桂林岩溶石山灌丛植物叶功能性状和土壤因子对坡向的响应
2018-04-19盘远方陈兴彬梁士楚陆志任黄宇欣倪鸣源覃彩丽刘润红
盘远方,陈兴彬,姜 勇,*,梁士楚,陆志任,黄宇欣,倪鸣源,覃彩丽,刘润红
1 广西师范大学珍稀濒危动植物生态与环境保护教育部重点实验室,广西师范大学生命科学学院, 桂林 541006 2 江西省林业科学院;江西省植物生物技术重点实验室, 南昌 330013
近年来,植物性状的变化与环境的关系成为植物生态学的研究热点之一。植物在漫长的进化和发展过程中,与环境相互作用,逐渐形成能够适应外部环境变化的形态和生理结构,主要体现在叶片、根系、种子等植物性状的差别上,这种能响应环境变化的植物性状被称为植物功能性状[1- 2]。叶功能性状是植物的重要特性之一,被广泛应用于植物功能性状研究,与植物生长对策及利用资源的能力紧密联系,体现了植物为获得最大化碳收获所采取的生存策略[3- 4]。叶功能性状主要包括生理性状(叶光合速率、组织氮含量、氮和水利用率)和叶面积分配性状(比叶面积、叶干物质含量、叶面积指数和叶片厚度),其中比叶面积(specific leaf area,SLA)、叶干物质含量(leaf dry matter content,LDMC)、叶片厚度(leaf thichness,LT)等易于测定,常被广泛应用于不同尺度叶性状研究中。SLA与植物相对生长速率、光合速率、叶氮含量有关,可用来反映植物的碳获取策略,对植物的相对生长速率具有重要影响,是生理过程权衡的最佳指示者[5- 6]。具有较大SLA的植物其叶片的资源捕获面积大,因而具有较高的净光合速率[7],具有较低SLA的植物能更好的适应资源贫瘠和干旱的环境[8]。叶干物质含量(LDMC)是植物获取资源的预测指标[9],反映叶片对干旱气候的适应程度,是资源获取轴上最稳定的变量。叶片厚度通常被作为一个非常有价值的性状特性,它与资源获取、水分保存和同化有关。李永华等对青海省沙珠玉治沙站17种主要植物叶性因子的比较研究表明:叶片厚度或密度的增加,有利于增加叶片内部水分向叶片表面扩散的距离或阻力,降低植物内部水分散失[10],同时刘金环等研究发现SLA和LDMC与LT之间存在某种关系,一般LT较厚的植物,其SLA和LDMC通常较小[8]。
桂林位于广西壮族自治区东北部,具有典型的岩溶石山生态环境特征,岩溶发育强烈、景观类型复杂、生态系统极为脆弱,这些特殊的生境条件,构成了石灰岩地区植物组成上的特殊性。Christianson和Niklas对美国东北部岩溶生境下冠层银杏叶多样性分布格局研究发现:土壤含水量是影响比叶面积的主要驱动因子[11]。Du等对中国西南喀斯特生境下的森林植物功能性状与土壤环境因子的探究表明:土壤含水量与叶干物质含量呈显著的负相关[12],Liu等对中国西南亚热带喀斯特落叶与常绿阔叶混交林的研究显示:植物叶干物质含量与比叶面积呈显著的负相关[13]。近年来,国内学者对岩溶石山植物功能性状及功能性状与环境耦合关系等方面进行了初步探索,但与热带雨林功能性状的丰富研究资料相比[14-15],岩溶石山植物功能性状的研究起步较晚,资料相对零散。例如,刘宏伟等研究了中梁山石灰山30种主要植物的比叶面积(SLA)和叶干物质含量(LDMC)等6个叶片性状间的变异[16],习新强等研究了黔中喀斯特山区植物的比叶面积(SLA)和叶面积(LA)在灌丛、落叶阔叶林、落叶常绿混交林3个演替阶段的变化规律[17]及马姜明等对桂林尧山常见植物叶功能性状的初步探究[18]。且从阴坡-阳坡环境梯度角度探讨岩溶石山植物叶片性状和环境因子变化规律的研究尚未有报道。阴坡和阳坡是最常见的两种生境,往往营造出局部的微气候生境,促使水分分布和太阳辐射的空间再分配[19],而岩溶石山典型的特点是峰丛地貌,具有明显的坡向变化。在不同的坡向上,土壤水分和温度具有明显差异,从而使得土壤养分的分布产生巨大的差异[20]。本文以桂林岩溶石山为研究对象,探讨岩溶石山地区主要植物叶功能性状比叶面积(SLA)、叶干物质含量(LDMC)、叶片厚度(LT)和环境因子在坡向梯度(阴坡-阳坡)的变化规律,揭示微气候生境对岩溶石山植物群落空间格局形成影响,以及阴阳坡植物生存策略的变化,为我国西南岩溶石山地区植被重建与恢复及生态系统服务功能评价提供科学依据。
1 研究地区与方法
1.1 研究地点概况
研究样地设在桂林境内临桂县西南部的典型溶岩石山,地理坐标为(24°38′—25°39′N,110°9′—110°42′E)。岩溶石山地貌特征显著(土层薄、植被不连续、地表水缺乏和地形破碎)[16],植被类型以灌丛为主,主要的优势种有马鞭草科的牡荆(Vitexnegundo)、大戟科的红背山麻杆(Alchorneatrewioides)和山麻杆(Alchorneadavidii) 、千屈菜科的紫薇(Lagerstroemiaindica)、金缕梅科的檵木(Loropetalumchinense)和豆科的龙须藤(Bauhiniachampionii)等。气候属于中亚热带湿润季风气候,雨量充沛,气候温和。年平均气温19℃,最热的8月份平均气温28℃,最冷的1月份平均气温8℃,全年无霜期309d,年平均降雨量为1856.7mm,降雨量年分配不均,秋、冬季干燥少雨,年平均蒸发量为1458.4mm[21]。
1.2 群落调查和样品取样
经过踏查,于2016年7—11月在桂林境内临桂县西南部岩溶石山,沿山体中部到山底,设置了24个采样点,分为阴坡和阳坡,样方大小为10m×10m,样地概况如表1所示。本研究采用每木调查法,记录每个样方内基径≥1cm的所有木本植物的种名、多度、基径、树高和空间坐标,同时记录岩石裸露率,利用手持GPS进行经纬度、海拔数据测定。同时对样方内的所有物种个体进行叶功能性状取样和测定,对不认识的物种采集回实验室进行标本鉴定,叶功能性状指标包括比叶面积(SLA,cm2/g)、叶干物质含量(LDMC,g/g)和叶片厚度(LT,mm)。此外,采用梅花五点法[22],在每个样方的4个顶点和中心点位置用土钻钻0—15cm的土样混合用作土壤养分分析,每个样方3个重复。
1.3 测定的项目与方法
1.3.1 土壤因子的测定
土壤含水量(SWC,%)采用烘干法测定,土壤温度(Tem,℃)通过土壤三参数仪(W.E.T Sensor Kit)获取,土壤pH值采用电导法测定(水土比为2.5∶1)[23];总有机碳含量(TOC,g/kg)和有机质含量(SOM,g/kg)采用重铬酸钾容量法测定[24];全氮含量(TN,g/kg)采用全自动凯氏定氮仪(KJELTECTM8400)测定[24];有效氮含量(available nitrogen,AN,g/kg)采用碱解扩散法测定[25];全磷含量(TP,g/kg)采用钼锑抗比色法测定[26]。
1.3.2 叶性状取样与测定
物种个体水平的叶功能性状取样和测定方法如下:对于样方内出现的每一株植物,均采集3—5片完好的向阳成熟的叶片,总共采了83个物种3167株植物。用精度为0.01mm的电子数显卡尺测量叶片的厚度,在获取的植物叶片主脉方向均匀选3个点,厚度取其3个点的平均值;采用精度为0.001g的电子天平称量叶片鲜重;使用叶面积仪(Yaxin- 1241)测定叶面积,随后叶片样品置于70°C的烘箱烘干72h后,称重测定叶片干重。
群落水平叶功能性状值(CWM)是由测定的物种个体水平的叶功能性状值,以物种多度为基础进行加权平均,从而分别得到各个性状在群落水平的平均值。具体的群落水平上的叶功能性状值计算由R.2.14.0里面的FD软件包完成[22]。
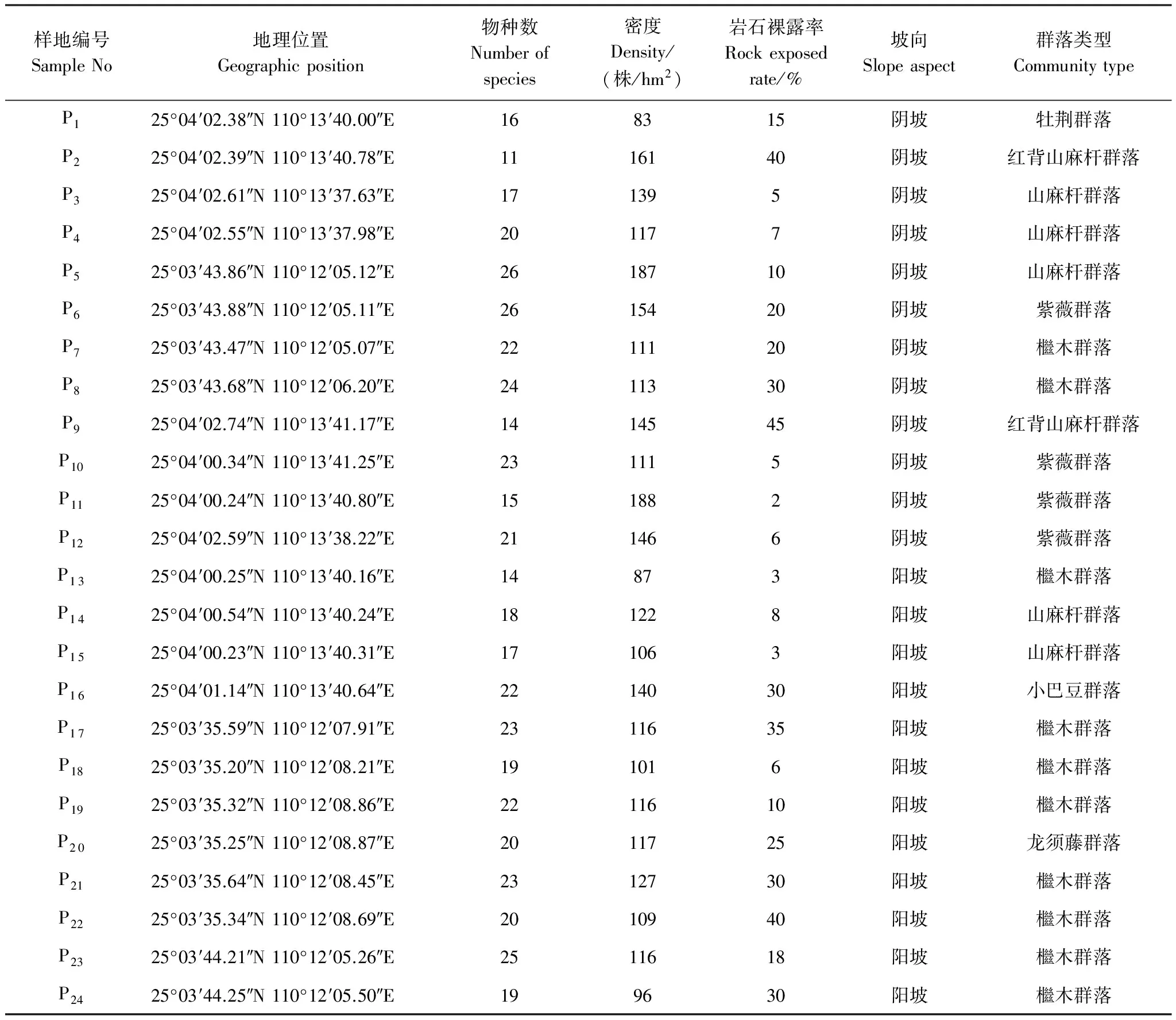
表1 不同样地生境基本概况
1.4 数据处理
采用Wilcoxon加符秩检验方法对不同坡向梯度(阴-阳坡)生境中叶功能性状和土壤环境因子进行差异性检验。进一步探究哪些环境因子对阴-阳坡群落水平上植物叶功能性状起重要影响,研究采用多元逐步回归方法,以每个叶功能性状作为因变量,土壤环境因子作为自变量进行模型回归拟合。所有数据统计分析均用R 2.14.0软件(R Development Core Team,2011)完成。
2 结果与分析
2.1 不同坡向土壤理化性质的变化
由图1可以看出,除土壤pH值和土壤全磷含量无显著差异外,其他土壤环境因子对不同坡向微生境产生显著的响应(P<0.05)。其中土壤温度和土壤全氮含量为阳坡大于阴坡;土壤有机质含量、土壤总有机碳含量和土壤有效氮含量为阴坡大于阳坡;土壤含水量为阴坡大于阳坡。
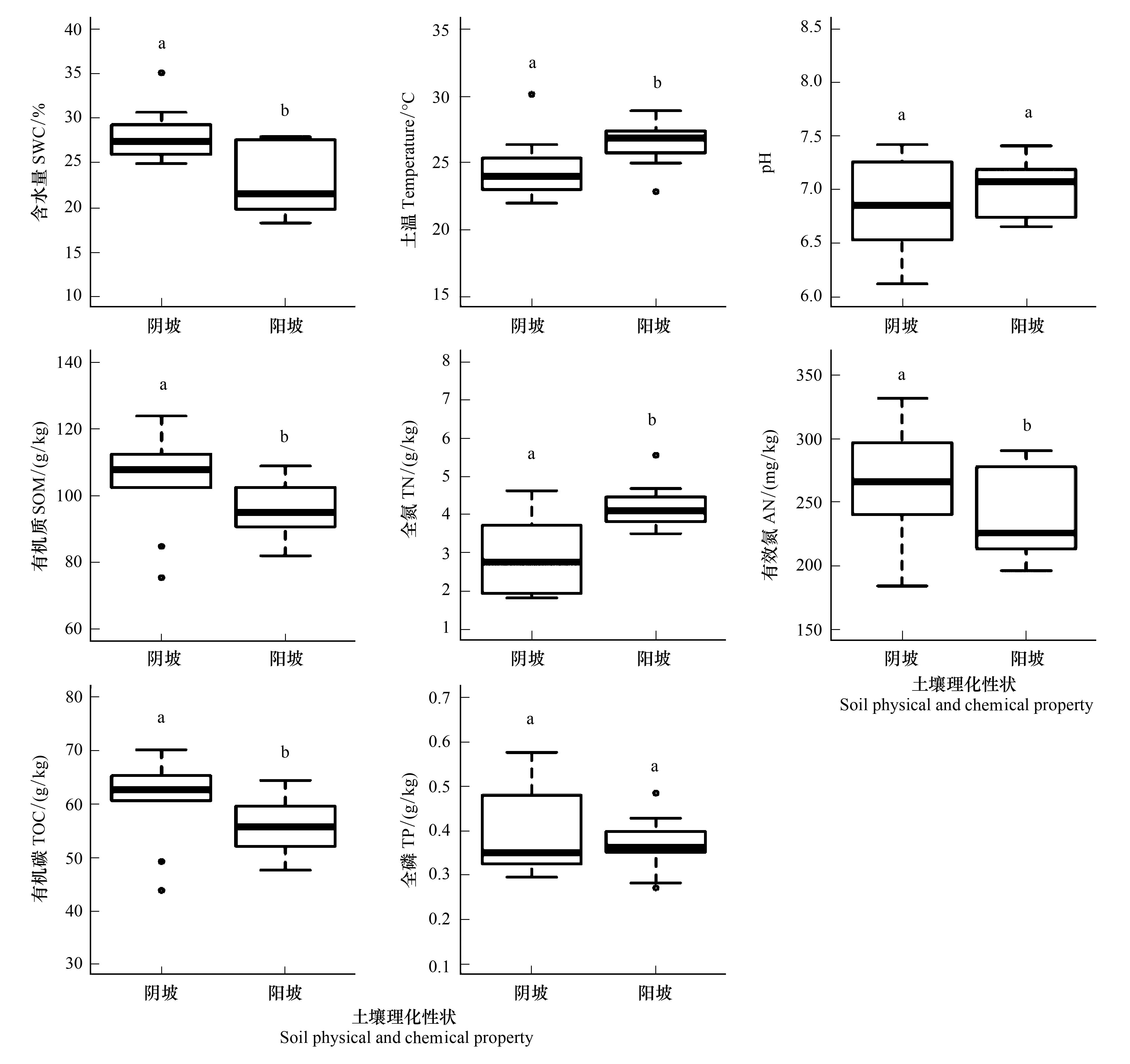
图1 不同样地土壤理化性状Fig.1 Soil physical and chemical properties of different sampling sitesSWC:土壤含水量 soil water content;Tem:土温 soil temperature;pH:酸碱度 power of hydrogen;SOM:有机质 soil organic matter;TN:全氮 total nitrogen;AN:有效氮 available nitrogen;TOC:总有机碳 total organic carbon;TP:全磷 total phosphorus
2.2 不同坡向植物叶功能性状的变化
由图2可以看出,群落水平叶功能性状(比叶面积、叶干物质含量和叶片厚度)在不同坡向梯度(阴-阳坡)生境中,差异性均显著(P<0.05)。其中比叶面积为阴坡大于阳坡,叶干物质含量和叶片厚度为阳坡大于阴坡。
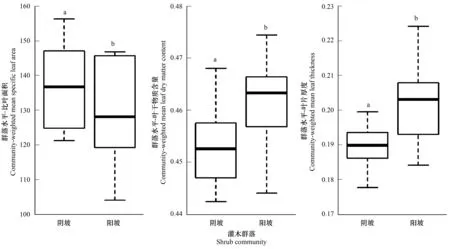
图2 不同坡向植物群落功能性状Fig.2 Functional traits of plant communities in different aspectsCWM_SLA:群落水平-比叶面积 Community-weighted mean specific leaf area;CWM_LDMC:群落水平-叶干物质含量 Community-weighted mean leaf dry matter content;CWM_LT:群落水平-叶片厚度 Community-weighted mean leaf thickness
2.3 植物叶功能性状与土壤因子
多元逐步回归结果表明,如表2所示,在阴坡上,对比叶面积有显著影响的是土壤有效氮含量(P<0.05),对叶干物质含量有显著影响的是土壤温度和土壤总有机碳含量(P<0.05),环境因子对叶片厚度作用不显著。在阳坡上,对比叶面积有显著影响的环境因子是土壤含水量和土壤pH值(P<0.05),对叶干物质含量有显著影响的环境因子是土壤全氮含量(P<0.05),对叶片厚度有显著影响的环境因子是土壤温度、土壤全氮含量和土壤全磷含量(P<0.05)。
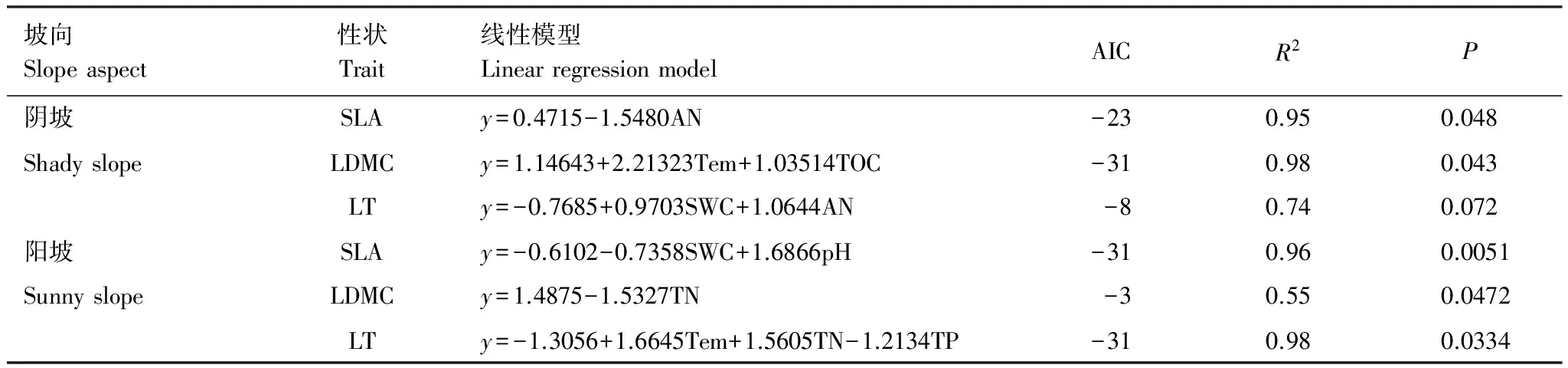
表2 植物功能性状与环境因子的线性回归模型
R:赤池信息准则 Akaike information criterion;R2:判定系数 R Square;SLA:比叶面积 Specific leaf area;LDMC:叶干物质含量 Leaf dry matter content;LT:叶片厚度 Leaf thickness;AN:有效氮 Available nitrogen;Tem:土温 Soil temperature;TOC:总有机碳 Total organic carbon;SWC:土壤含水量 Soil water content;pH:酸碱度 Power of hydrogen;TN:全氮 Total nitrogen;TP:全磷 Total phosphorus;AIC: 最小信息准则Akaike information criterion
3 讨论
3.1 土壤因子对坡向的响应
植物的生长发育情况不仅取决于自身的生理特性,环境因子对其生长将产生深刻的影响,这是一种间接作用,是地形因子通过影响其他一些生态因子而作用于植物。图1显示土壤含水量阴坡大于阳坡,而土壤温度阳坡大于阴坡,这与刘旻霞等对高寒草甸阳坡-阴坡梯度上环境因子特征及其与地上生物量和物种丰富度的关系研究中土壤含水量与土壤温度具有显著负相关的结论一致,阴坡-阳坡土壤含水量的变化可能是由阴阳坡的温度差异造成的[20]。土壤有机质、总有机碳和有效氮含量阴坡明显高于阳坡(图1),表明阴坡具有相对较高的土壤营养资源[27],可能是坡向通过改变坡面水热条件、植被分布及土壤运移,进而影响腐殖质的积累加强,造成土壤有机物的输入、矿化以及运移,使不同坡向土壤总有机碳的累积出现显著差异。朱猛等对祁连山森林草原带坡面尺度土壤总有机碳分布研究表明总有机碳含量与土壤水分含量呈正相关关系[28];朱云云等对坡向因子对黄土高原草地群落功能多样性的影响进行研究,结果表明:阳坡土壤风化严重,且在雨季水土流失严重及土壤矿化[29];汝海丽等对黄土丘陵区微地形条件下草本群落特征与土壤水分及养分关系分析表明:阳坡接受的太阳辐射量大于阴坡,使得阳坡土壤温度高[30],而岩溶石山阳坡风化和水土流失强烈,土壤养分难以富集,加之植被凋落物有机质输入较少,土壤容重增加、氮素和持水减少,土壤退化[31],从而导致在阴坡土壤有机质、总有机碳和有效氮含量上明显高于阳坡,这与本研究结果一致。尽管阴坡-阳坡的土壤养分含量有差异,但不同植物通过其自身的吸收和调节机制维持叶内养分的相对平衡,这是植物对生存环境的适应结果[32]。
3.2 植物叶功能性状对坡向的响应
坡向是一种重要的地形因子,影响了坡面接受的太阳光辐射和地面与风向的夹角[33],坡向与光照强度和光照面积呈负相关,与土壤含水量之间呈负相关[34],地形特征通过光照、温度和降水等生态因子的再分配间接影响植被的生长。从阳坡到阴坡的生境变化过程中,环境因子发生了显著变化,阳坡的土壤养分含量和水分含量都比较低,土壤比较贫瘠,植物平均比叶面积都比较小。对比下阴坡生境资源较充足,植物比叶面积性状显著大于阳坡[35]。比叶面积反映植物获取和利用资源的能力,与植物的生存对策有密切的联系。通常情况下,比叶面积较高的植物,其叶片的光捕获面积较高,而比叶面积较低的植物,叶的大部分物质用于构建保卫构造,常形成厚度较大而面积较小的叶片[32]。图1、图2显示,阳坡的比叶面积和土壤含水量显著低于阴坡,且阴坡土壤养分资源相对阳坡较丰富,植被可能在水力条件胁迫下加大了叶片生物量的生长速度,使植被比叶面积减小,这样有利于植被在水分较少的环境中储存水分,也增强了叶片对强光的防护作用[16,36],因此植被的比叶面积在水分较好的阴坡中比较大。叶干物质含量是植物生态学研究中的首选指标,它可以很好的指示植物对养分元素的保存能力[37],图2显示,叶干物质含量对坡向具有依赖性可能是由于植被应对不同环境的胁迫表现出构型和生物量分配策略分异。党晶晶对祁连山高寒草地甘肃臭草叶性状与坡向间的关系研究表明:光照条件充足的阳坡并不需要通过叶面积的增加而增大光合速率,反而阳坡向上土壤水分最低,致使水力限制增加,植株可能会将叶生物量投资于机械支持结构与微管结构,而且大叶的呼吸和蒸腾成本更高,不利于其光拦截能力和光合碳获取能力最大化[19],叶干物质含量的增加,使叶片内部水分向叶片表面扩散的距离或阻力增大,降低植物内部水分散失[36],因此阳坡植被具有较高的叶干物质含量,这可能提高其对环境适合度。阳坡光照充足,植物叶片为了避免水分大量丧失,限制了细胞的分裂和生长,植物生长缓慢[38]。阳坡在水分和养分比较有限的条件下植物会投资更多的物质用于防卫或增加叶肉细胞密度,形成厚而小的叶片[39],阴坡温度低于阳坡而土壤含水量高于阳坡,良好的水热条件有利于细胞体积和数目增加,从而促进了叶面积的扩展[40]。本研究中,在接受光照最为强烈、土壤含水量较少的阳坡,叶片尽管能够获得最多的光照资源,但受到了水分等因子的限制,叶片出于防卫而增加细胞密度,而在阴坡水热条件良好有利于细胞体积和数目增加,从而促进了叶面积的扩展,因此在阳坡有较大的叶片厚度(LT)。所以,坡向的梯度变化是造成岩溶石山植物叶功能性状差异的主要原因。
3.3 植物叶功能性状与土壤因子的关系
土壤养分含量高低决定植物利用资源的策略方式。其一,通过塑造群落功能性状的表型特征来影响不同坡向植物的策略;其二,通过影响物种内的性状变异以影响植物策略的改变[32]。本研究中,植物叶功能性状如比叶面积、叶干物质含量和叶片厚度与土壤温度、氮含量、总有机碳含量和磷含量随不同的坡向梯度变化均有显著的相关关系[41]。表2显示,在阴坡上,对比叶面积产生显著影响的是土壤有效氮含量,可能是有效氮是土壤中最重要的一种活性氮的形态[42],且阴坡水分充足,水土流失作用相对阳坡较弱,致使退化的土壤养分和物理性能得到改善,因此土壤有效氮资源丰富,能被植物直接吸收利用[43-44]。而在阳坡上,对比叶面积产生显著影响的是土壤含水量和pH,主要是因为岩溶石山阳坡是以易风沙化的碱性石灰土为主,水分蒸发量大,在地表水与地下水资源匮乏条件下植物会投资更多的物质用于防卫或增加叶肉细胞密度[45]。叶干物质含量主要反映的是植物对养分元素的保存能力[46],在阴坡上,对叶干物质含量产生显著影响的是土壤温度和土壤总有机碳含量,这可能是阴坡土壤水热条件合适,有利于土壤有机物分解,因此,植物叶片为了增加碳获取资源,将生物量主要投资于叶,以加大光捕获面积,获得更高的光合能力[47]。而在阳坡上,对叶干物质含量产生显著影响的是土壤全氮含量,可能是因为,岩溶石山阳坡风化和水土流失严重,加之植被凋落物有机质输入较少,土壤氮营养元素难以富集,土壤容重增加和持水减少[31],因此溶解性的氮源相对较低[48]。在土壤资源缺乏的环境中,植物会投资更多物质用于细胞的构建,使植物能很好地适应贫瘠的环境[49]。而在阳坡上,对叶片厚度产生显著影响的环境因子是土壤温度、土壤全氮含量和土壤全磷含量,可能是氮磷元素是植物生长发育最主要的限制因子。植物氮磷含量取决于土壤养分供应和植物氮磷需求间的动态平衡[50],土壤全磷含量较低时,植物在低营养环境中可能采取较为保守的营养投资策略[51],且在阳坡上,土壤温度偏高,导致土壤水分大量蒸发[10],植物为了降低内部水分散失,植物会加大叶片生物量的增长速度,增加叶片厚度,从而提高叶片内部水分向叶片表面扩散的距离或阻力[52]。因此,同一叶功能性状在阴坡-阳坡梯度上受到不同环境因子的控制,同时会对所处的生境产生一定的适应性。
4 结论
通过对岩溶石山不同坡向梯度(阴-阳坡)生境中叶功能性状和土壤环境因子进行研究。结果显示:比叶面积为阴坡大于阳坡,叶干物质含量和叶片厚度为阳坡大于阴坡,表明阴坡水热条件良好,从而促进了叶面积的扩展。土壤含水量、有机质含量、总有机碳含量和有效氮含量为阴坡大于阳坡,土壤温度和全氮含量为阳坡大于阴坡,表明在岩溶石山地区阴坡的土壤资源比较丰富,阳坡土壤资源较贫瘠。尽管不同坡向梯度土壤养分含量不同,但植物叶功能性状采取不同的生存策略来适应坡向梯度(阴-阳坡)的微生境,揭示了微气候生境对岩溶石山植物群落叶功能性状的影响,为我国西南岩溶石山地区植被重建与恢复及生态系统服务功能评价提供科学依据。
参考文献(References):
[1] Violle C, Navas M L, Vile D, Kazakou E, Fortunel C, Hummel I, Garnier E. Let the concept of trait be functional! Oikos, 2007, 116(5): 882- 892.
[2] Cornelissen J H C, Lavorel S, Garnier E, Díaz S, Buchmann N, Gurvich D E, Reich P B, Steege H T, Morgan H D,van der Heijden M G A, Pausas J G, Poorter H. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Australian Journal of Botany, 2003, 51(4): 335- 380.
[3] Taylor S H, Franks P J, Hulme S P, Spriggs E, Christin P A, Edwards E J, Woodward F I, Osborne C P. Photosynthetic pathway and ecological adaptation explain stomatal trait diversity amongst grasses. New Phytologist, 2012, 193(2): 387- 396.
[4] 陈文, 王桔红, 马瑞君, 齐威, 刘坤, 张丽娜, 陈学林. 粤东89种常见植物叶功能性状变异特征. 生态学杂志, 2016, 35(8): 2101- 2109.
[5] 杨冬梅, 章佳佳, 周丹, 钱敏杰, 郑瑶, 金灵妙. 木本植物茎叶功能性状及其关系随环境变化的研究进展. 生态学杂志, 2012, 31(3): 702- 713.
[6] Donovan L A, Maherali H, Caruso C M, Huber H, de Kroon H. The evolution of the worldwide leaf economics spectrum. Trends in Ecology & Evolution, 2011, 26(2): 88- 95.
[7] 张莉, 温仲明, 苗连朋. 延河流域植物功能性状变异来源分析. 生态学报, 2013, 33(20): 6543- 6552.
[8] 刘金环, 曾德慧, Lee D K. 科尔沁沙地东南部地区主要植物叶片性状及其相互关系. 生态学杂志, 2006, 25(8): 921- 925.
[9] 周欣, 左小安, 赵学勇, 刘川, 吕朋. 科尔沁沙地植物功能性状的尺度变异及关联. 中国沙漠, 2016, 36(1): 20- 26.
[10] 李永华, 罗天祥, 卢琦, 田晓娅, 吴波, 杨恒华. 青海省沙珠玉治沙站17种主要植物叶性因子的比较. 生态学报, 2005, 25(5): 994- 999.
[11] Christianson M L, Niklas K J. Patterns of diversity in leaves from canopies ofGinkgobilobaare revealed using Specific Leaf Area as a morphological character. American Journal of Botany, 2011, 98(7): 1068- 1076.
[12] Du H, Peng W X, Song T Q, Zeng F P, Wang K L, Song M, Zhang H. Spatial pattern of woody plants and their environmental interpretation in the karst forest of southwest China. Plant Biosystems, 2015, 149(1): 121- 130.
[13] Liu C C, Liu Y G, Guo K, Zhao H W, Qiao X G, Wang S J, Zhang L, Cai X L. Mixing litter from deciduous and evergreen trees enhances decomposition in a subtropical karst forest in southwestern China. Soil Biology and Biochemistry, 2016, 101: 44- 54.
[14] Jiang Y, Zang R G, Lu X H, Huang Y F, Ding Y, Liu W D, Long W X, Zhang J Y, Zhang Z D. Effects of soil and microclimatic conditions on the community-level plant functional traits across different tropical forest types. Plant and Soil, 2015, 390(1/2): 351- 367.
[15] Jiang Y, Zang R G, Letcher S G, Ding Y, Huang Y F, Lu X H, Huang J H, Liu W D, Zhang Z D. Associations between plant composition/diversity and the abiotic environment across six vegetation types in a biodiversity hotspot of Hainan Island, China. Plant and Soil, 2016, 403(1/2): 21- 35.
[16] 刘宏伟, 王微, 左娟, 陶建平. 中梁山石灰岩山地30种主要植物叶片性状研究. 西南师范大学学报:自然科学版, 2014, 39(9): 50- 55.
[17] 习新强, 赵玉杰, 刘玉国, 王欣, 高贤明. 黔中喀斯特山区植物功能性状的变异与关联. 植物生态学报, 2011, 35(10): 1000- 1008.
[18] 马姜明, 张秀珍, 梁士楚, 陈婷, 黄秋菊. 桂林尧山常见植物叶片性状研究. 广西师范大学学报:自然科学版, 2012, 30(1): 77- 82.
[19] 党晶晶, 赵成章, 李钰, 侯兆疆, 董小刚. 祁连山高寒草地甘肃臭草叶性状与坡向间的关系. 植物生态学报, 2015, 39(1): 23- 31.
[20] 刘旻霞, 王刚, 盛红梅. 高寒草甸阳坡-阴坡梯度上环境因子特征及其与地上生物量和物种丰富度的关系. 兰州大学学报:自然科学版, 2013, 49(1): 76- 81.
[21] 胡刚, 梁士楚, 张忠华, 谢强. 桂林岩溶石山青冈栎群落的数量分析. 生态学杂志, 2007, 26(8): 1177- 1181.
[22] 卜文圣, 臧润国, 丁易, 张俊艳, 阮云泽. 海南岛热带低地雨林群落水平植物功能性状与环境因子相关性随演替阶段的变化. 生物多样性, 2013, 21(3): 278- 287.
[23] 尚晓娜, 宋平顺, 杨锡, 何禄仁, 赵建邦, 丁永辉. 甘肃不同地域甘草有效成分含量与土壤因子关系的研究.中国农学通报, 2012, 28(28): 245- 249.
[24] Bao S D. Soil Agro-Chemistry Analysis. Beijing: China Agriculture Press, 2000.
[25] 孙娇, 赵发珠, 韩新辉, 杨改河, 白孙宝, 郝文芳. 不同林龄刺槐林土壤团聚体化学计量特征及其与土壤养分的关系. 生态学报, 2016, 36(21): 6879- 6888.
[26] 庞成庆, 秦江涛, 李辉信, 刘金花. 秸秆还田和休耕对赣东北稻田土壤养分的影响. 土壤, 2013, 45(4): 604- 609.
[27] 胡耀升, 么旭阳, 刘艳红. 长白山森林不同演替阶段比叶面积及其影响因子. 生态学报, 2015, 35(5): 1480- 1487.
[28] 朱猛, 刘蔚, 秦燕燕, 曹建军, 李会亚, 赵玉. 祁连山森林草原带坡面尺度土壤有机碳分布. 中国沙漠, 2016, 36(3): 741- 748.
[29] 朱云云, 王孝安, 王贤, 邓美皎. 坡向因子对黄土高原草地群落功能多样性的影响. 生态学报, 2016, 36(21): 6823- 6833.
[30] 汝海丽, 张海东, 焦峰, 薛超玉, 郭美丽. 黄土丘陵区微地形条件下草本群落特征与土壤水分及养分关系分析. 草地学报, 2016, 24(4): 776- 782.
[31] 盛茂银, 熊康宁, 崔高仰, 刘洋. 贵州喀斯特石漠化地区植物多样性与土壤理化性质. 生态学报, 2015, 35(2): 434- 448.
[32] 刘旻霞, 马建祖. 甘南高寒草甸植物功能性状和土壤因子对坡向的响应. 应用生态学报, 2012, 23(12): 3295- 3300.
[33] 黄端, 王冬梅, 任远, 覃云斌, 吴林川. 漓江水陆交错带植物叶性状对水淹胁迫的响应及经济谱分析. 生态学报, 2017, 37(3): 750- 759.
[34] 宝乐, 刘艳红. 东灵山地区不同森林群落叶功能性状比较. 生态学报, 2009, 29(7): 3692 - 3703.
[35] 刘旻霞, 马建祖. 阴阳坡植物功能性状与环境因子的变化特征. 水土保持研究, 2013, 20(1): 102- 106.
[36] 胡耀升, 么旭阳, 刘艳红. 长白山不同演替阶段森林植物功能性状及其与地形因子间的关系. 生态学报, 2014, 34(20): 5915- 5924.
[37] 张曦, 王振南, 陆姣云, 杨梅, 杨惠敏. 紫花苜蓿叶性状对干旱的阶段性响应. 生态学报, 2016, 36(9): 2669- 2676.
[38] 陈晨, 刘丹辉, 吴键军, 康慕谊, 张金屯, 刘全儒, 梁钰. 东灵山地区辽东栎叶性状与地形因子关系. 生态学杂志, 2015, 34(8): 2131- 3139.
[39] 高景, 徐波, 王金牛, 周海燕, 王彦星, 吴彦. 南方灌草丛典型灌木不同叶片性状的相关性及其对不同光环境的响应. 生态学杂志, 2015, 34(9): 2424- 2431.
[40] 李钰, 赵成章, 董小刚, 侯兆疆, 马小丽, 张茜. 高寒草地狼毒枝-叶性状对坡向的响应. 生态学杂志, 2013, 32(12): 3145- 3151.
[41] 杭夏子, 翁殊斐, 袁喆. 华南5种园林灌木叶性状特征及其对环境响应的研究. 西北林学院学报, 2014, 29(2): 243- 247.
[42] 朱秋莲, 邢肖毅, 张宏, 安韶山. 黄土丘陵沟壑区不同植被区土壤生态化学计量特征. 生态学报, 2013, 33(15): 4674- 4682.
[43] 盛茂银, 刘洋, 熊康宁. 中国南方喀斯特石漠化演替过程中土壤理化性质的响应. 生态学报, 2013,33(19): 6303- 6313.
[44] 刘兴诏, 周国逸, 张德强, 刘世忠, 褚国伟, 闫俊华. 南亚热带森林不同演替阶段植物与土壤中N、P的化学计量特征. 植物生态学报, 2010, 34(1): 64- 71.
[45] 朱媛君, 杨劼, 万俊华, 张璞进, 牛明丽, 赵利清, 清华. 毛乌素沙地丘间低地主要植物叶片性状及其相互关系. 中国沙漠, 2015, 35(6): 1496- 1504.
[46] Vergutz L, Manzoni S, Porporato A, Novais R F, Jackson R B. Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants. Ecological Monographs, 2012, 82(2): 205- 220.
[47] 刘金根, 薛建辉. 坡向对香根草护坡地植物群落特征的影响. 生态学杂志, 2009, 28(3): 384- 388.
[48] 张川, 张伟, 陈洪松, 聂云鹏, 叶莹莹, 王克林. 喀斯特典型坡地旱季表层土壤水分时空变异性. 生态学报, 2015, 35(19): 6326- 6334.
[49] 夏宣宣, 张淑勇, 张光灿, 方立东, 张松松, 李辉. 黄土丘陵区土壤水分对黄刺玫叶片光响应特征参数的影响. 生态学报, 2016, 36(16): 5142- 5149.
[50] 宋彦涛, 周道玮, 王平, 李强. 松嫩草地66种草本植物叶片性状特征. 生态学报, 2013, 33(1): 79- 88.
[51] 龙文兴, 臧润国, 丁易. 海南岛霸王岭热带山地常绿林和热带山顶矮林群落特征. 生物多样性, 2011, 19(5): 558- 566.
[52] 许洺山, 赵延涛, 杨晓东, 史青茹, 周刘丽, 张晴晴, Arshad A, 阎恩荣. 浙江天童木本植物叶片性状空间变异的地统计学分析. 植物生态学报, 2016, 40(1): 48- 59.
