胚根的生长,能有效提高欧报春种子的发芽速度和生活力,增强发芽整齐度;当GA浓度达到150 mg/L时,干旱胁迫下欧报春种子的萌发率、发芽指数、生长速率、活力指数、发芽势分别比CK2低4.67百分点、0.21、0.32 mm/d、4.39、5.33 百分点,且与CK2相比差异不显著(P>0.05),表明较高浓度的GA反而会抑制种子的萌发和生长。
2.3 CaCl2浸种处理对干旱胁迫下欧报春种子萌发的影响
由表3可知,不同浓度CaCl2浸种对干旱胁迫下欧报春种子的萌发率、发芽指数、活力指数、生长速率、发芽势影响不显著(P>0.05);随CaCl2浓度的增大,干旱胁迫下欧报春种子的萌发率、生长速率、活力指数、发芽势均呈先升后降趋势,其中CaCl2浓度为50 mg/L时,干旱胁迫下欧报春种子的萌发率、发芽指数、生长速率、活力指数、发芽势相对最高,分别为42.98%、0.92、2.24 mm/d、18.95、24.35%,除生长速率外其他几个指标分别比CK2高22.98百分点、0.25、10.36、7.02百分点,生长速率比CK2低0.05 mm/d,但相互间差异不显著(P>0.05);CaCl2浓度为200 mg/L时,干旱胁迫下欧报春种子的萌发率、发芽指数、生长速率、活力指数、发芽势相对最低,分别为31.33%、0.68、2.04 mm/d、13.67、17.69%,但欧报春种子的萌发率、发芽指数、活力指数、发芽势仍分别比CK2高9.33百分点、0.01、5.08、0.36百分点。可以看出,CaCl2能促进干旱胁迫下欧报春种子的萌发,提高种子的发育速度和生活力,以CaCl2浓度为 50 mg/L 时的效果最为明显。
2.4 ABA浸种处理对干旱胁迫下欧报春种子萌发的影响
由表4可知,不同浓度ABA浸种对干旱胁迫下欧报春种子萌发的影响有明显不同;随ABA浓度的增大,干旱胁迫下欧报春种子的萌发率、发芽指数、活力指数、发芽势均呈先升后降趋势,其中ABA浓度为2 mg/L时,干旱胁迫下欧报春种子的萌发率、发芽指数、生长速率、活力指数、发芽势达到最高值,分别为55.33%、1.64、3.92 mm/d、74.10、42.67%,分别比CK2高33.33百分点、0.96、1.63 mm/d、65.51、25.34百分点,除萌发率与CK2相比差异显著(P<0.05)外,其他指标与CK2差异不显著(P>0.05);当ABA浓度为8 mg/L时,干旱胁迫下欧报春种子的萌发率、发芽指数、生长速率、活力指数、发芽势相对最低,分别为19.33%、0.64、2.15 mm/d、7.26、16.67%,分别比CK2低2.67百分点、0.04、0.14 mm/d、1.33、0.66百分点,说明高浓度脱落酸会抑制干旱胁迫下欧报春种子的萌发,影响干旱胁迫下欧报春种子的萌发率、发芽指数、活力指数、发芽势,从而影响欧报春种子萌发的速度及整齐度。
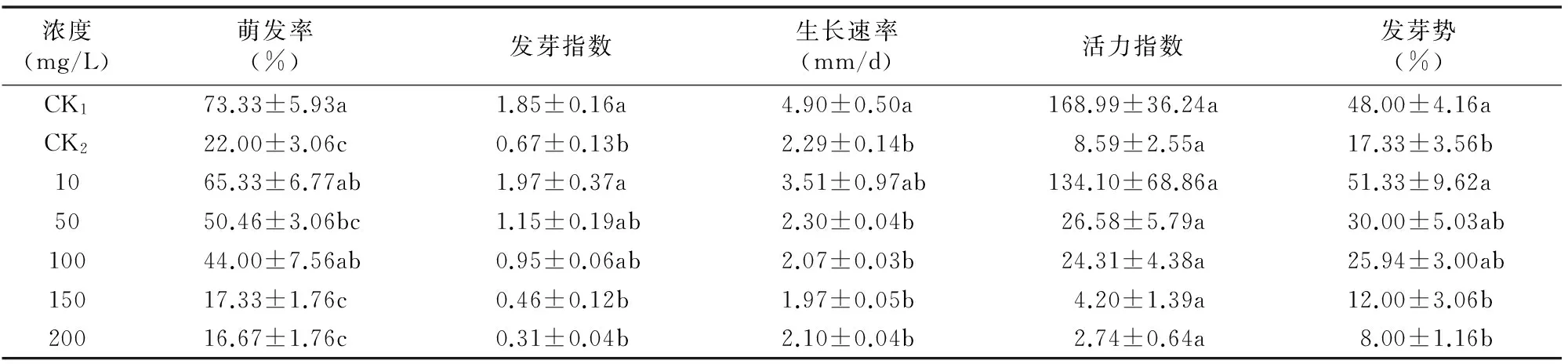
表2GA对干旱胁迫下欧报春种子萌发指标的影响

表3CaCl2对干旱胁迫下欧报春种子萌发指标的影响
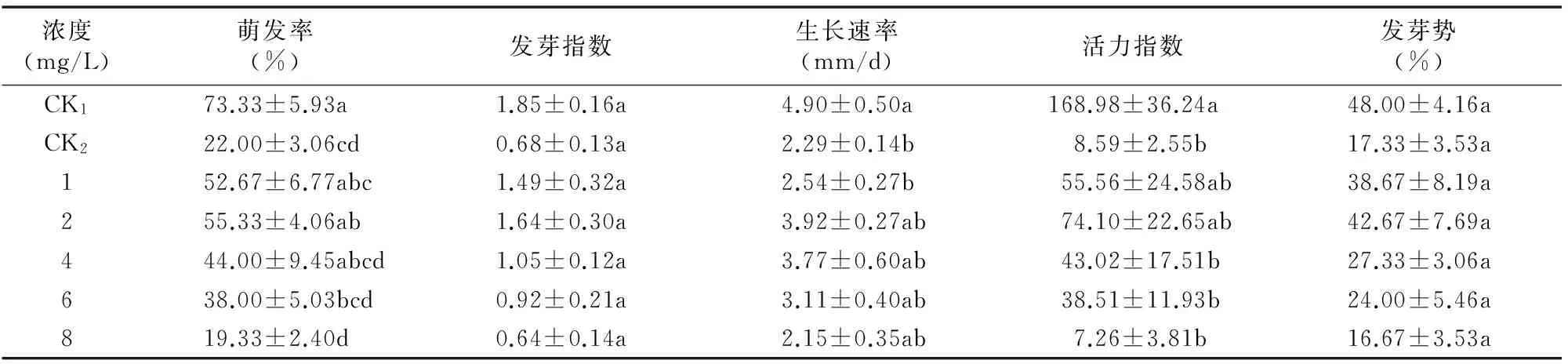
表4ABA对干旱胁迫下欧报春种子萌发指标的影响
3 结论与讨论
与双蒸水正常浸种(CK1)相比,5%聚乙二醇6000(PEG-6000)模拟干旱胁迫对欧报春种子的萌发有明显的抑制作用,干旱胁迫下欧报春种子的萌发率、发芽指数、生长速率、活力指数、发芽势均有显著降低(P<0.05),而甜菜碱、赤霉素、氯化钙和脱落酸这4种外源物在适宜浓度下浸种能明显缓解干旱胁迫对欧报春种子萌发的抑制作用,与干旱胁迫(CK2)处理的欧报春种子相比,其萌发率、发芽指数、生长速率、活力指数、发芽势等各项指标有较为明显的提高。缓解干旱胁迫抑制作用效果相对较好的有甜菜碱浓度300 mg/L、赤霉素浓度10 mg/L、氯化钙浓度50 mg/L、脱落酸浓度2 mg/L。
甜菜碱属季铵型水溶性生物碱,是已研究过的150多种代谢物中最有效的渗透调节剂之一[6],参与细胞的渗透调节,当植物受干旱胁迫时,它能降低细胞的渗透势,保护和维持生物大分子的结构和完整性,有效缓解干旱胁迫对植物造成的伤害[7]。张艳等研究发现,甜菜碱浓度为400 mg/L时,能明显改善干旱胁迫下白三叶种子的各发芽指标[8];李小白研究表明,甜菜碱浓度为1.5 mmol/L时,能提高干旱胁迫下谷子的发芽势和发芽率[9];王娟研究发现,甜菜碱浓度为 20 mmol/L 时,对干旱胁迫下鸡冠花种子的萌发有明显的缓解作用[10];刘海英研究指出,甜菜碱浓度为1 mmol/L时最能有效缓解干旱胁迫下桔梗种子萌发的抑制作用,且随甜菜碱浓度的升高,缓解作用减弱,反而会抑制干旱胁迫下种子的萌发率[11]。本研究结果表明,甜菜碱浓度为300 mg/L时,能缓解种子在PEG干旱胁迫下的抑制作用,并随甜菜碱浓度增大到一定程度,反而会抑制干旱胁迫下种子的萌发。
有研究表明,赤霉素参与植物生长发育的全过程,如种子萌发、茎的伸长、花的诱导和发育、种子和果实的生长等[12-14]。在种子萌发过程中,GA能破除种子的休眠,促进淀粉酶和其他水解酶的合成,并对受损的细胞膜起到一定的修复作用[15]。此外,GA在幼苗抗逆性形成方面也具有重要作用,这可能和GA可诱导与抗逆性形成相关的基因表达和蛋白质合成有关[16-17]。代勋等研究发现,用10 mg/L赤霉素浸种小桐子种子,可以提高干旱胁迫下小桐子种子的发芽率[18];杨阳等发现,赤霉素浓度为200 mg/L时,对干旱胁迫后的沙冬青种子萌发有促进作用[19];坎平等研究发现,0.5 mmol/L 赤霉素处理能提高烟草种子的抗旱性[20];任永霞等发现,50 mg/L赤霉素浸种能促进干旱胁迫下紫花苜蓿种子的萌发[21]。本试验结论与代勋等的研究结论[18]较为吻合,赤霉素浓度为10 mg/L时,可缓解干旱胁迫对欧报春种子萌发的抑制作用。
钙不仅是一种大量元素,也是一种第二信使,各种逆境胁迫初期都会诱发细胞质中Ca2+水平的增加,形成具有不同时空特点的Ca2+指纹,这些Ca2+指纹通过钙调素(CaM)、钙依赖蛋白激酶(CDPK)、促细胞分裂原活化蛋白激酶(MAPK)等解码途径诱发植物耐逆性的形成[15-17]。因此,Ca2+在植物感受、转导和响应低温、干旱等逆境胁迫过程中起到重要作用。李小白研究发现,氯化钙浓度为10 mmol/L(1 110 mg/L)时能提高干旱胁迫下谷子的发芽势和发芽率[9];代勋等研究发现,用氯化钙浓度为10 mmol/L浸种小桐子种子,可以提高干旱胁迫下小桐子种子的发芽率[18];任永霞等发现,用 5 mmol/L(555 mg/L)氯化钙浸种能促进干旱胁迫下紫花苜蓿种子的萌发[21]。本试验研究结果与李小白等的研究结果[9,18,21]差异较大,说明促进干旱胁迫下种子萌发的最佳氯化钙浓度因植物种类不同而有所差异,但氯化钙浓度在 1~10 mmol/L 之间是否存在促进种子萌发的最适浓度还有待进一步研究。
目前,大量研究表明,当植物遭到干旱、低温、盐害等环境胁迫时,细胞会迅速积累ABA[22],产生这种现象的原因可能是逆境胁迫增加了叶绿体膜对ABA的通透性,加快了根系合成ABA向叶片运输及积聚,而ABA能调节气孔开度,减少蒸腾失水,缓解干旱,能引起某些相关基因的表达,这些使植物对干旱的适应至关重要。同样,外源ABA使植物对逆境也有缓解作用[23]。彭浩等研究发现,脱落酸浓度为4 mg/L时,具有显著缓解玉米种子干旱胁迫的效果[24],而本研究结果与其存在差异,说明脱落酸缓解干旱胁迫下种子萌发的浓度因植物种类不同而可能有差别。
参考文献:
[1]黄祯强. 欧报春的温室盆栽试验及在上海园林中的应用[J]. 中国园艺文摘,2011,27(6):28-29.
[2]王娟娟,张文辉,刘新成. NaCl胁迫对3种不同处理四翅滨藜种子萌发的影响[J]. 西北农业学报,2010,19(1):104-111.
[3]Zhang H X,Zhou D W. Research on seed germination ecology[J]. Acta Agrestia Sincis,2009,17(1):131-133.
[4]张洁明,孙景宽,刘宝玉,等. 盐胁迫对荆条、白蜡、沙枣种子萌发的影响[J]. 植物研究,2006,26(5):595-599.
[5]廉英杰. 提高陈辣椒种子发芽势和发芽率的方法[J]. 新疆农业科技,2013(4):42-43.
[6]Sakamoto A,Murata N. Genetic engineering of glycinebetaine synthesis in plants:current status and implications for enhancement of stress tolerance[J]. Journal of Experimental Botany,2000,51(342):81-88.
[7]Robinson S P,Jones G P. Accumulation of glycine betaine in chloroplasts provides osmotic adjustment during salt stress[J]. Functional Plant Biology,1986,13(5):659-668.
[8]张 艳,彭 燕,何小双. 甜菜碱浸种对干旱胁迫下白三叶种子萌发特性的影响[J]. 中国草地学报,2014,36(4):31-37.
[9]李小白. 干旱条件下外源CaCl2、海藻糖和甜菜碱对谷子萌发及幼苗生长的影响[D]. 晋中:山西农业大学,2013.
[10]王 娟. 外源硝普钠、甜菜碱及腐殖酸对干旱胁迫下下鸡冠花种子萌发和幼苗生理的影响[D]. 重庆:西南大学,2014.
[11]刘海英. 外源ALA、SNP和GB对干旱胁迫桔梗种子萌发及幼苗生理生化特性的影响[D]. 重庆:西南大学,2013.
[12]Hedden P,Phillips A L. Manipulation of hormone biosynthetic genes in transgenic plants[J]. Current Opinion in Biotechnology,2000,11(2):130-137.
[13]Brooking I R,Cohen D. Gibberellin-induced flowering in small tubers ofZantedeschia‘Black Magic’[J]. Scientia Horticulturae,2002,95(1/2):63-73.
[14]Joly C,Maftah A,Riou-Khamlichi C. Alteration of gibberellin response in transgenic tobacco plants which express a human Lewis fucosyltransferase[J]. Plant Physiology and Biochemistry,2004,42(7/8):629-637.
[15]刘永庆,Bino R J,Karssen C M. 赤霉素与脱落酸对番茄种子萌发中细胞周期的调控[J]. 植物学报,1995,37(4):274-282.
[16]吕 彪,许耀照,王治江,等. 聚乙二醇胁迫下赤霉素浸种对番茄种子萌发和幼苗生长的影响[J]. 干旱地区农业研究,2009,27(4):136-139.
[17]温福平,张 檀,张朝晖,等. 赤霉素对盐胁迫抑制水稻种子萌发的缓解作用的蛋白质组分析[J]. 作物学报,2009,35(3):483-489.
[18]代 勋,李忠光,龚 明. 赤霉素、钙和甜菜碱对小桐子种子萌发及幼苗抗低温和干旱的影响[J]. 植物科学学报,2012,30(2):204-212.
[19]杨 阳,刘秉儒,贾倩民,等. 赤霉素对干旱胁迫下沙冬青种子萌发的影响[J]. 江苏农业科学,2014,42(5):271-274,275.
[20]坎 平,王莎莎,马文广,等. 赤霉素引发同时提高烟草种子及幼苗抗旱性和抗冷性[J]. 种子,2014,33(2):30-34,38.
[21]任永霞,郭郁频. 钙和赤霉素浸种对干旱胁迫下紫花苜蓿种子萌发的影响[J]. 黑龙江畜牧兽医,2016,7(1):138-141.
[22]李 畅,苏家乐,肖 政,等. 旱涝交替胁迫下杜鹃花叶片内源激素含量的变化[J]. 江苏农业科学,2016,44(7):233-235.
[23]曾 荣,邵 闰,杨 娟,等. 嫁接和喷施抗寒剂对三角梅抗寒性的影响[J]. 江苏农业科学,2016,44(1):202-204.
[24]彭 浩,王晓强,赵 强,等. 水杨酸与脱落酸对干旱胁迫下玉米种子萌发的影响[J]. 济宁学院学报,2015,36(3):72-77.
