异质养分环境下邻株竞争对3个木荷种源干物质积累及氮磷效率的影响
2018-04-19姚甲宝周志春楚秀丽徐红兵童建设
姚甲宝,周志春,楚秀丽,徐红兵,童建设
1 中国林业科学研究院亚热带林业研究所浙江省林木育种技术研究重点实验室, 富阳 311400 2 中国林业科学研究院亚热带林业实验中心, 分宜 336600 3 浙江省淳安县富溪林场, 淳安 311701
氮和磷是植物生长发育所必需的两种大量元素,对植物生产力提高起着举足轻重的作用[1- 3]。然而,由于有机物分解、无机养分扩散和土壤生化过程等原因,土壤中的氮和磷通常呈高度的异质性分布[4- 5]。植物能够通过根系形态和生理塑性变化增加对异质分布土壤养分资源的获取[6- 7],进而影响林木生长发育、种间竞争以及群落生产力。
不同植物对异质养分环境的适应能力存在明显差异[8],并受邻株竞争影响[9- 10]。吴开贤等[11]研究指出,无竞争下异质氮处理促进玉米(Zeamaysspp)和马铃薯(Solanumtuberosum)的生长,竞争则改变了两作物对异质性氮的觅养行为。两作物间作提高了玉米的觅养精确性,使玉米生长加快,但抑制马铃薯的生长。Fransen和de Kroon[12]发现,同质养分环境下紫羊茅(Festucarubra)和黄花茅(Anthoxanthumodoratum)单独生长时的根系养分吸收能力基本相同,而异质养分环境下两种植物间竞争降低了紫羊茅根系养分吸收效率,但提高了黄花茅根系的养分吸收能力。已有研究的多局限于邻株竞争时植物种间根系形态或生理指标差异比较上[6- 7,12],较少涉及植物基因型觅养塑性差异与生长竞争优势关联研究。因此开展异质养分环境和邻株竞争对林木干物质量与养分效率影响的研究,对更好地揭示不同林木基因型生长竞争能力差异机制,发挥其潜在的生理功能,提高森林生产力等具有重要的理论和实践意义。
木荷(Schimasuperba)是我国亚热带地区重要的珍贵阔叶用材、生物防火和生态防护造林树种,具有速生、丰产、材质优异和适应性强等特点。本研究组自2001年来开展了我国木荷的多目标育种,发现其存在丰富的种源和家系变异,并初选出一批优良种源和家系,同时发现使用不同木荷种源营造的混交林、纯林生产力差异巨大[13- 15],这应与不同木荷种源竞争能力和氮磷效率有关。本研究选择3个不同生产力木荷种源,模拟垂直异质性森林土壤环境,开展木荷同种及与杉木异种邻株竞争的盆栽试验,分析不同种源木荷生物量分配和养分效率变化,探讨木荷种源获取和利用资源策略与其生长表现之间的关系,揭示木荷种源生长竞争能力差异机制,旨在为选育氮磷效率高和竞争能力强的木荷新品种提供科学理论依据。
1 材料和方法
1.1 试验材料与试验设计
试验所有植物材料为浙江南部(龙泉)、福建北部(建瓯)和江西南部(信丰)3个生产力差异较大的木荷种源,杉木种子来自浙江省龙泉市林科院第3代杉木无性系种子园。采用直径25cm、高度40cm的无纺布袋作为盆栽容器。盆栽基质取自浙江省淳安县富溪林场的贫瘠酸性红壤,其全氮和全磷含量分别为0.34g/kg和0.35g/kg,水解氮、速效钾和有效磷含量分别为45.30、40.90、1.08g/kg,pH值为5.12,有机质含量为6.11g/kg。构建不同养分环境所用控释肥委托山东金正大集团专门生产,控释氮肥(全氮含量为440g/kg)、控释磷肥(有效磷440g/kg)和控释钾肥(有效钾含量440g/kg)的肥效均为2a。
试验采用2种养分环境、3种栽植方式和3个种源三因素析因设计,其中养分环境构建方法为:利用风干过筛后贫瘠酸性红壤为基质,将盆栽容器分为4层,每层高度10cm,层间用塑料网隔开,每层加入控释肥构建同质或异质养分环境。基于文献[16]报道的森林土壤数据,构建同质养分环境时,上下4层均匀加入等量的控释肥(在每千克基质中加入控释氮肥124.31g、控释磷肥20.27g和控释钾肥134.31g,相当于有效N、P和K含量分别为100、10、100mg/kg);构建异质养分环境时,第1层基质均匀加入3倍同质养分含量的控释肥,第2层加入1倍同质养分含量控释肥,第3、4层均不添加控释肥,从而模拟土壤垂直空间养分分布的异质性,即表层养分丰富,向深层迅速降低的森林土壤养分含量自然分布状况。异质和同质养分环境加入的N、P和K养分总量相同。在两种养分环境下,分别设置木荷单植、同一种源木荷双株纯植、木荷与杉木两株混植3种栽植方式。
1.2 盆栽试验
试验地为浙江省淳安县富溪林场(119.13°E,29.48°N),其所处海拔为130m,年平均气温17.5℃,年降水量为1701.2mm。盆栽试验在林场钢构自控荫棚内进行,荫棚四周放置林场培育的普通容器苗,该试验各处理盆栽苗摆放在靠近中心位置。3个木荷种源及杉木种子于2015年1月10日播种,5月7—8日选用生长整齐一致的芽苗移栽。纯植或混植的两株芽苗分别栽植在盆中线的正中点两侧5cm处,单植处理则在每盆正中间栽植1株木荷。每试验处理20盆,即重复20次。苗木按正常管理直至采收。
1.3 试验收获与指标测定
试验于2015年12月收获,每处理随机选取6株生长正常的1年生植株量测其苗高和地径,然后小心将根系与土壤分开,洗净根系浮泥,带回实验室用去离子水清洗干净。将苗木分成根、茎和叶3部分,经105℃杀青30min,80℃烘干至恒量,然后测定根、茎、叶和全株的干物质量,并计算根冠比(根干物质量/地上部分干物质量)。采用凯氏定氮法和浓H2SO4-H2O2消煮-钼锑抗比色法分别测定每株苗木各器官的氮和磷含量[17]。
1.4 数据分析
本研究所采用的磷效率包括植物从土壤中吸收磷的效率(即磷吸收效率)和同化利用所吸收磷的效率(即磷利用效率)两方面[18],其计算公式为:磷吸收效率=各器官或全株磷吸收量(mg/株);磷利用效率=各器官或全株干物质量/各器官或全株磷吸收量(g/mg),氮吸收效率和利用效率计算方法同上。采用Excel 2007软件计算和整理所得试验数据,并进行图表绘制。各处理苗木的干物质量和氮、磷吸收利用效率等指标使用SPSS 19.0软件按种源、养分环境、栽植方式及其交互作用进行三因素、二因素及单因素方差分析,并用DUNCAN法进行多重比较,以检验木荷苗木各性状的差异显著性(P<0.05)。
2 结果与分析
2.1 邻株竞争对木荷种源苗木干物质积累和分配的影响
方差分析结果显示,种源、养分环境和栽植方式对木荷全株及根、茎、叶干物质量的影响均达到极显著或显著水平(表1)。比较分析发现,木荷福建建瓯种源干物质生产能力较强,其全株干物质量分别较浙江龙泉和江西信丰种源高134.6%和61.3%。与同质养分环境相比,异质养分环境下3个木荷种源的全株干物质量均有所增加,其中福建建瓯种源增幅最大。木荷干物质量还存在显著种源×栽植方式互作效应,在异质养分环境下,福建建瓯种源在混植栽培时,其全株干物质量最高,分别较单植和纯植高26.2%,和91.5%。氮纯植时受到种内竞争的影响,其全株干物质量减小了34.0%。与之不同,混植时,江西信丰种源生长表现较差,全株干物质量显著降低,浙江龙泉种源受影响较小,而同种邻株竞争则有利于浙江龙泉种源的干物质积累,但对江西信丰种源影响不明显。相比之下,在同质养分环境中,江西信丰种源在邻株存在时均具有较高的干物质量,福建建瓯和浙江龙泉种源干物质量变化规律与异质养分环境相似。3个种源的根、茎和叶干物质量也有类似变化规律(表2)。
表1木荷种源干物质积累、分配及氮、磷效率的方差分析(F值)
Table1ANOVAanalysisondrymatteraccumulation,allocationandnitrogenandphosphorusefficiencyofS.superbaprovenances(Fvalue)
种源、养分环境、栽植方式、种源×养分环境、种源×栽植方式、养分环境×栽植方式和种源×养分环境×栽植方式的自由度分别为1、2、2、2、4、2和4;*P<0.05;**P<0.01。WDM:全株干物质量Whole plant dry mass;RDM:根干物质量Root dry mass;SDM:茎干物质量Stem dry mass;LDM:叶干物质量Leaf dry mass;RSR:根冠比Root:shoot ratio;PAE:全株磷吸收效率Phosphorus absorption efficiency;PUE:全株磷利用效率Phosphorus utilization efficiency;NAE:全株氮吸收效率Nitrogen absorption efficiency;NUE:全株氮利用效率Nitrogen utilization efficiency
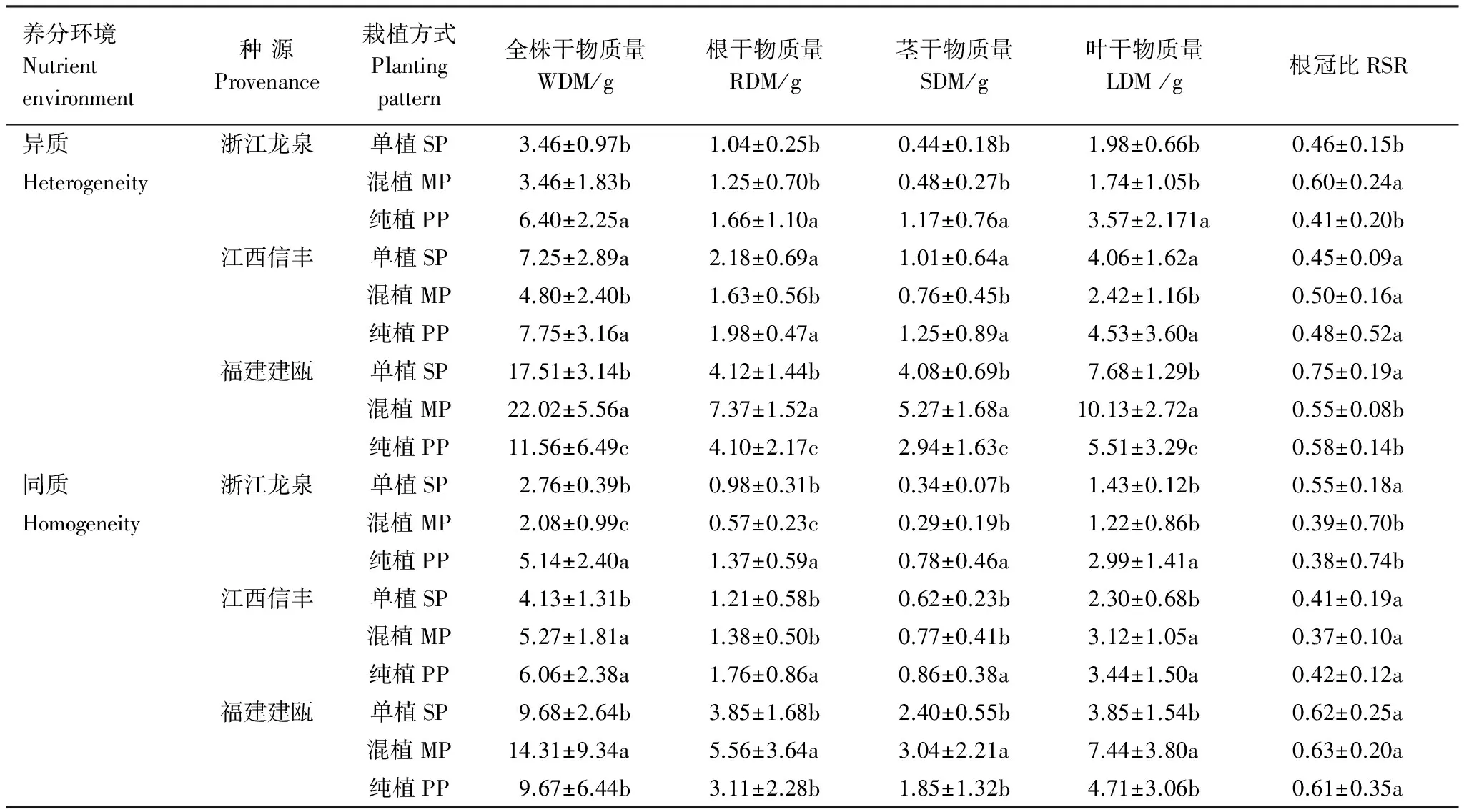
表2 栽植方式对3个木荷种源的干物质积累及其分配影响(平均值±标准差,n=6)
SP:单植 Single-planted; MP:双株混植 Mixed-planted ; PP双株纯植Pure-planted;不同小写字母表示同一种源不同栽植方式间差异显著(P<0.05)
根冠比用以反映植株生物量在地下和地上部分的分配比例。研究结果显示,两种养分环境中,福建建瓯种源根干物质量和根冠比均显著地大于其他种源,说明其根系较为发达。在异质养分环境中,干物质量较大的福建建瓯种源其根冠比在混植时显著降低,表明干物质较多地分配至地上部分。而在纯植时其根冠比较单植处理也明显下降,结合其各器官生物量均下降的结果,说明同种竞争对建瓯种源根部负向影响比地上部分更大。浙江龙泉种源根冠比仅在混植时显著提高,干物质较多地分配到地下部分,结果影响了地上部分的生长。与以上2个种源不同,在不同养分环境中邻株竞争存对江西信丰种源的根冠比影响较小,说明其干物质分配调节能力较低(表2)。
2.2 邻株竞争对木荷种源的氮吸收和利用效率的影响
养分环境和邻株竞争不仅影响了木荷幼苗的干物质积累,而且显著影响了木荷幼苗对氮素的吸收,且种源间差异显著(表1)。异质养分环境中3个木荷种源的氮吸收效率明显高于同质养分环境。比较不同种源氮吸收效率可知,干物质生产能力较强的福建建瓯种源,其全株和根、茎、叶的氮吸收效率也具有显著优势(图1)。在异质养分环境中,不同种源全株和叶氮吸收效率对邻株竞争的响应不同,存在显著的种源×栽植方式互作效应。混植栽培有利于福建建瓯种源对氮素的吸收,其全株氮吸收效率分别较单植和纯植高56.8%和82.6%。与之相反,混植对江西信丰种源负向影响较大,全株氮吸收效率下降约40%,但浙江龙泉种源受影响较小。同种邻株竞争显著增强了浙江龙泉种源的氮吸收能力,全株氮吸收效率较单植增长了74.9%,福建建瓯和江西信丰种源变幅不大,3个种源根、茎和叶的氮吸收效率也有相似变化趋势。
除叶片氮利用效率外,木荷全株和各器官的氮利用效率种源间及栽植方式间差异均未达到显著水平,这表明各种源间的生长差异主要与其氮吸收效率高低有关。在同质养分环境下,3种源的氮效率变化规律与异质养分环境类似。
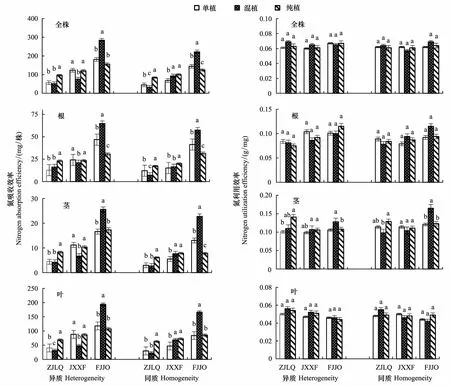
图1 栽植方式对3个木荷种源的氮吸收效率与利用效率的影响Fig.1 Effect of planting patterns on nitrogen absorption and utilization efficiency of three provenances of S. superba ZJLQ:浙江龙泉种源Longquan of Zhejiang provenance; JXXF:江西信丰种源Xinfeng of Jiangxi provenance; FJJO: 福建建瓯种源Jian′ou of Fujian provenance;不同小写字母表示同一种源不同栽植方式间差异显著(P<0.05);图中数据为平均值±标准差(n=6)
2.3 邻株竞争对木荷种源的磷吸收和利用效率的影响
由表1和图2可以看出,3个木荷种源在异质养分环境中均具有较高的磷吸收和利用效率。木荷磷效率的种源效应显著,干物质量最大的福建建瓯种源全株磷吸收效率成倍高于其他种源。而其全株磷利用效率也明显高于浙江龙泉种源,但与江西信丰种源之间无明显差异。不同种源磷效率还与邻株竞争方式有关,在异质养分环境中,与杉木混植时,福建建瓯种源的全株磷吸收效率增加明显,分别较单植和纯植增长了118.2%和106.4%,各器官的磷吸收效率变化趋势与之相似。同时各器官的磷利用效率也有不同程度的提高,尤其茎增加明显,说明其茎的物质转运能力明显增强。与之不同,江西信丰种源在混植时各磷吸收效率指标却显著降低了24.6%—60.8%,而浙江龙泉种源磷利用效率显著高于单植。纯植时,浙江龙泉和江西信丰种源表现为磷吸收能力增强,磷利用效率下降的背向趋势,而同种邻株竞争对福建建瓯种源的磷吸收和利用效率影响均不明显。在同质养分环境中,3个种源磷效率受栽植方式及其互作效应的影响显著,呈现磷吸收与利用效率相反的变化规律,如福建建种源在混植时磷吸收效率最大,但其磷利用效率却不高,而纯植时其磷吸收效率较低,但利用效率却较高,表明福建建瓯种源能运用不同生理策略应对邻株竞争(图2)。
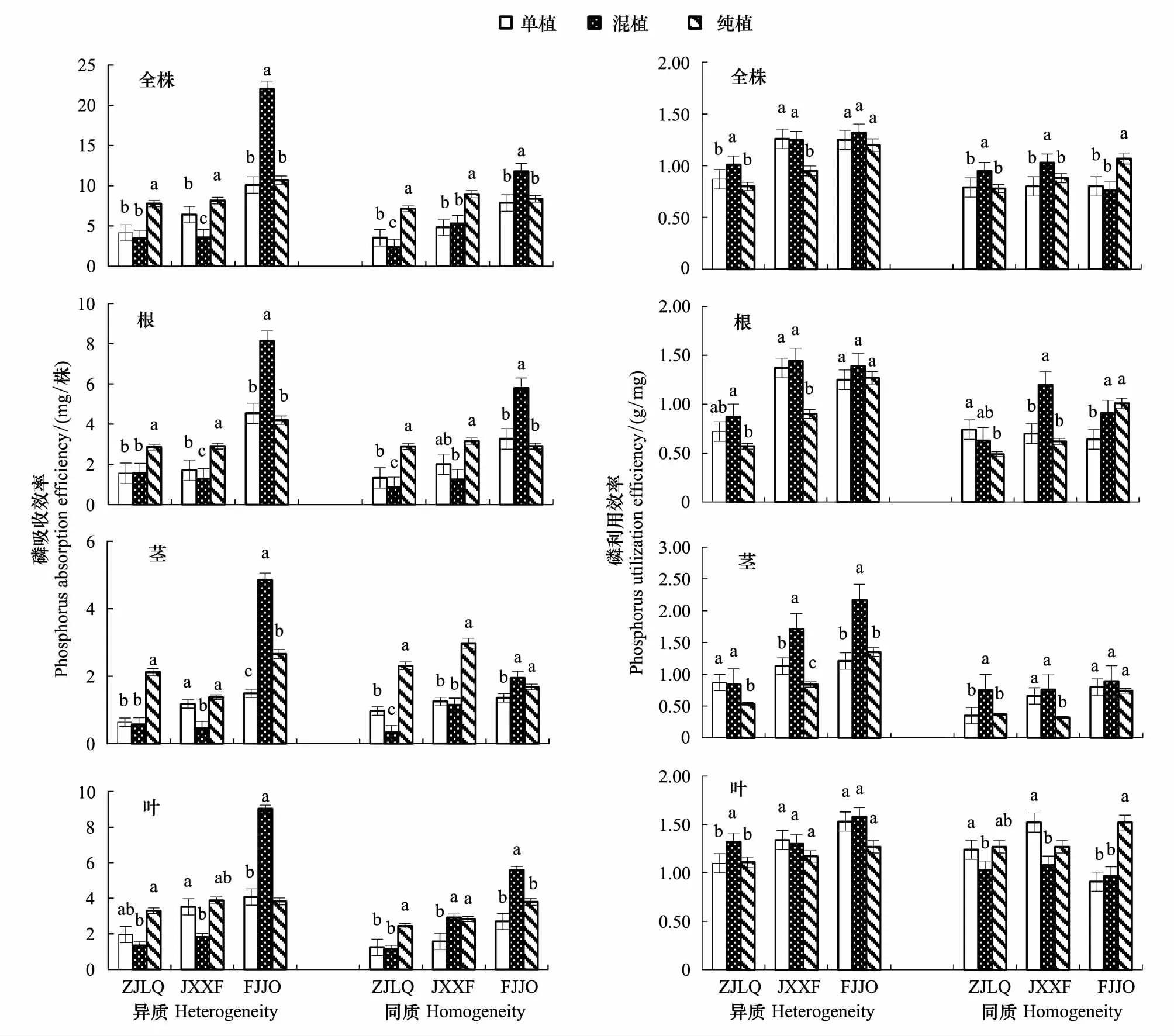
图2 栽植方式对3个木荷种源的磷吸收效率与利用效率的影响Fig.2 Effect of planting patterns on phosphorus absorption and utilization efficiency of three provenances of S. superba
3 讨论与结论
林木的觅养行为非常复杂,不仅因树种或品种而异,而且还与土壤养分斑块有关。当遭遇高度异质性土壤养分环境时,植物根系通过各种形态和生理塑性反应更多地从土壤中吸收养分并促进生长[19],本研究也证实了这一结论,在异质养分环境中木荷种源干物质量、氮、磷的吸收效率及磷利用效率均显著高于同质养分环境,且种源效应显著,以福建建瓯种源最高。而当邻株存在时,目标植物的根系和生理形态特性会进一步被调节[19],本文中在异质养分环境之异种混植模式下,福建建瓯种源的干物质量及氮磷效率指标明显的高于单植和双株纯植模式,表现出较强的生长优势。
不同养分环境中,植物生长的差异往往与其生物量分配有密切关系[20],通常认为当养分受胁迫时植物会向根系分配较高比例的生物量,以保证体内养分平衡和养分吸收能力[21],而在养分充足时,营养物质不再是植物生长的限制因子,植物将分配较多的生物量至地上部分,提高其光合的能力,更好地促进植物生长[22- 23]。本研究中,在异质养分环境中,福建建瓯种源生物量分配(根冠比)表现出较高的表型可塑性,混植时根冠比显著减小,将更多干物质分配到地上部分,用于高径生长和叶片光合所需,进而产生更多的干物质。相比之下,浙江龙泉种源则将更多干物质投入到地下部分,结果影响了地上部分和干物质积累,江西信丰种源干物质分配调节能力较弱,其生长情况一般。因此福建建瓯种源较高的生物量分配塑性使其在混植栽培时较其他种源生长优势明显。
植物生长优势(干物质生产能力)的生物学基础在于资源的有效利用,在植物营养方面主要是养分吸收效率的增加和养分利用效率的提高[24]。已有的研究表明,许多植物氮、磷吸收和利用效率存在丰富遗传多样性[25- 27]。Zhang等[28]研究表明,不同种源木荷的磷素吸收和利用效率表现出明显差异,并证明了木荷种源吸收效率与干物质量的呈正相关关系。本研究结果显示,福建建瓯种源的氮、磷吸收效率不仅显著高于江西信丰和浙江龙泉种源,而且其生理可塑性较强,在异质养分环境下混植时,其全株及各器官氮、磷吸收效率较单植高1—2倍,这与其较高的干物质量相一致。但混植时并未明显增加其氮、磷利用效率,表明福建建瓯种源的生长优势在于氮、磷吸收效率的增加,而非利用效率的改变。养分吸收效率和利用效率对于植物生长的相对重要性,一般因植物种类或基因型不同而有所差异。梁霞等[29]认为磷高效杉木(Cunninghamialanceolata)无性系具有较高的磷素吸收效率和利用效率。而宋平等[30]则研究发现,在异质磷环境下,磷吸收效率对马尾松(Pinusmassoniana)无性系干物质量的促进作用更为突出,而干物质量与磷利用效率之间的相关性较弱,甚至负相关,这与本试验结果相似。
植物的养分效率是其对养分吸收、转运和利用等效率综合作用的结果[35]。本试验观测到,混植时福建建瓯种源的根、茎和叶的氮、磷素吸收与利用效率均最大,说明其氮、磷素吸收、转运及利用能力最强,从而最终实现了较高的生产力。与之不同的是,江西信丰和浙江龙泉种源各器官的氮、磷吸收和利用效率变化规律不一致,使得其干物质量较低。由于树木生活史较长,各阶段响应异质环境资源的机制差异较大[36],需要延长试验时间,或采用不同基因型木荷造林对该试验结果做进一步观测,以期更全面地揭示植物觅养塑性与种源生长竞争优势的关联性。
参考文献(References):
[1] Warren C R. How does P affect photosynthesis and metabolite profiles ofEucalyptusglobulus? Tree Physiology, 2011, 31(7): 727- 739.
[2] Huang W J, Zhou G Y, Liu J X. Nitrogen and phosphorus status and their influence on aboveground production under increasing nitrogen deposition in three successional forests. Acta Oecologica, 2012, 44: 20- 27.
[3] Ostertag R. Foliar nitrogen and phosphorus accumulation responses after fertilization: an example from nutrient-limited Hawaiian forests. Plant and Soil, 2010, 334(1/2): 85- 98.
[4] 冯广, 姚兰, 艾训儒, 黄继红, 路兴慧, 丁易, 臧润国. 鄂西南亚热带常绿落叶阔叶混交林土壤空间异质性. 生态学报, 2016, 36(23): 7635- 7643.
[5] Tsunoda T, Kachi N, Suzuki J I. Interactive effects of soil nutrient heterogeneity and belowground herbivory on the growth of plants with different root foraging traits. Plant and Soil, 2014, 384(1/2): 327- 334.
[6] Zhang Y, Ma X H, Zhou Z Z. The influence of light conditions and interspecific competition on the root foraging traits and seedling growth of two tree species. Plant Biosystems, 2012, 146(1): 7- 14.
[7] 李洪波. 植物响应养分空间异质性分布的动态过程及调控根系获取养分的策略研究[D]. 北京: 中国农业大学, 2014.
[8] Mou P, Jones R H, Tan Z Q, Bao Z, Chen H M. Morphological and physiological plasticity of plant roots when nutrients are both spatially and temporally heterogeneous. Plant and Soil, 2013, 364(1/2): 373- 384.
[9] Cahill J F Jr, McNickle G G, Haag J J, Lamb E G, Nyanumba S M, St. Clair C C. Plants integrate information about nutrients and neighbors. Science, 2010, 328(5986): 1657- 1657.
[10] Rajaniemi T K. Root foraging traits and competitive ability in heterogeneous soils. Oecologia, 2007, 153(1): 145- 152.
[11] 吴开贤, 安曈昕, 范志伟, 周锋, 薛国峰, 吴伯志. 根间相互作用对玉米与马铃薯响应异质氮的调控. 生态学报, 2015, 35(2): 508- 516.
[12] Fransen B, de Kroon H, Berendse F. Soil nutrient heterogeneity alters competition between two perennial grass species. Ecology, 2001, 82(9): 2534- 2546.
[13] 辛娜娜, 张蕊, 范辉华, 陈康, 徐有明, 周志春. 5年生木荷生长和形质性状的家系变异和选择. 林业科学研究, 2014, 27(3): 316- 322.
[14] 王秀花, 陈柳英, 马丽珍, 范辉华, 叶穗文, 周志春. 7年生木荷生长和木材基本密度地理遗传变异及种源选择. 林业科学研究, 2011, 24(3): 307- 313.
[15] 周志春, 范辉华, 金国庆, 王月生, 陈奕良, 饶龙兵. 木荷地理遗传变异和优良种源初选. 林业科学研究, 2006, 19(6): 718- 724.
[16] 苏松锦, 刘金福, 何中声, 洪伟, 张金彪. 格氏栲天然林土壤养分空间异质性. 生态学报, 2012, 32(18): 5673- 5682.
[17] 张万儒, 杨光滢, 屠星南, 张萍. 森林土壤分析方法. 北京: 中国标准出版社, 2000: 74- 115.
[18] 曹靖, 张福锁. 低磷条件下不同基因型小麦幼苗对磷的吸收和利用效率及水分的影响. 植物生态学报, 2000, 24(6): 731- 735.
[19] Zhang D S, Zhang C C, Tang X Y, Li H G, Zhang F S, Rengel Z, Whalley W R, Davies W J, Shen J B. Increased soil phosphorus availability induced by faba bean root exudation stimulates root growth and phosphorus uptake in neighbouring maize. New Phytologist, 2016, 209(2): 823- 831.
[20] 常瑞英. 养分水平和氮磷比对入侵植物空心莲子草与非入侵种竞争关系的影响[D]. 济南: 山东大学, 2013.
[21] 陈智裕, 李琦, 邹显花, 马祥庆, 吴鹏飞. 邻株竞争对低磷环境杉木幼苗光合特性及生物量分配的影响. 植物生态学报, 2016, 40(2): 177- 186.
[22] 冷华妮, 陈益泰, 段红平, 饶龙兵, 王永军, 胡韵雪. 磷胁迫对不同种源枫香生长及氮、磷吸收利用率的影响. 应用生态学报, 2009, 20(4): 754- 760.
[23] Schmid C, Bauer S, Bartelheimer M. Should I stay or should I go? Roots segregate in response to competition intensity. Plant and Soil, 2015, 391(1/2): 283- 291.
[24] 乔振江, 蔡昆争, 骆世明. 低磷和干旱胁迫对大豆植株干物质积累及磷效率的影响. 生态学报, 2011, 31(19): 5578- 5587.
[25] Gaju O, Allard V, Martre P, Snaped J W, Heumeze E, LeGouis J, Moreau D, Bogard M, Griffiths S, Orford S, Hubbart S, Foulkes M J. Identification of traits to improve the nitrogen-use efficiency of wheat genotypes. Field Crops Research, 2011, 123(2): 139- 152.
[26] Beatty P H, Anbessa Y, Juskiw P, Carroll R T, Wang J, Good A G. Nitrogen use efficiencies of spring barley grown under varying nitrogen conditions in the field and growth chamber. Annals of Botany, 2010, 105(7): 1171- 1182.
[27] Fageria N K, de Morais O P, dos Santos A B. Nitrogen use efficiency in upland rice genotypes. Journal of Plant Nutrition, 2010, 33(11): 1696- 1711.
[28] Zhang R, Zhou Z C, Luo W J, Wang Y, Feng Z P. Effects of nitrogen deposition on growth and phosphate efficiency ofSchimasuperbaof different provenances grown in phosphorus-barren soil. Plant and Soil, 2013, 370(1/2): 435- 445.
[29] 梁霞, 刘爱琴, 马祥庆, 冯丽贞, 陈友力. 磷胁迫对不同杉木无性系酸性磷酸酶活性的影响. 植物生态学报, 2005, 29(1): 54- 59.
[30] 宋平, 张蕊, 张一, 周志春, 丰忠平. 模拟氮沉降对低磷胁迫下马尾松无性系细根形态和氮磷效率的影响. 植物生态学报, 2016, 40(11): 1136- 1144.
[31] 刘敏娜, 刘晓霞, 丁文雅, 陈秋会, 林咸永. 不同菠菜基因型氮素吸收与利用效率的差异及其评价. 浙江大学学报: 农业与生命科学版, 2012, 38(5): 599- 607.
[32] Garnett T, Conn V, Kaiser B N. Root based approaches to improving nitrogen use efficiency in plants. Plant, Cell & Environment, 2009, 32(9): 1272- 1283.
[33] 姚甲宝, 楚秀丽, 周志春, 童建设, 王晖, 余家中. 不同养分环境下木荷种源生长和根系发育对邻株竞争响应的差异. 应用生态学报, 2017, 28(4): 1087- 1093.
[34] 李洪波, 薛慕瑶, 林雅茹, 申建波. 土壤养分空间异质性与根系觅食作用: 从个体到群落. 植物营养与肥料学报, 2013, 19(4): 995- 1004.
[35] 何鹏, 吴敏, 韦家少, 覃怀德, 吴炳孙. 不同磷水平对橡胶树幼苗氮钾吸收、分配与利用的影响. 中国农学通报, 2011, 27(16): 1- 6.
[36] 冯晓燕, 刘宁, 郭晋平, 张芸香. 控制光照条件下华北山地4个混交树种幼苗幼树的形态响应和可塑性. 林业科学, 2013, 49(11): 42- 50.
