尖吻鲈仔、稚鱼异速生长研究
2018-05-09刘亚娟程大川周胜杰马振华
刘亚娟,程大川,周胜杰,马振华,胡 静,杨 蕊
(1.天津农学院,天津 300384;2.中国水产科学研究院南海水产研究所热带水产研究开发中心,海南三亚 572018;3.农业部南海渔业资源开发利用重点实验室,广州 510300)
异速生长,又名相对生长,即指生物个体某一类功能器官的相对生长速率不同于身体其他部分的相对生长速率的现象,是长期以来动植物个体形态建成过程中为适应复杂的外界环境而保留的发育规律[1-2]。在硬骨鱼类早期的生长发育阶段里,受到遗传和环境因素的影响,仔、稚鱼各功能器官的发育表现出不同步性[3]。早期阶段鱼类的生长发育中普遍保留有异速生长规律,例如在美洲鲥鱼(Alosa sapidissima)[4]、大麻哈鱼(Oncorhynchus keta Walbaum)[5]、红 鳍 笛 鲷(Lutjanus erythopterus)[6]、粗 唇 龟 鲻 (Chelon labrosus)[7]等物种的仔、稚鱼阶段中,重要头部器官都会优先发育,为眼、口的快速分化创造条件,提高呼吸及摄食能力,使其适应繁复多变的环境。
尖吻鲈(Lates calcarifer),别称金目鲈、盲曹,分类学上归于硬骨鱼纲,鲈形目,鲈亚目,锯盖鱼科,尖吻鲈属,是澳大利亚、印度、东南亚等热带地区咸淡水、网箱及池塘养殖的常见经济鱼类。尖吻鲈肉质细嫩、口感鲜美、生长迅速,具有较高的营养价值及保健功效[8-12]。近几年来,广州、福建、珠海、海南等沿海城市尖吻鲈养殖产业发展迅速,具有显著的经济效益。目前国内外对尖吻鲈仔、稚鱼的研究主要集中在基因表达[13]、捕食动力学[14]以及鱼苗培育[15]等方面,而对于其早期发育阶段中相关功能器官、结构的生长模式还没有系统的研究。因此,本研究对尖吻鲈仔、稚鱼各主要器官的可量指标进行测量分析,确定其异速生长情况,并从仔、稚阶段器官的发生、分化及形态的差异推断异速生长的原因,旨在丰富尖吻鲈发育过程的基础生物学资料,并为其苗种的培育技术提供实践指导。
1 材料与方法
1.1 实验材料
尖吻鲈受精卵购于海南陵水县,运至中国水产科学研究院南海水产研究所热带水产研究开发中心育苗车间。首先将受精卵在水温27.5℃下平衡20 min,之后轻缓地加入到500 L容量的孵化器中。至仔鱼2日龄时,将其从孵化器中分别转移到3个相同的2 500 L容量的育苗桶中,自此进入了仔、稚鱼饲养阶段,保持桶内鱼苗密度为每桶60个·L-1。用300μm孔径纱网拦截幼苗,调节苗桶阀门,日换水量为苗桶容量的2倍,每日固定清理排水口过滤网。光照条件为每日2 000流明,盐度保持在33±0.8(Mean±SD),水温控制在(29.0±1.0)℃范围内。每桶放置4个气石,同时为减小气泡破裂力对幼苗的伤害,要求气量需调节的恰到好处。
从尖吻鲈2日龄开始投喂轮虫,轮虫密度保持于10~20个·mL-1到10日龄时停止投喂。9日龄起开始投喂丰年虫的无节幼体,初始密度为0.1个·mL-1,之后逐日递增直到5个·mL-1。19日龄起开始驯化尖吻鲈进食微颗粒饲料[16],每日8∶30到19∶00期间每隔1 h加入适口的浮性颗粒饲料,根据鱼苗对微颗粒饲料(Otohime A1 Marubeni Nisshin Feed Co.,Ltd.Tokyo,Japan)的进食情况酌情递减投喂量,直到驯化完全。前期在投喂生物饵料时,于投喂前需对生物饵料(轮虫和丰年虫无节幼体)进行12 h营养强化,强化剂品牌来自INVE Aquaculture。育苗阶段水体中加入海水拟微球藻(Nannocholoropsis sp.,青岛宏邦生物技术有限公司),以保证育苗桶中的轮虫能够有足够饵料,同时为尖吻鲈仔、稚鱼提供绿色的水体环境。育苗过程中,采用虹吸的方法以去除苗桶中的排泄物、剩余饵料和死去的仔、稚鱼。实验至36日龄结束,体长为56.26±7.02(Mean±SD)mm。
1.2 研究方法
采用体视显微镜(Olympus SZ40型)观察以及One-plus A3010相机拍照采集信息并作记录。自仔鱼孵化后第1日龄起,每日8∶00从桶内随机抽取10个样本,麻醉剂浓度为40 mg·L-1,麻醉剂品牌为AQYI-S(New Zealand Ltd),麻醉后用体视显微镜观察其各相关器官的形态变化并拍照记录。
图1中所示躯干、尾部、头部的各个指标[6]使用Auto CAD 2014测量,图片标尺精确至0.01 mm,测量后,采用浓度为10%的中性福尔马林溶液将全部样本进行固定,于避光环境下保存以备查用。

图1 尖吻鲈仔、稚鱼测量图Fig.1 Illustration measurement for Lates calcarifer注:1.吻长;2.眼径;3.头高;4.头长;5.上颌长;6.躯干长;7.尾鳍长;8.体高;9.背鳍长;10.胸鳍长;11.腹鳍长;12.臀鳍长;13.全长Note:1.Rostrum length;2.Eye diameter;3.Head height;4.Head length;5.Upper jaw length;6.Trunk length;7.Tail fin length;8.Body height;9.Dorsal fin length;10.Pectoral length;11.Pelvic fin length;12.Anal fin length;13.Total length
1.3 数据处理
用数学模型y=a ebx来拟合全长与日龄的函数关系[17]。式中x为日龄,y为该日龄下所对应的全长,b代表生长速率,a则为y轴的截距。鱼类异速生长模型采用幂函数方程y=axb,仔、稚鱼全长用x表示,用y表示x相应的各个器官的长度,其中a是曲线在y轴的截距,而b是异速生长指数。当b值大于1的情况下,功能器官表现出正异速速增长(快速生长);当b值小于1时,功能器官表现为负异速增长(慢速生长);当b值为1时,器官呈现等速生长。功能胃的形成是稚鱼期到来的标志[18],尖吻鲈在16 d时胃腺开始发育,因此,以16日龄(day post hatch,DPH)作为节点将尖吻鲈生长曲线划分,不同的生长阶段采用不同生长方程来表示:y=a1xb1,y=a2xb2。对b1、b2是否等于1做 t检验,此外对 b1、b2做 t检验,以检测两个 b值是否差异显著(置信区间95%)。使用SPSS 18.0软件对生长模型进行拟合非线性回归,并使用EXCEL 2016进行拟合分段回归模型,以残差平方和最小和决定系数R2最大为曲线拟合的标准,显著性检测使用SPSS 18.0进行。
2 结果与分析
2.1 尖吻鲈仔、稚鱼全长和日龄的关系
尖吻鲈仔、稚鱼全长与日龄的关系方程为y=1.692 1×e0.1131x,其中R2为0.939 0,拟合曲线可靠性较高(图2)。1日龄尖吻鲈仔鱼全长为(2.2±0.52)mm,经过 36 d生长,全长达到(56.26±7.02)mm,平均增长率为1.501 1 mm·d-1。

图2 尖吻鲈仔、稚鱼日龄与全长的关系Fig.2 Correlation between total length,day post hatch of larval and juvenile Lates calcarifer
2.2 尖吻鲈仔、稚鱼发育过程中各功能器官的异速生长
2.2.1 尖吻鲈头部各器官的异速生长
尖吻鲈仔、稚鱼的吻长(图3-a)、口宽(图3-b)、眼径(图3-c)、头高(图3-d)与全长之间出现了异速生长的情况。16日龄之前,它们的异速生长指数b1分别为1.236 4、1.071 6、1.079 6、0.965 6,吻长、口径、头高三个指标的b值与1均具有显著性差异(P<0.05),正异速生长得以表现。眼径的异速生长指数b1与1差异不显著,表现为等速生长。16日龄后,b2分别为1.104 3、1.105 6、0.609 8、0.806 7,吻长、口径、头高、眼径的b2值与1均具有显著性差异(P<0.05),吻长、口径表现为正异速生长,头高、眼径表现为负异速生长。故在16日前后,尖吻鲈的吻长、口径均表现出正异速生长,头高在16日前表现出正异速生长,16日后转化为负异速生长,16日前尖吻鲈眼径表现为等速生长,16日龄后为负异速生长。16日龄时,尖吻鲈全长为(17.60±1.77)mm,分析发现可得出吻长、口径、头高和眼径在此时均存在生长拐点。

图3 尖吻鲈仔、稚鱼头部各器官异速生长曲线Fig.3 Allometric growth curve of head functional organs in larval and juvenile Lates calcarifer
2.2.2 尖吻鲈仔、稚鱼身体各部分的异速生长
尖吻鲈身体部分的异速生长曲线主要包括头长(图4-a)、体高(图4-b)、躯干长(图4-c)。头长的异速生长指数 b1为1.115 8,b2为1.051 7,与1均显著性差异(P<0.05),表现为正异速生长。体高的异速生长指数 b1为1.012 8,b2为1.046 5,与 1无差异显著(P>0.05),表现为等速生长。躯干长的异速生长指数b1为1.048 3,b2为1.019 8,与1无显著差异(P>0.05),等速生长规律得以表现。故说明尖吻鲈头长在仔鱼期、稚鱼期均为快速生长;体高、躯干长在仔鱼期、稚鱼期均为等速生长。16日龄时,尖吻鲈全长为(17.60±1.77)mm,通过异速生长指数显著性的差异,实际得出头长此时存在生长拐点,体高、肛前长不存在生长拐点。
2.2.3 尖吻鲈仔稚鱼游泳器官的异速生长
尖吻鲈仔、稚鱼的外部游泳器官主要为背鳍(图5-a)、腹鳍(图5-b)、胸鳍(图 5-c)、臀鳍(图5-d)、尾鳍(图 5-e),起着导向、平衡及推进的作用。尖吻鲈仔鱼期的尾鳍的异速生长指数b1为0.879 7,与1均呈现显著性差异(P<0.05),负异速生长规律得以呈现。胸鳍的异速生长指数b1为1,表现为等速生长。稚鱼期,尾鳍的异速生长指数b2为1.030 1,与1无显著差异(P<0.05),表现为等速生长,胸鳍的异速生长指数b2为1.086 9,与1具有显著性差异(P<0.05),表现出正异速生长。分析两者的b1和b2后,发现尖吻鲈尾鳍、胸鳍在16日龄均存在生长拐点,前后表现出不同的生长速度。
尖吻鲈的背鳍的测量在14日龄左右才开始,腹鳍、臀鳍在16日龄时才能准确测量。背鳍、腹鳍、臀鳍的异速生长指数 b值分别为1.067 3、1.083 3、1.152 0,与 1均差异性显著(P<0.05),均表现为正异速生长。

图4 尖吻鲈仔、稚鱼身体各部分异速生长曲线Fig.4 Allometric growth curve of body sections of larval and juvenile Lates calcarifer
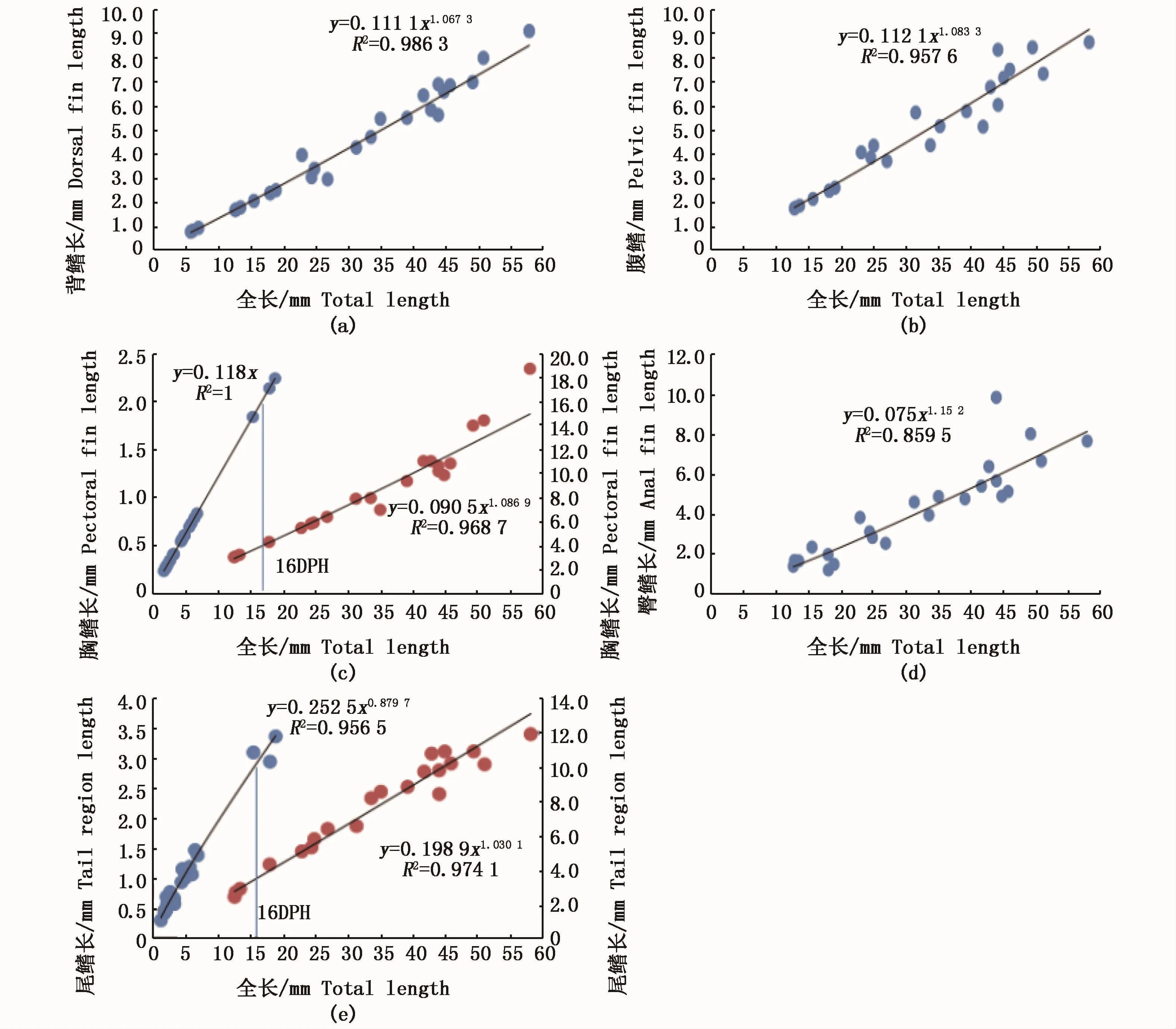
图5 尖吻鲈仔、稚鱼游泳器官异速生长曲线Fig.5 Allometric growth curve of swimming functional organs of larval and juvenile Lates calcarifer
3 讨论
仔、稚鱼阶段在鱼类整体生命过程中是高死亡率阶段,也是其对外界压力和环境因子最为敏感的时期[19]。在硬骨鱼类发育的早期阶段,自然水域中天敌捕食与饥饿是影响仔、稚鱼的成活率和资源补充的重要因素[20-21]。骨骼畸形、开口饵料不适口、寄生虫等是鱼苗繁育过程中的重要挑战,另外,寻找合适的养殖密度、光照强度、溶氧量以及活体饵料等均是鱼类养殖的关键环节[22-27]。在仔、稚鱼不同的发育阶段,由于个体发育的生理因素以及环境因素(溶氧、pH等)的影响,在形态学、生理学等方面,各种关键器官(如头、口、鳍等)都经历了显著的变化[26],那些对早期存活发挥重要作用的器官得以选择性地优先发育,使其获得适应于环境的最佳性能,并且这种优先性会随着发育阶段的不同而做出改变。这种优先发育规律是鱼类在长期进化历程中演化的自我保护机制,表现为与主动摄食及躲避敌害相关的一些关键器官优先发育。在美洲鲥[4]、大麻哈鱼[5]、红鳍笛鲷[6]、粗唇龟鲻[7]等物种中也观测到了这种异速生长的发育机制,与本研究的结果一致。这种优先发育机制直接影响到尖吻鲈仔、稚鱼的捕捉信息,运动生长及同化作用等生理和生态学活动,最终关系到鱼类躲避敌害、繁衍种群的能力,达到与饲养环境的协调[27]。
3.1 器官发育
尖吻鲈仔、稚鱼的头部形态学参数(眼径、吻长、上颌长、头长)相对于全长均具有异速生长的特点,说明其头部与仔、稚鱼整体亦呈异速生长的关系。仔、稚鱼阶段摄食功能的强弱与口器功能密切相关,口裂大小直接影响取食饵料的机能,口宽决定捕获饵料的大小,食性及所处发育阶段影响其口宽比[28]。仔、稚鱼的口宽变化及变态对于仔、稚鱼选择适口饵料以及食性的转变至关重要[29-30]。受精卵在孵化后,随着卵黄囊内营养物质的不断消耗以及油球的吸收,仔鱼开口后,进入混合营养阶段,向外界摄食的压力不断增大,口裂的快速生长也是其对于不同大小食物的适应[31-33]。本实验中,尖吻鲈吻长、上颌长前后都表现为快速生长。仔、稚鱼早期的生长发育和饵料类型与其营养结构密切相关。在尖吻鲈早期发育过程中,自2日龄末开始张口,投喂轮虫,10日龄停止投喂,随着仔鱼的生长,9日龄开始投喂营养更丰富的丰年虫无节幼体,19日龄开始过渡到营养全面的微颗粒饲料。通过一系列的饵料转变,尖吻鲈获得了充足的营养源,保证了各器官的快速发育,而在16日龄的拐点前后,尖吻鲈的吻长、上颌长在仔、稚鱼阶段一直处于优先发育,上、下颌仍然继续生长,来获取体形更大,能量更高的食物,以获得充足的营养。
在本实验中,尖吻鲈的眼径在16日龄由等速生长转化为慢速生长,眼径的生长拐点与鲈鲤(Percocyprispingi pingi)14~15日龄相似[34],比条石鲷(Oplegnathus fasciatus)20日龄[35]和青龙斑[Epinephelus coioides(♀)×E.lanceolatus(♂)]20~21日龄晚[36]。本研究中尖吻鲈眼径的生长速率要小于口径和吻长,在稚鱼阶段转化为慢速生长,表明在尖吻鲈早期发育过程中口径和吻长的重要性要高于眼睛,可能是由其生长环境和生殖方式决定。
鱼类头高决定头部容积,头部容积关系到内部器官的发育,头部容积还可能限制仔、稚鱼口咽部、鳃的发育,进一步影响仔鱼的呼吸和消化系统的功能,而这些功能对仔、稚鱼的生长发育是至关重要的[37]。在16日龄以前头高与全长呈正异速生长,至拐点处,口咽腔和鳃等器官的发育已较为完善,头高由优先发育进入次要发育阶段,开始呈现负异速生长。许氏平鲉(Sebastes schlegelii)的早期发育也得到相似的结果[38]。
3.2 躯干部分发育
早期发育过程中,鱼类躯干部位的形态建成主要包括骨节、肌节结构的分化生长以及消化系统的发育[38]。仔鱼开口后,消化道后半部分弯曲形成早期的肠,之后肝脏、肾等消化器官的功能随着体长的增长而不断完善。而体高及肛前长相对体长全长而言为等速生长,表现为内部消化系统的完善是一个相对缓慢的过程。分析认为尖吻鲈仔、稚鱼骨节、肌节的分化及进一步发育可能在其早期发育过程中占次要地位,而肛前长并未表现出快速生长,可能是由于便于减少头部与尾部的距离,控制鱼体的长度,保证其在早期发育阶段身体保持平衡,使运动更加协调。在尖吻鲈仔、稚鱼阶段头长均为快速生长,在优先发育头部器官的过程中提供了空间基础,是发挥最佳摄食功能的选择,是鱼类在长期的发育进程中适应自然选择的结果[5]。
3.3 游泳器官的发育
早期尖吻鲈的运动器官主要包括鳍褶、尾鳍、胸鳍。胸鳍在16日龄出现生长拐点,胸鳍在仔鱼期保持等速生长,保持身体增长所需的同步运动能力,胸鳍主司平衡作用,稚鱼期它的快速生长为尖吻鲈平衡能力的提高奠定了基础,并与快速生长的背鳍、臀鳍、腹鳍一起协同作用,使稚鱼期的尖吻鲈平衡、游泳能力增强,进一步使其躲避敌害以及在不适环境下、增强主动摄食的能力[39]。背鳍、臀鳍、腹鳍在开口后16日龄以较全长更快的生长速率快速发育,充分说明了鳍条在尖吻鲈早期发育过程中的重要作用。此时的游动能力提高,稚鱼能以不同的游动方式游动。尖吻鲈仔鱼的尾鳍在16日龄前为慢速生长,之后转变为等速生长,这与红鳍笛鲷[6]、美洲鲥[4]、赤眼鳟(Spualiobarbus curriculus)[40]等研究结果不同,分析认为尖吻鲈初孵尾鳍的发育已较为完善,鳍褶作为辅助,代谢损耗降到最低,使短时间内更多的能量用来供给头部及身体部分比如胸鳍的发育,从而大大提高仔鱼的游泳及平衡能力。随着鳍褶的退化,各鳍条功能逐渐完善,各鳍条的相互协调提高了尖吻鲈的游泳能力,使其适应复杂多变的外界环境。
本文对尖吻鲈仔、稚鱼阶段的异速生长发育规律进行了较为系统的研究,探讨了其外部形态特征的发展及其重要的生态学意义。通过对尖吻鲈仔、稚鱼的发育研究可知,在尖吻鲈的早期发育过程中,仔、稚鱼的摄食、呼吸、游泳、感觉等大多数器官均表现出异速生长的特点。这些重要器官在仔、稚鱼早期快速的发育,使孵化后仔、稚鱼在早期生长阶段中,在最短时间内实现器官功能与环境的协调,从而使其获得早期生存相关的各种能力,使仔鱼主动摄食、躲避敌害的能力增强,这也是长期自然选择过程中的生态学适应[7]。因此,根据尖吻鲈早期发育的特点,在人工培育苗种生产中,应尽可能克服尖吻鲈早期发育的几个瓶颈[26](开口及驯化等),根据尖吻鲈自身的生长规律,创造鱼苗所需的最佳环境条件,投喂适口性较好的开口饵料,从而有效获取外源营养而降低死亡率,在尖吻鲈自然繁育的季节,对其产卵场进行保护,创造良好的产卵和育苗环境,对推动尖吻鲈养殖业发展具有重要意义。
参考文献:
[1] PETRER RH.The ecological implications of body size(2nd ed)[M].Cambridge:Cambridge University Press,1986.
[2] NIKLASK J.Plant allometry:he scaling of form and process[M].Chicago:University of Chicago Press,1994.
[3] 马 境,章龙珍,庄 平,等.施氏鲟仔鱼发育及异速生长模型[J].应用生态学报,2007,18(12):2875-2882.MA J,ZHANGL Z,ZHUANGP.Development and allometric growth patterns of larval Acipenser schrenckii.[J].Chinese Journal of Applied Ecology,2007,18(12):2875-2882.
[4] 高小强,洪 磊,刘志峰,等.美洲鲥仔稚鱼异速生长模式研究[J].水生生物学报,2015,39(3):638-644.GAO X Q,HONG L,LIU Z F,et al.The study of allometric growth pattern of American shad larvae and juvenile(Alosa Sapidissima)[J].Acta Hydrobiologyca sinica,2015,39(3):638-644.
[5] 宋洪建,刘 伟,王继隆,等.大马哈鱼卵黄囊仔鱼异速生长及其生态学意义[J].水生生物学报,2013,37(2):329-335.SONG H J,LIU W,WANG J L,et al.Allometric growth during yolk-sac larvae of chum salmon(Oncorhynchus keta Walabaum)and consequent ecological significance[J].Acta Hydrobiological Sinica,2013,37(2):329-335.
[6] 程大川,马振华,江世贵,等.红鳍笛鲷仔、稚鱼异速生长[J].水生生物学报,2017,41(1):206-213.CHENG D C,MA Z H,JIANGSG,et al.Allometric growth in larval and juvenile crimson snapper Lutjanus Erythopterus[J].Acta Hydrobiologyca sinica,2017,41(1):206-213.
[7] KHEMIS I B,GISBER E,ALCARAZ C,et al.Allometric growth patterns and development in larvae and juveniles of thick-lipped grey mullet Chelon labrosus reared in mesocosm conditions[J].Aquaculture Research,2013,44(12):1872-1888.
[8] 闵宽洪,朱 健,张成锋,等.尼罗尖吻鲈与几种鱼肌肉营养成分的比较[J].扬州大学学报,2008,29(1):52-55.MIN K H,ZHU J,ZHANG C F,et al.Analysis and comparison on the nutritional components of Lates niloticus with other fish species[J].Journal of Yangzhou University,2008,29(1):52-55.
[9] 孙典荣,李 渊,王雪辉,等.海南岛近岸海域鱼类物种组成和多样性的季节变动[J].南方水产科学,2012,8(1):1-7.SUN D R,LI Y,WANG X H,et al.Seasonal changes of species composition and diversity of fishes in coastal waters of Hainan Island,China[J].South China Fisheries Science,2012,8(1):1-7.
[10] 赵淑江,张晓举,李崇德,等.中国海水养殖鱼类中的外来物种 [J].海洋科学,2006,30(10):75-80.ZHAO SJ,ZHANG X J,LI C D,et al.Alien fishes of mariculture in China[J].Marine Sciences,2006,30(10):75-80.
[11] 陈忠康.尖吻鲈人工繁殖技术现状及其发展前景[J].现代渔业信息,1991,6(12):21-23.CHEN Z K.The status and prospect of artificial propagation technique of Lates calcarifer[J].Mordern Fisheries Information,1991,6(12):21-23.
[12] 莫介化,张邦杰,林加敬,等.人工海水培育尖吻鲈稚幼鱼的生长特性[J].现代渔业信息,2002,17(2):21-24.MO J H,ZHANG B J,LIN J J,et al.The growth feature in larval and juvenile of Lates calcarifer cultivated in artificial sea water[J].Mordern Fisheries Information,2002,17(2):21-24.
[13] NEWTON J R,SANTIS C D E,JERRY D R.The gene expression response of the catadromous perciform barramundi Latescalcarifer to an acute heat stress[J].Journal of Fish Biology,2012,(81):81-93.
[14] ROBEIRO F F,QIN J G.Pray size selection and cannibalistic behavior of juvenile barramundi Lates calcarifer[J].Journal of Fish Biology,2015,(86):1549-1566.
[15] CAREY G R,KRAFT P G,CRAMAP R L,et al.Effect of incubation temperature on muscle growth of barramundi Lates calcarifer at hatch and postexogeous feeding[J].Journal of Fish Biology,2009,(74):77-89.
[16] JERRY D R,SMITH K C.The genetics of Asian sea bass Lates calcarifer[M].Florida:CRC Press,2013.
[17] 庄 平,宋 超,章龙珍,等.全人工繁殖西伯利亚鲟仔稚鱼发育的异速生长[J].生态学杂志,2009,28(4):681-687.ZHUANG P,SONG C,ZHANG L Z,et al.Allometric growth of artificial bred Siberian sturgeon Acipenser baeri larvae and juveniles[J].Chinese Journal of Ecology,2009,28(4):681-687.
[18] STROBAND H W J,KROON A G.The development of the stomach in Clarias lazera and the intestinal absorption of protein macromolecules[J].Cell and Tissue Research,1981,215:397-415.
[19] 殷名称.鱼类早期生活史研究与进展[J].水产学报,1991,15(4):349-355.YIN M C.Advances and studies on early life history of fish[J].Journal of Fisheries of China,1991,15(4):349-355.
[20] IGUCHI K,MIZUNON.Early starvation limits survival in amphidromous fishes[J].Journal of Fish Biology,1999,54(1):705-712.
[21] KAMLER E.Early life history of fish:An energetics approach[M].London:Chapman&Hall,1992.
[22] KOUMOUMOUNDOUROSG,DIVANACH P,KENTOURI M.Osteological development of the vertebral column and of the caudal complex in Dentex dentex[J].Journal of Fish Biology,1999(54):424-436.
[23] 郑 珂,岳 昊,郑攀龙,等.海水养殖鱼类仔、稚鱼骨骼发育与畸形发生[J].中国水产科学,2016,23(1):250-261.ZHENG K,YUE H,ZHENG P L,et al.Skeletal ontogeny and deformities in commercially cultured marine fish larvae[J].Journal of Fishery Sciences of China,2016,23(1):250-261.
[24] 黄建盛,陈 刚,张建东,等.温度、pH及体重对低盐水体下斜带石斑鱼幼鱼耗氧率与排氨率的影响[J].动物学杂志,2016,51(6):1038-1048.HUANG J S,CHEN G,ZHANG JD,et al.Effects of temperature,pH and body wet weigh on oxygen consumption rate and ammonia excretion rate of orange-spotted grouper Epinephelus coioides juveniles cultured in low-salt wate[J].Chinese Journal of Zoology,2016,51(6):1038-1048.
[25] 区又君,李加儿,艾 丽,等.广东池塘培育条石鲷仔、稚、幼鱼的早期发育和生长[J].南方水产科学,2014,6(10):66-71.OU Y J,LI J E,AI L,et al.Early development and growth of larval,juvenile and young Oplegnathus fasciatus reared in pond in Guangdong Province[J].South China Fisheries Science,2014,6(10):66-71.
[26] GLUCKMANN I,HURIAUX F,FOCANT B,et al.Postembryonic development of the cephalic skeleton in Docentrachus labrax (Pisces, Perciformes,Serranidae)[J].Bulletin of Marine Science,1999,65(65):11-36.
[27] OLLA B L,DAVISM W,RYER C H.Behavioural responses of larval and juvenile walleye pollock(Theragra chalcogramma):Possible mechanisms controlling distribution and recruitment[J].ICES Marine Science Symposia,1995(201):3-15.
[28] 代田昭彦.水产饵料生物学[M].刘世英,雍文岳,译.北京:农业出版社,1985:149-177.DAI T Z Y.Biological science of aquatic feed[M].LIU SY,YONG W Y,translated.Beijing:Agicultural Press,1985:149-177.
[29] 区又君,李加儿,丁彦文.人工培育条件下真鲷仔稚鱼的生物学特性[C]∥全国首届青年水产学术研讨会论文集.上海:同济大学出版社,1995:315-324.OU Y J,LI J E,DING Y W.Biological features of the larval and juvenile of snapper sea bream cultivated in artificial conditions[C]//The first national youth symposium proceedings.Shanghai:Tongji University Press,1995:315-324.
[30] 区又君,李加儿.人工培育条件下鲻鱼早期发育的生理生态研究[J].热带海洋,1998,17(4):29-39.OU Y J,LI J E.The ecophysiological studies of the early development of Mugil cephalus under artificial conditions[J].Journal of Tropical Oceans,1998,17(4):29-39.
[31] OSSE J W M,BOOGAART J G M,SINK G M J.Priorities during early growth of fish larvae[J].Aquaculture,1997,155(1-4):249-258.
[32] GISBER B E,DOROSHOV SI.Allometric growth in green sturgeon larvae[J].Journal of Applied Ichthyology,2006,22(S1):202-207.
[33] PENA R,DUMAS S.Development and allometric growth patterns during early larval stages of the spotted sand bass Paralabrax maculatofasciatus(Pereoidei:Serranidea)[J].Scientia Marina,2009,73(S1):183-189.
[34] 何勇凤,吴兴兵,朱永久,等.鲈鲤仔鱼的异速生长模式[J].动物学杂志,2013,48(1):8-15.HE Y F,WU X B,ZHU Y J,et al.Allometric growth pattern of Percocypris pingi pingi larvae[J].Chinese Journal of Zoology,2013,48(1):8-15.
[35] 何 滔,肖志忠,刘清华,等.条石鲷早期发育阶段的生长模式[J].水产学报,2012,36(8):1242-1248.HE T,XIAO Z Z,LIU Q H,et al.Allometric growth in rock bream larvae(Oplegnathus fasciatus Temminck et Schlegel 1844)[J].Journal of Fisheries of China,2012,36(8):1242-1248.
[36] 吴水清,李加儿,区又君,等.斜带石斑鱼(♀)×鞍带石斑鱼(♂)杂交子代仔、稚鱼的异速生长[J].中国水产科学,2014,21(3):503-510.WU SQ,LI J E,OU Y J,et al.Allometric growth of hybrid grouper(Epinephelus coioides♀ ×E.lanceolatus♂)larvae and juveniles[J].Journal of Fishery Sciences of China,2014,21(3):503-510.
[37] SINK G M J,BOOGAART J G M,OSSE J W M.Larval growth patterns in Cyprinus carpio and Clarias gariepinus with attention to the finfold[J].Journal of Fish Biology,1997,50(6):1339-1352.
[38] 席 丹,张秀梅,吕红建,等.许氏平鲉早期异速生长模式的研究[J].中国海洋大学学报,2014,44(12):28-34.XI D,ZHANG X M,LV H J,et al.Studies on the early allometric growth pattern of black rockfish Sebastes schlegelii[J].Journal of Ocean University of China,2014,44(12):28-34.
[39] 于晓明,张秀梅.鱼类游泳能力测定方法的研究进展[J].南方水产科学,2011,8(4):76-84.YU X M,ZHANG X M.Research progress on measurement of fish swimming ability[J].South China Fisheries Science,2011,8(4):76-84.
[40] 陈方灿,李新辉,李 捷,等.珠江肇庆江段赤眼鳟开口后仔、稚鱼的异速生长分析[J].广东农业科学,2015(3):103-109.CHEN F C,LI X H,LI J,et al.Allometric growth of Spualiobarbus curriculus larvae and juveniles in Zhaoqing Reach of Pearl River[J].Guangdong Agricultural Sciences,2015(3):103-109.
