不同计算方式表征三峡水库临界层深度的合理性
2018-05-02
(三峡大学 水利与环境学院,湖北 宜昌 443002)
Sverdrup[1]于1953年建立了海洋春季水华暴发的临界层理论,认为水体分层下的水体混合层深度小于临界层深度是海洋春季水华暴发的根本原因;该理论也被较好地用于解释淡水湖泊水华的季节性暴发和演替的规律[2-5];对于河道型水库香溪河,其水华爆发频繁,很多学者也透过临界层理论对其水华爆发的原因做出了各自的解释[6-8]。
临界层理论能否较好地运用于水库,需要对水体临界层深度做出合理界定。多数研究采用真光层深度代替临界层深度,一些学者用光照强度为表面光强1%对应的深度(用Z1%表示)来代替临界层深度[6,9-12],也有学者将水下光强为14μmol/(m2·s) 对应的深度(用Z14表示)称为临界层深度[13],还有学者直接用2.7SD(用Z2.7SD表示)来表征临界层深度[7,8,14]。 上述计算方法均是在海洋、湖泊中得到合理运用后被相关学者直接套用于三峡水库,关于上述方法是否适用于河道型水库的研究却很少见。
本文利用黑白瓶测氧法,通过测定溶解氧的变化计算每天的临界层深度(Zcr),再与通过光照计算出的Z1%,Z14,Z2.7SD进行比较,试分析上述计算方式套用于香溪河库湾的合理性及原因,可为临界层理论运用于三峡水库提供借鉴意义。
1 材料与方法
根据香溪河自然环境特点及实验实施的可行性,选取香溪河水华多发河段高岚河河段为观测站点进行实验。
1.1 黑白瓶法基本原理
通过测定水中溶解氧的变化,间接计算有机物质的生成量,是用黑白瓶测氧法研究初级生产力的基本原理。当带有浮游生物样品的黑白瓶悬挂水中曝光时,黑瓶中藻类由于得不到光照,只能进行呼吸作用,黑瓶中的溶解氧将会减少;与此同时,白瓶中的藻类在光照条件下,光合作用与呼吸作用同时进行,白瓶中的溶解氧量一般会明显增加,假定光照条件与黑暗条件下的呼吸强度相等,就可根据挂瓶曝光期间内,黑、白瓶中的溶解氧变化计算出光合作用与呼吸作用的强度[15]。
1.2 实验材料和试剂
采样瓶:250mlBOD瓶。每次取样前BOD瓶用稀盐酸洗涤后再用蒸馏水洗净。
挂瓶器材:分层采水器,长尼龙绳,木夹子,黑布袋
测量仪器:多参仪(hydrolabDS5,美国HACH公司);ZDS-10W-1D水下光量子仪 (测量波长范围:400~760nm);萨氏盘(Secchi Disc)。
实验材料:测定溶解氧的实验仪器和药剂。
1.3 测定步骤
1.3.1 水样采集
水体中的藻类主要在表层至8m水深之间活动[16],故本实验分层按0,0.5,1,2,3,4,5,6,7,8m分, 共分10层。每层取水250ml的BOD瓶4瓶。第1瓶用黑布袋套好挂回原水层,用来测定黑瓶的溶解氧;第2瓶直接挂回原水层,作为白瓶测定溶解氧;第3瓶作为初始值直接测量溶解氧;第四瓶用于藻类鉴定。挂完水样后用多参仪测定水温;用光量子仪测定水下光照;用萨氏盘测定透明度。同时记录气候、环境指标。灌水时将导管插到BOD瓶底部,灌满瓶后还要溢出2~3倍体积的水,保证所有样品瓶中的溶解氧与所采水样的溶解氧完全一致。曝光24h后,取回所挂的2瓶水样并挂上新的水样,重复以上工作,共持续10d。
1.3.2 黑白瓶的固定
曝光结束后,取出黑白瓶,立即加入1ml硫酸锰溶液和2ml碱性碘化钾溶液对瓶中溶氧进行固定,固定摇匀后放入黑暗处带回室进行滴定测氧。若瓶中有大量气泡,加药剂时应微微倾斜BOD瓶避免氧气逸出。
1.4 计算方法
1.4.1 各水层日生产量(O2,mg/L)计算
总生产量=白瓶溶解氧量-黑瓶溶解氧量。
净生产量=白瓶溶解氧量-初始瓶溶解氧量。
呼吸作用量=初始瓶溶解氧量-黑瓶溶解氧量。
1.4.2 临界层深度计算
按上面方法计算出各水层日净生产量。根据测定的各水层净生产量与对应水深拟合函数关系:
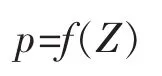
式中 p为水层日净生产量;Z为水深。
当p=0时,对应的Zco为光补偿深度,即光合作用强度等于呼吸作用强度的水层深度[17],Zco以上的水层是有有机物积累的,Zco以下是消耗有机物的。将各天的函数式在深度上积分得水柱日净生产量P=0-zf(Z)dZ,当P=0时对应的Zcr为临界层深度,即整个水柱有产物积累或消耗有机物的水层分界深度[1]。
2 结果与分析
2.1 净生产力p的垂向分布
在为期10d的连续监测中,经历了2次降雨,这10d的气候情况如表1。

表1 气温、气候变化情况
实验结果表明,降雨对藻类生产力影响很大[18],雨水冲击和雨天微弱的阳光使藻类生产力极大降低。2016年9月11日和9月17日两次降雨都使藻类生产量骤降,也改变了藻类的垂向分布规律。
各水层净产量如图1,净产量主要产生在0~2m的水层。2016年9月9日和9月16日为晴天,光照最强,这两天产量最高,分别为1.76mg/L和1.77mg/L。降雨期间产量很低,9月11日的降雨冲散了聚集的藻,使藻分散在各层水体中,随后连续的晴天又使藻类慢慢向水面聚集。9月13日~9月16日是雨后逐渐天晴的过程,也是藻类生产量逐步回升的过程。可以看出生产力的雨后恢复缓慢,而降雨对生产力的影响却是骤然的[18]。 2m以下净产量降低很快,在3~5m之间变为0,其后均为负值。说明这10d的光补偿深度都在3~5m,晴天补偿深度略大于阴雨天,这与晴天光照强度大,透射到水里的有效辐射更深有关。各天5m以下的产生情况相差不大。通过每天分层取样鉴定,发现在水层5m以下的水体中,藻类相对较少且分布均匀,呼吸作用大致相同,光合作用随水深递减。
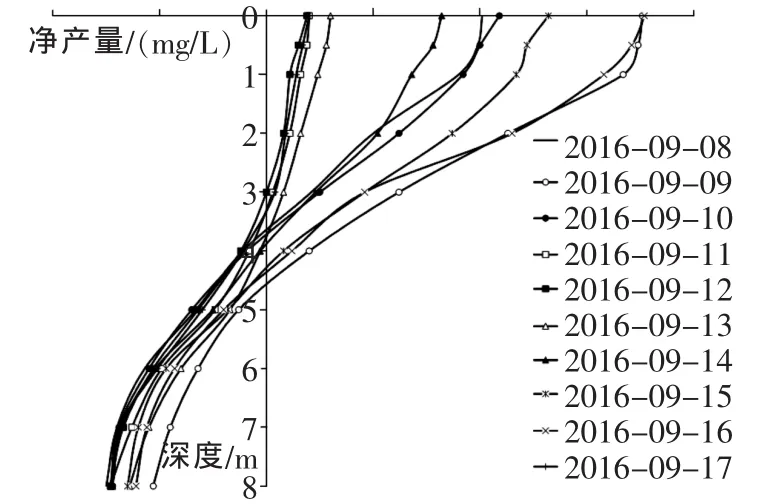
图1 每日净产量的垂向分布
2.2 光照强度IZ垂向分布与光衰减系数k
每日水下光分布情况如图2,每日表层光照强度差别较大,晴天最大值高达1111μmol/(m2·s),雨天最低低于46.21μmol/(m2·s)。
光照强度沿水深衰减过程均符合Sverdrup[1]在临界层理论中提出的指数分布公式:
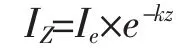
式中 IZ为水下Z深度的光照强度;Ie为水面光照强度;k为水下光衰减系数。
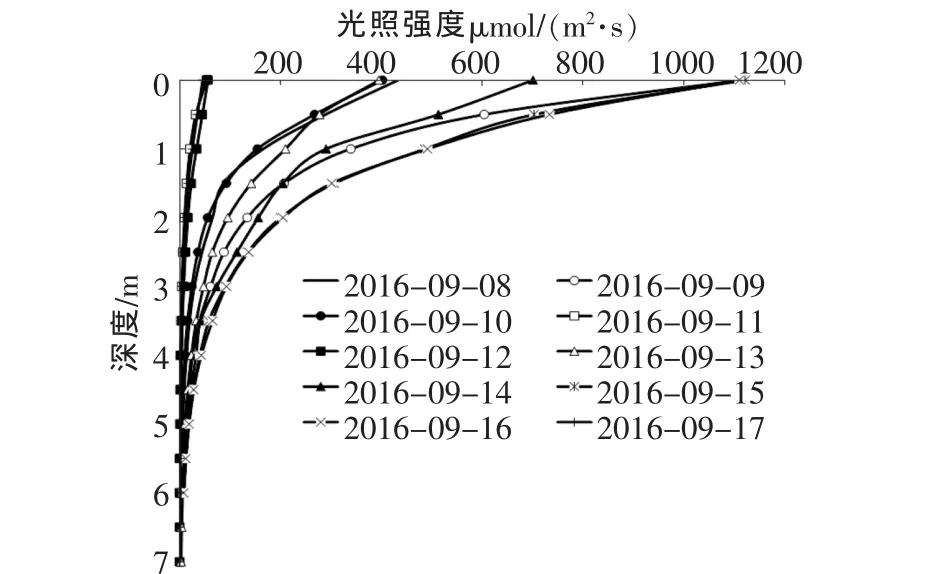
图2 每日水下光分布情况
由图2可知,在真光层内光照强度快速衰减,在水下7m左右接近0。真光层是根据水体透明度计算而来,而水体透明度与水中悬浮粒子、藻颗粒对光的散射和吸收直接相关[8]。所以光照强度在水下衰减,可以很好地反应水中藻颗粒的量及分布情况。
根据上述指数公式,对实验期间各天的光照进行指数函数拟合,求出各天的光衰减系数。2016-09-08~2016-09-17,两种光衰减系数的变化情况如图3,分别求出了2m以内的光衰减系数和整个真光层的光衰减系数。
图3结果表明,晴天2m以内的光衰减系数明显大于整个真光层的衰减系数,说明晴天光照强度在2m以内水层衰减的比下层快,这反映了晴天藻类聚集在水面的现象。随着天气变化,两种光衰减系数的大小关系也跟着变化,反应出藻颗粒在水中的分布情况随天气的变化。
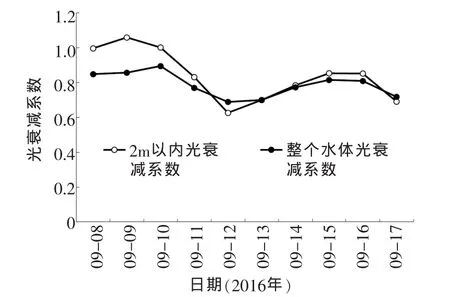
图3 两种光衰减系数的变化情况
2.3 临界层深度Zcr与光补偿深度Zco的变化规律
根据第1.4章节积分式求得各天的临界层深度Zcr和光补偿深度Zco,如图4。
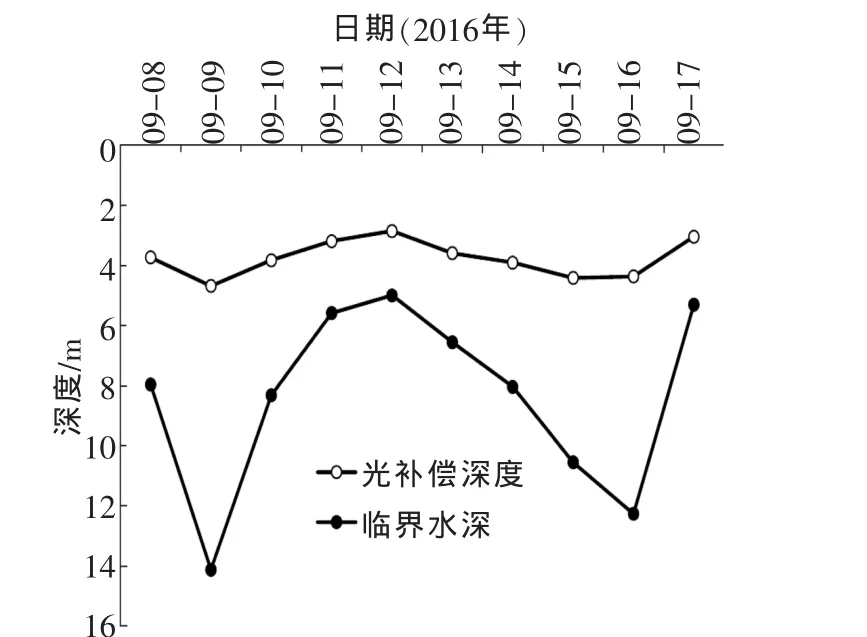
图4 Zcr和Zco变化趋势
由图4可知,Zcr波动很大,最大值高达14.12m,最小值低至5.00m。整体符合晴天深,雨天浅,降雨会使临界层深度骤然降低,而雨后天晴恢复的过程是缓慢的。Zco变化不大,均在4m左右,晴天略高于阴雨天。光补偿点只考虑单层水体的生产情况,与上、下水层产量的积累或消耗均无关系。Zco波动不大说明藻类在4m左右水层的光合作用与呼吸作用大致相同。Zcr与Zco变化趋势的不同,说明藻类初级生产力的差异主要产生在4m以上的水层。
2.4 按光照强度计算3种临界层深度(Z1%,Z14,Z2.7SD)
真光层深度指开放水体中阳光照射所达、光合作用得以发生的水体深度,为水柱中支持净初级生产力的部分,其底部为临界深度,海洋、湖泊、河流等水域生态系统中藻类基本分布在这一层[8]。很多学者在研究中把真光层深度等同于临界层深度。根据不同的计算方法即可确定不同的临界层深度,如图5,按3种不同的方式计算出临界层深度。
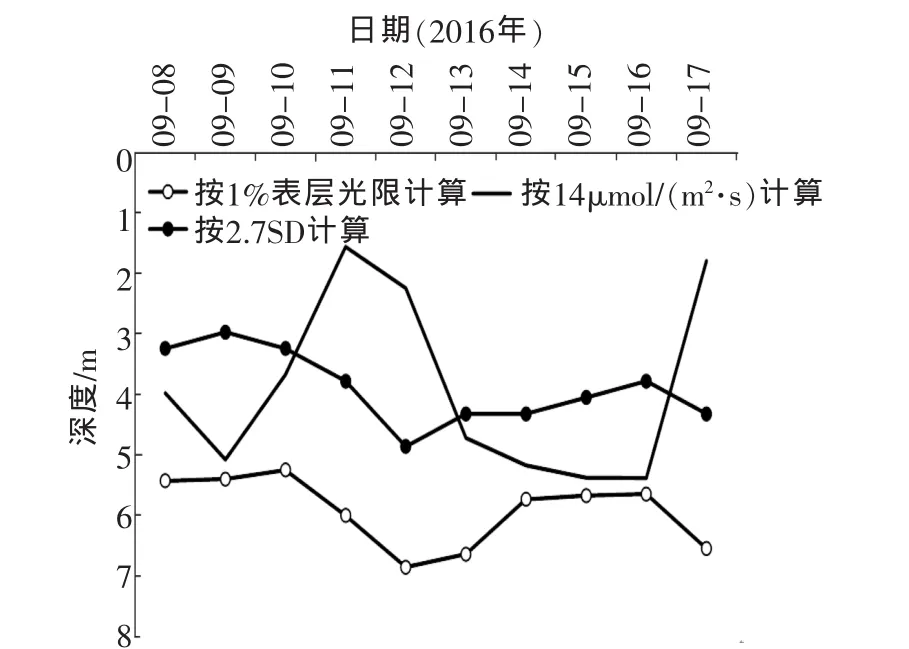
图5 不同计算方式下的临界层深度
由图5可以看出Z1%和Z2.7SD波动较小,二者的变化趋势相似,后者的深度整体小于前者。
(1)Z1%和Z2.7SD均受表层水体中藻细胞浓度影响较大,当藻类聚集在水面时会急剧降低,Z1%按百分比计算而来,与表层光照强度基数大小无关。Z1%和Z2.7SD均可反应监测时期的现存藻生物量大小及分布情况,但不能反应各水层藻细胞所受到的实际光照强度,因而无法根据Z1%和Z2.7SD直接推测这些藻细胞当日的光合作用产量。
(2)Z14波动较大,其波动规律同表层光照强度关系密切。Z14同时考虑到不同藻密度水体对光照强度的衰减作用和表层光照强度基数的大小,但由于各天表层光照强度差异远大于不同藻密度水体衰光作用的差异,故Z14受表层光照强度基数大小的影响更为突出。
3 相关关系分析
为探明实际临界层深度Zcr与真光层深度有何关系,本文对Zcr与Z1%,Z14,Z2.7SD及表层光照强度Ie做出了相关关系计算,结果如表2。
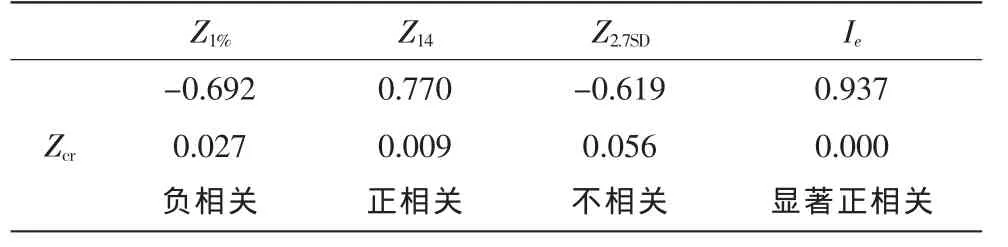
表2 相关关系统计
3.1 Zcr与Z1%存在相关性
相关性不显著(r=-0.692)。Z1%受现存藻密度影响较大,与表层光照强度基数大小无关。例如:9月17日表层光照为46.21μmol/(m2·s),其10%只有4.62μmol/(m2·s),计算到1%的水层没有生产意义,使Zcr与Z1%相关性不显著。同时,当藻类光合作用增强时,藻密度会变大,水体透光性会降低,Z1%会变小;随后,水体透光性降低又会反过来迫使藻类光合作用降低,从而使水体透光性又升高,Z1%又变大,因此藻类光合作用与Z1%存在一个负反馈调节过程。以至于在调节过程中的不同阶段测量,Zcr与Z1%可能会出现正、负不同的相关性。
3.2 Zcr与Z14的相关性略大于与Z1%的相关性
其原因是Z14中考虑到表层光照强度的大小,避免了Z1%中因Ie×1%过大或过小造成的误差,但Zcr与Z14的相关程度(r=0.770)也不高,藻类生产力除了受光照强度影响大,受藻生物量大小及藻颗粒在水中分布情况的影响也大[20]。在相同环境下,大部分藻聚集在表层水体的生产力会比聚集在14μmol/(m2·s)水层时的生产力大,因Z14无法考虑到藻颗粒数量和分层情况,故不能很好地描述实际临界层深度。
3.3 Zcr与Z2.7SD无相关性
Z2.7SD根据光密度得来,测量时局限在水面很浅的一层,在淡水湖泊中可以近似代表整个水深的透光情况,但是河道型水库一般比淡水湖泊深很多,表层1~2m无法代表整个水深的透光情况。同时,光密度值是通过人眼观察所得,不同的人眼力不同,相同的人在不同环境下读数也会有所差异。
4 讨论
通过上述分析,淡水湖泊和海洋中运用较好的临界层理论并不能简单地套用在三峡水库,河道型水库应具有适合其水环境条件的表示方法。
河道型水库相比于湖泊有以下特点:河水深度比湖水深度深很多,河水一般比湖水透明度高;河库中的藻生物量一般比湖泊中少,但垂向迁移能力更强,迁移程度更大;河库中水流速度较大,受三峡大坝调洪影响,库区水位波动也大。
三峡水库和普通湖泊主要的不同表现在水深和藻类迁移特点上,对于不同的季节和储、泄水工况,其计算方式应当存在差异。本实验在夏末秋初储水阶段进行,从相关性分析中发现表层光照强度与临界层深度有较高的相关性(r=0.973)。为探明Zcr与Ie的相关关系,拟合函数图像如图6。
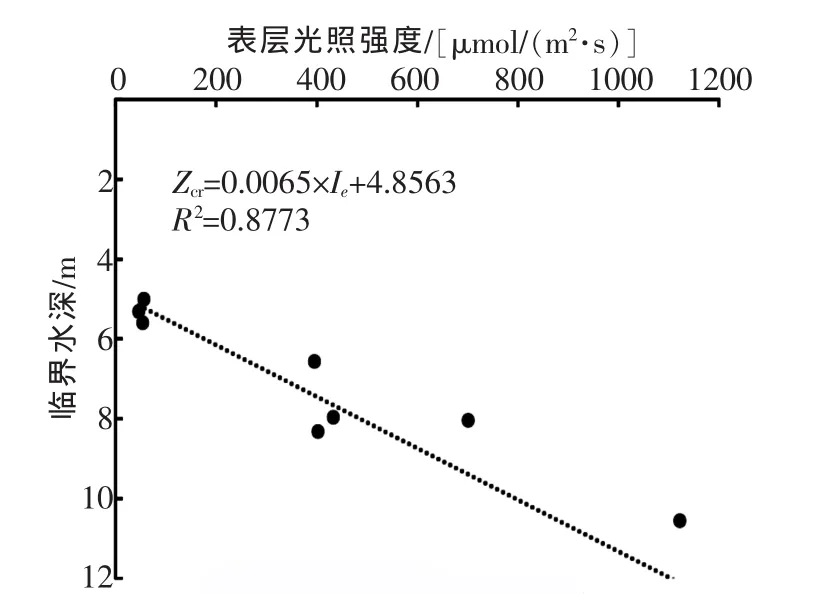
图6 Zcr和Ie的关系曲线
由于没有考虑到现存生物量的影响,函数式Zcr=0.0065×Ie+4.8563只适用于这个季节香溪河特定的地方,对于拥有不同藻生物量的地点会有不同的关系式。笔者看来,仅仅考虑光照是不能推求出通用公式的,应结合藻生物量和其分层情况对临界层深度做进一步研究,推求真正适合三峡库区的计算方法。
5 结语
(1)利用光照计算出的真光层深度代替临界层深度在实验方法上具有简单、易操作性,但光照是无法完全代表初级生产力的。在河道型水库中应当结合藻生物量对其计算方法进行改进。
(2)由于河道型水库一般较深,Z2.7SD不再具有近似代表整个水深透光性的作用,固不能直接运用于三峡水库。
(3)Z1%和Z14与Zcr具有一定的相关性,但均需要根据实际情况进行改进。相比于Z1%,Z14更能适应香溪河多变的天气和光照强度,在没有更好的计算方法之前,Z14应是学者研究香溪河水库首选的计算方法。
参考文献:
[1]Sverdrup H U.On conditions for the vernal blooming of phytoplankton[J].ICES Journal of Marine Science,1953,18(3): 287-295.
[2]Berger S A,Diehl S,Stibor H,et al.Water temperature and mixing depth affect timing and magnitude of events during spring succession of the plankton [J].Oecologia,2007,150(4): 643-654.
[3]Fonseca B M,Bicudo C E M.Phytoplankton seasonal variation in a shallow stratified eutrophic reservoir (Gar as Pond,Brazil)[J].Hydrobiologia,2008,600(1): 267-282.
[4]Ganf G G,Oliver R L.Vertical Separation of Light and Available Nutrients as a Factor Causing Replacement of Green Algae by Blue-Green Algae in the Plankton of a Stratified Lake[J].Journal of Ecology,1982,70(3):829-844.
[5]Lindenschmidt K E, Chorus I.The effect of water column mixingonphytoplanktonsuccession,diversityandsimilarity[J].Journal of Plankton Research,1998,20(10):1927-1951.
[6]刘流,刘德富,肖尚斌,等.水温分层对三峡水库香溪河库湾春季水华的影响[J].环境科学,2012,33(9):3046-3050.
[7]姚绪姣,刘德富,杨正健,等.三峡水库香溪河库湾冬季甲藻水华生消机理初探 [J].环境科学研究,2012,25(6):645-651.
[8]陈洋,杨正健,黄钰铃,等.混合层深度对藻类生长的影响研究[J].环境科学,2013,34(8): 3049-3056.
[9]Kirk J T O.Light and photosynthesis in aquatic ecosystems[M].Cambridge: Cambridge University Press,1994.
[10]Khanna D R,Bhutiani R,Chandra K S.Effect of the euphotic depth and mixingdepth on phytoplanktonic growth mechanism [J].Internetional Journal of Environmental Research,2009,3(2): 223-228.
[11]Sakshaug E,Bricaud A,Dandonneau Y,et al.Parameters of photosynthesis: definitions,theory and interpretation of results [J].Journal of Plankton Research,1997,19(11):1637-1670.
[12]刘流,刘德富,黄钰铃,等.香溪河库湾春季水华纵向分布对水层结构的响应[J].三峡大学学报,2012,34(2):1-6.
[13]Reinart A, Arst H, Erm A, et al.Optical and biological properties of Lake lemiste,a water reservoir of the city of Tallinn II: Light climate in Lake lemiste [J].Lakes&Reservoirs Research&Management,2001,6(1):75-84.
[14]Lawrence I, Bormans M, Oliver R, et al.Physical and nutrient factors controlling algal succession and biomass in Burrinjuck Reservoir [R].Canberra: Land and Water Resources Australia,2000.
[15]赵道全,申秀英,周晓林,等.南湾水库初级生产力的初步研究[J].河南水产,2003(4): 31-32.
[16]杨正健,徐耀阳,纪道斌,等.香溪河库湾春季影响叶绿素a的环境因子[J].人民长江,2008,39(15): 33-35.
[17]合田健.水环境指标[M].北京:中国环境科学出版社,1989.
[18]纪道斌,李媛,孔松,等.典型暴雨洪水对三峡水库香溪河库湾水华的影响[J].中国农村水利水电,2013(6):39-44.
[19]杨东方,陈生涛,胡均,等.光照、水温和营养盐对浮游植物生长重要影响大小的顺序[J].海洋环境科学,2007,26(3):201-207.
[20]蔡琳琳,朱广伟,李向阳.太湖湖岸带浮游植物初级生产力特征及影响因素[J].生态学报,2013,33(22): 7250-7258.
