不同天气水稻光合日变化对大气CO2浓度和温度升高的响应
——FACE研究
2018-04-19袁嫚嫚朱建国王伟露
袁嫚嫚,朱建国,刘 钢,王伟露,3
1 中国科学院南京土壤研究所,土壤与农业可持续发展国家重点实验室,南京 210008 2 安徽省农业科学院土壤肥料研究所,安徽省养分循环与资源环境省级实验室,合肥 230031 3 中国科学院大学,北京 100049
工业革命以来,人类活动通过燃烧化石燃料和汽车、航天器的尾气排放等途径在大气中排放了大量温室气体和气溶胶,造成大气二氧化碳(CO2)浓度持续和温度同时持续升高,光合有效辐射(PAR)下降。根据不同模型预测,到2050年大气CO2浓度升高到470—570μmol /mol,温度上升0.8—3.2℃[1]。Wild[2]的综述认为,从1950—1980年是全球暗化时期,即PAR在全球范围普遍下降,平均每10年下降0.8%—7%,但1980—2000年,全球多数地区PAR开始上升,但远未弥补长时期的下降。而Che等[3]研究认为中国从1950—2000年PAR平均每10年下降了1.3%,近年的研究表明中国华北平原和长江中下游等地区的PAR仍在下降中[4- 6]。
水稻是光温敏感作物,CO2浓度、温度和PAR变化势必影响水稻光合作用。CO2浓度升高加速了水稻生育进程,导致叶片光合速率增加,气孔导度、蒸腾速率减小;CO2浓度和温度增加的幅度和方式影响着水稻光合作用结果[7- 8]。Figueiredo等[9]利用开顶式气室控制CO2浓度和温度,与ambient比较,高CO2浓度对水稻生产力的促进作用能够弥补增温对生产力的损伤。而Cai等[10]利用放空气CO2浓度升高系统(FACE),采用红外加热的方式增温,结果表明与CO2浓度增加效应相反,增温大大削弱了水稻的生产力,二者共同作用,水稻产量仍显著下降。不同试验模拟平台上,高CO2浓度和高温对水稻生产力影响的结果不一致,而从光合作用角度解析作物生产力变化的原因有待进一步研究。谢立勇等[11]利用FACE研究表明,高CO2浓度提高了北方粳稻光合能力,以覆盖地膜的方式增温对水稻光合作用影响不明显,这种增温方式仅能影响水稻根部区域,增温效果有限。2013年课题组为了模拟CO2浓度和温度升高的流动大气环境,在江苏江都原有的FACE系统基础上进行了提升,建立了同时升高水稻冠层空气CO2浓度和温度的新试验平台(T-FACE)[12]。周宁等[13]基于此平台研究发现,增温对水稻光合日变化的影响因生育期不同而异,CO2浓度对水稻光合作用的影响明显大于增温,而不同天气对其光合特性的影响则未见研究。不同天气PAR相差较大,PAR是太阳辐射中波长为400—700nm,是能被植物利用进行光合作用的唯一能量来源[14],晴天和阴天通过PAR的强弱影响着水稻光合作用。因此,本试验以目前生产上大面积种植的高产优质粳稻“南粳9108”为试验材料,利用T-FACE稻田开放系统模拟21世纪中叶大气环境,研究CO2浓度、温度和天气对水稻剑叶光合作用日变化的影响,以期为评估和应对气候变化对水稻生产的影响提供依据。
1 材料与方法
1.1 试验地点与平台
试验于2015年在中国稻田T-FACE研究技术平台上进行。平台位于江苏省扬州市江都区小纪镇良种场试验田(119°42′0″E,32°35′5″N)。试验田所在地区年均降水量980mm 左右,年均蒸发量大于1100mm,年平均温度约14.9℃,年日照时间大于2100h,年平均无霜期220d,耕作方式为水稻-冬小麦轮作。
试验田土壤类型为砂壤土,土壤理化性质为:有机质18.4g/kg,全氮1.45g/kg,全磷0.63g/kg,全钾14.0g/kg,速效磷10.1mg/kg,速效钾70.5mg/kg,容重1.16g/cm3,pH7.2。
平台共有3个FACE试验圈和3个对照(Ambient)圈。FACE圈设计为正六角形,直径12m,平台运行时通过FACE圈周围的管道向中心喷射纯CO2气体,并在FACE和Ambient 圈中特定位置加装热水增温管道,以热辐射形式向增温区域进行增温处理,CO2放气管的高度距水稻冠层为50cm左右,增温管道约5—10cm,增温管道高度距水稻冠层为20cm左右。利用计算机网络对平台CO2浓度和水稻冠层温度进行监测和控制,根据大气中的CO2浓度、风向、风速、作物冠层高度的CO2浓度和温度自动调节CO2气体的释放速度和方向以及增温管道中热水流速,使水稻主要生育期FACE圈内CO2浓度保持比大气高200μmol/mol,所有圈内增温区域水稻冠层空气温度比大气环境温度高1—2℃。FACE圈之间以及FACE圈与对照圈之间的间隔大于90m,以减少CO2释放对其他圈的影响。对照田块没有安装FACE管道,所有田块非增温区域没有安装热水增温管道,其余环境条件与自然状态一致[12,15]。为监测大气增温幅度,在FACE和Ambient 圈增温与非增温区域,距水稻冠层20cm处安装SI- 111红外温度(Campbell公司,美国)实时记录气温,每1min记录一次。
平台CO2熏气时间为6月25日至10月19日,增温时间为7月1日至10月19日,FACE圈每日熏气和增温处理时间为日出至日落。
1.2 试验材料
供试水稻品种为南粳9108,大田旱育秧,5月20日播种,6月17日移栽,种植密度为24穴/m2,每穴2苗,秧苗均为1蘖苗。采用复合肥(有效成分N∶P2O5∶K2O=15%∶15%∶15%)和尿素(含N 46.7%)配合施用。施N总量为22.5g/m,其中基肥占40%(6月17日)施用;30%作分蘖肥施用(7月1日),30%作穗肥施用(7月26日)。施P、K 量均为9g/m,作基肥施用。水分管理:6月17日至7月20日保持水层(约3cm),7月21日至8月10日多次轻搁田(自然落干后保持3天→灌水1天→干旱4天→灌水1天,如此4天干旱1天灌水反复),8月11日至收获前10天间隙灌溉(3天保水2天干旱),之后断水至10月26日收获。其它管理亦按高产田标准和要求执行。
1.3 试验处理
本试验为裂区设计,主区为CO2处理,设大气背景CO2浓度(AC,ambient CO2,约390μmol/mol)和高CO2浓度(EC,elevated CO2,约590μmol/mol,比大气背景CO2浓度高200μmol/mol)2个水平。裂区为温度处理,设大气环境温度(AT,ambient temperature) 和高温(ET,elevated temperature,比环境温度高1—2℃)2个水平。即4个处理,分别为环境CO2浓度和温度处理(ACAT)、环境CO2浓度和高温处理(ACET)、高CO2浓度和环境温度处理(ECAT)、高CO2浓度和高温处理(ECET)。
1.4 测定方法
采用LI- 6400 便携式光合系统分析仪(LI-COR,USA)测定水稻剑叶净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr),并根据净光合速率与蒸腾速率的比值计算叶片水分利用率(WUE)。
分别选择阴天和晴朗天气,于8月30日(阴天,相当于齐穗期),9月16日(晴天,相当于灌浆中期),从9: 00—17: 00每隔1h测定一次。测定剑叶叶片顶端的1/2至1/3处的上表面,每个处理连续测定3片有代表性的剑叶取平均值。采用LI-COR注入系统控制CO2浓度,Ambient圈和FACE圈CO2浓度分别设定为390μmol/mol和590μmol/mol。选用红蓝光源,依据测定时自然光源设置光强,温度设定取决于当时的作物冠层温度。
1.5 数据处理与统计分析
应用Excel 2007进行数据处理和图表绘制。统计分析软件采用一般线性模型,以SPSS 17.0进行方差分析,采用Duncan法进行多重比较,显著水平设P<0.01(极显著)、P<0.05(显著)和P≥0.05(无显著差异),分别用**、*和ns表示,相关性分析采用Pearson双侧显著检验。
2 结果与分析
2.1 阴天与晴天天气条件比较
从图1可以看出,不同天气条件下大气温度和PAR变化规律不同。
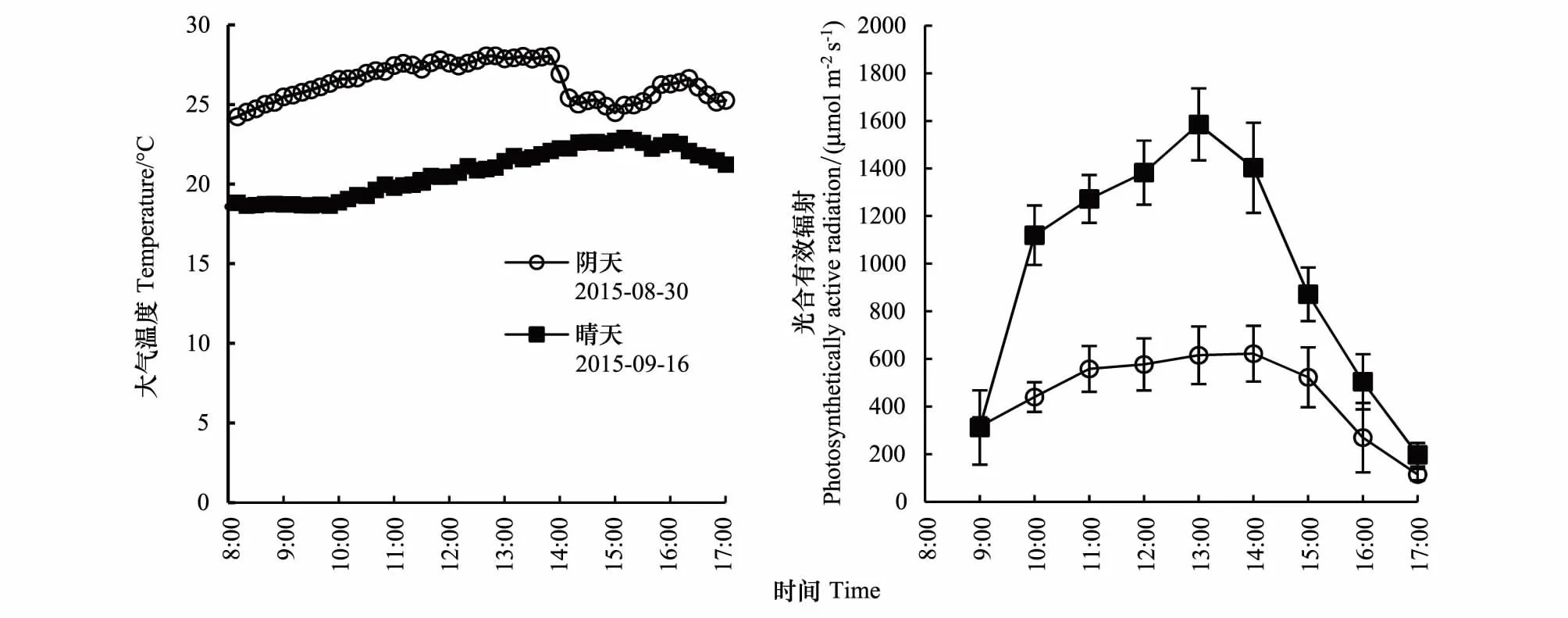
图1 不同天气下大气温度和光合有效辐射的日变化Fig.1 Diurnal variation of temperature and photosynthetic active radiation between cloudy and sunny days
8:00—17:00,阴天大气温度为24.05℃—28.06℃,平均为26.34℃,在13:00左右达到高峰;晴天大气温度为18.59℃—22.91℃,平均为20.74℃,在15:00左右达到峰值。
阴天比晴天降低了PAR。9:00—17:00,阴天PAR随时间变化比较平缓,其值为114.3—621.5mol m-2s-1,均值为448.3mol m-2s-1;晴天PAR变为单峰曲线,范围为197.5—1585.3mol m-2s-1,均值为960.8mol m-2s-1,在13:00左右达到峰值。
从表1可以看出,利用红外线测温仪测定,CO2浓度升高200μmol/mol导致大气温度升高,数据显示阴天和晴天分别使水稻冠层气温平均增加了0.30℃和0.39℃。阴天和晴天增温设施对水稻冠层大气增温幅度相近。环境CO2浓度的增温处理的阴天和晴天分别平均增温1.48℃和1.15℃,水稻在高温条件下,蒸腾作用提高,导致冠层温度下降,因此,这个增温效应是热水管道增温和蒸腾降温的综合效应;CO2浓度升高的增温处理阴天和晴天分别平均增温0.92℃和1.04℃,一方面CO2浓度升高,温度升高,一方面管道增温,蒸腾降温,因此,这个增温效果是CO2增温、管道增温、蒸腾降温的综合效应。从整体看,整体增温幅度在0.92℃—1.48℃之间,变幅不是太大,达到了试验设计使水稻冠层气温增高1—2℃的目标。
2.2 CO2浓度和温度升高的不同天气下水稻光合作用的日变化特征
2.2.1CO2浓度和温度升高的不同天气下水稻净光合速的日变化特征
由图2可知,CO2浓度升高显著增加了水稻净光合速率(Pn),而温度升高呈相反趋势,仅在阴天的13:00、14:00、16:00和晴天的12:00达到显著水平,CO2浓度和温度同时升高对水稻剑叶Pn的交互作用不明显。
晴天比阴天提高了水稻Pn对CO2浓度升高响应的幅度。与环境CO2浓度和高温处理(ACET)比较,晴天水稻Pn均值分别比高CO2浓度和环境温度处理(ECAT)和高CO2浓度和高温处理(ECET)分别增加了58.4%和56.7%,阴天分别增加了38.9%和24.7%(表2)。

表1 不同天气下增温设施的增温效果/℃
ACET—ACAT:环境CO2浓度和高温处理—环境CO2浓度和温度处理 ambient CO2and elevated temperature—ambient CO2and ambient temperature;ECET—ACAT:高CO2浓度和高温处理—环境CO2浓度和温度处理 elevated CO2and elevated temperature—ambient CO2and ambient temperature;ECAT—ACAT:高CO2浓度和环境温度处理—环境CO2浓度和温度处理 elevated CO2and ambient temperature—ambient CO2and ambient temperature;ECET—ECAT:高CO2浓度和高温处理—高CO2浓度和环境温度处理 elevated CO2and ambient temperature—elevated CO2and ambient temperature
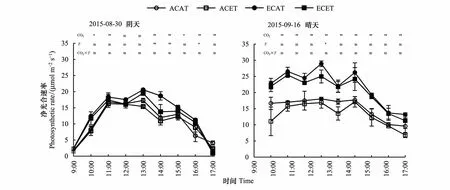
图2 CO2浓度和温度升高的不同天气下水稻净光合速率的日变化Fig.2 Diurnal variation of rice net photosynthetic rat of elevated CO2 concentration and temperature between cloudy and sunny daysT:温度 temperature;ACAT:环境CO2浓度和温度处理 ambient CO2 and ambient temperature;ACET:环境CO2浓度和高温处理 ambient CO2 and elevated temperature;ECAT:高CO2浓度和环境温度处理 elevated CO2 and ambient temperature;ECET:高CO2浓度和高温处理 elevated CO2 and elevated temperature。图中数据为平均值±标准差(n=3),ns,不显著,*和** 分别表示在P<0.05和P<0.01水平上差异显著
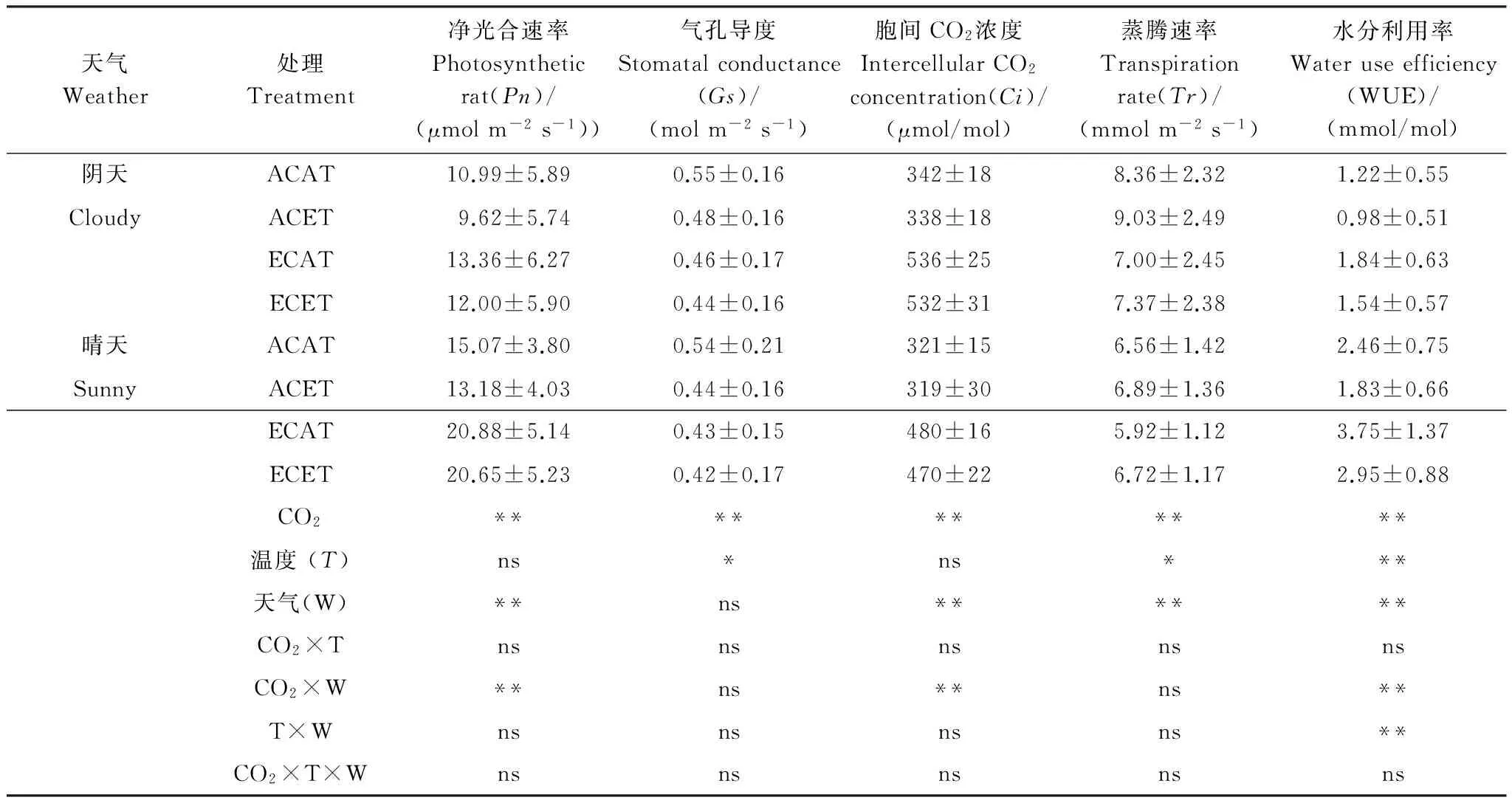
天气Weather处理Treatment净光合速率Photosyntheticrat(Pn)/(μmolm-2s-1))气孔导度Stomatalconductance(Gs)/(molm-2s-1)胞间CO2浓度IntercellularCO2concentration(Ci)/(μmol/mol)蒸腾速率Transpirationrate(Tr)/(mmolm-2s-1)水分利用率Wateruseefficiency(WUE)/(mmol/mol)阴天ACAT10.99±5.890.55±0.16342±188.36±2.321.22±0.55CloudyACET9.62±5.740.48±0.16338±189.03±2.490.98±0.51ECAT13.36±6.270.46±0.17536±257.00±2.451.84±0.63ECET12.00±5.900.44±0.16532±317.37±2.381.54±0.57晴天ACAT15.07±3.800.54±0.21321±156.56±1.422.46±0.75SunnyACET13.18±4.030.44±0.16319±306.89±1.361.83±0.66ECAT20.88±5.140.43±0.15480±165.92±1.123.75±1.37ECET20.65±5.230.42±0.17470±226.72±1.172.95±0.88CO2**********温度(T)ns*ns***天气(W)**ns******CO2×TnsnsnsnsnsCO2×W**ns**ns**T×Wnsnsnsns**CO2×T×Wnsnsnsnsns
图中数据为平均值±标准差(n=27);ns,差异不显著;*和*** 分别表示在P<0.05和P<0.01水平上差异显著
不同处理,水稻Pn阴天为单峰曲线,13:00达到峰值,与PAR显著相关;晴天为双峰曲线,分别于12:00和14:00出现两个峰值,且12:00出现全天Pn最大值,在12:00与14:00之间出现了光合“午休”。同一处理,晴天水稻Pn显著高于阴天的,ACAT、ACET、ECAT和ECET晴天水稻Pn均值比阴天的分别增加了37.1%、40.0%、56.3%和72.0%。
2.2.2CO2浓度和温度升高的不同天气下水稻气孔导度的日变化特征
不同天气下,CO2浓度和温度升高均不同程度降低了水稻气孔导(Gs)(图3),其响应程度经统计分析,所测的9h日变化时间中阴天和晴天分别有33.3%和44.4%达到显著水平,二者同时升高对水稻Gs的交互作用不明显。
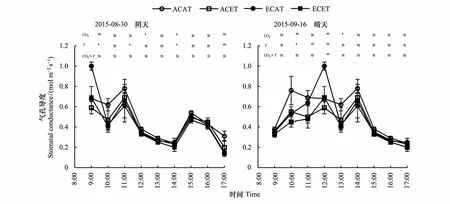
图3 CO2浓度和温度升高的不同天气下水稻气孔导度的日变化Fig.3 Diurnal variation of rice stomatal conductance of elevated CO2 concentration and temperature between cloudy and sunny days
由表2知,不同天气下水稻Gs度均以ACAT最高,阴天和晴天分别比ACET、ECAT和ECET平均提高了13.8%、19.7%、26.0%和21.7%、27.0%、29.5%,表明水稻晴天对高CO2浓度和高温的响应程度大于阴天。
阴天水稻Gs日变化趋势比较复杂,各处理的Gs均值由9:00的0.74mol m-2s-1下降到10:00的0.48mol m-2s-1,继而上升到11:00的0.69mol m-2s-1,再下降到14:00的0.23mol m-2s-1,然后有所提高又下降到17:00的0.20mol m-2s-1。晴天水稻Gs变化呈双峰曲线,峰值出现的时间与Pn一致。
2.2.3CO2浓度和温度升高的不同天气下水稻胞间CO2浓度的日变化特征
从图4可以看出,不同天气下,CO2浓度升高显著增加了水稻胞间CO2浓度(Ci),温度升高表现为抑制趋势。经统计分析,与增温效应相似,高CO2浓度和高温交互作用对水稻Ci影响仅在11:00和12:00达到显著差异,其他时间差异不显著。
水稻Ci受环境影响较大,FACE圈水稻剑叶Ci明显高于Ambient圈,而同一圈中不同处理水稻Ci日变化幅度不大。不同天气下,相同处理阴天的水稻Ci日变化均值显著高于晴天,ACAT、ACET、ECAT和ECET分别增加了6.8%、6.0%、11.7%和13.4%(表2)。
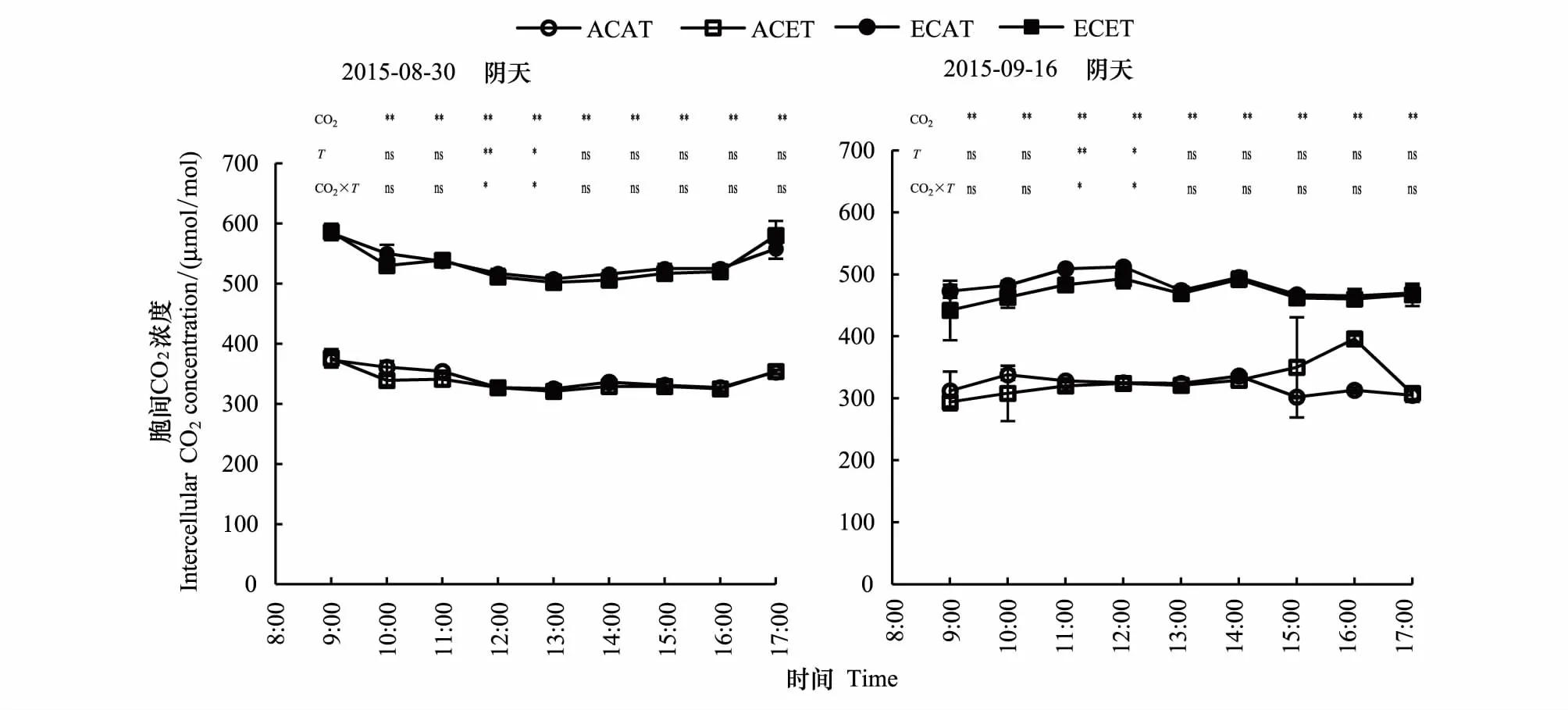
图4 CO2浓度和温度升高的不同天气下水稻胞间CO2浓度的日变化Fig.4 Diurnal variation of rice intercellular CO2 concentration of elevated CO2 concentration and temperature between cloudy and sunny days
2.2.4CO2浓度和温度升高的不同天气下水稻蒸腾速率的日变化
由图5可以看出,CO2浓度和温度升高对不同天气下水稻蒸腾速率(Tr)日变化的影响程度不一致,前者表现为抑制趋势,后者表现为促进趋势,二者交互作用不明显。由表2可以知,从均值上比较,CO2浓度升高对水稻Tr日变化的抑制作用达到极显著水平,降幅为10.8%—29.0%,温度升高的促进作用达到显著水平,增幅为5.0%—13.5%。
不同天气条件水稻Tr差异极显著。水稻Tr日变化为单峰曲线,阴天和晴天分别于13:00和14:00左右达到峰值,阴天的日变化的峰值和变幅度均大于晴天的。阴天ACAT、ACET、ECAT和ECET水稻剑叶Tr的日变化均值分别比晴天增加了27.4%、31.2%、18.3%和9.8%,平均增加了20.9%。
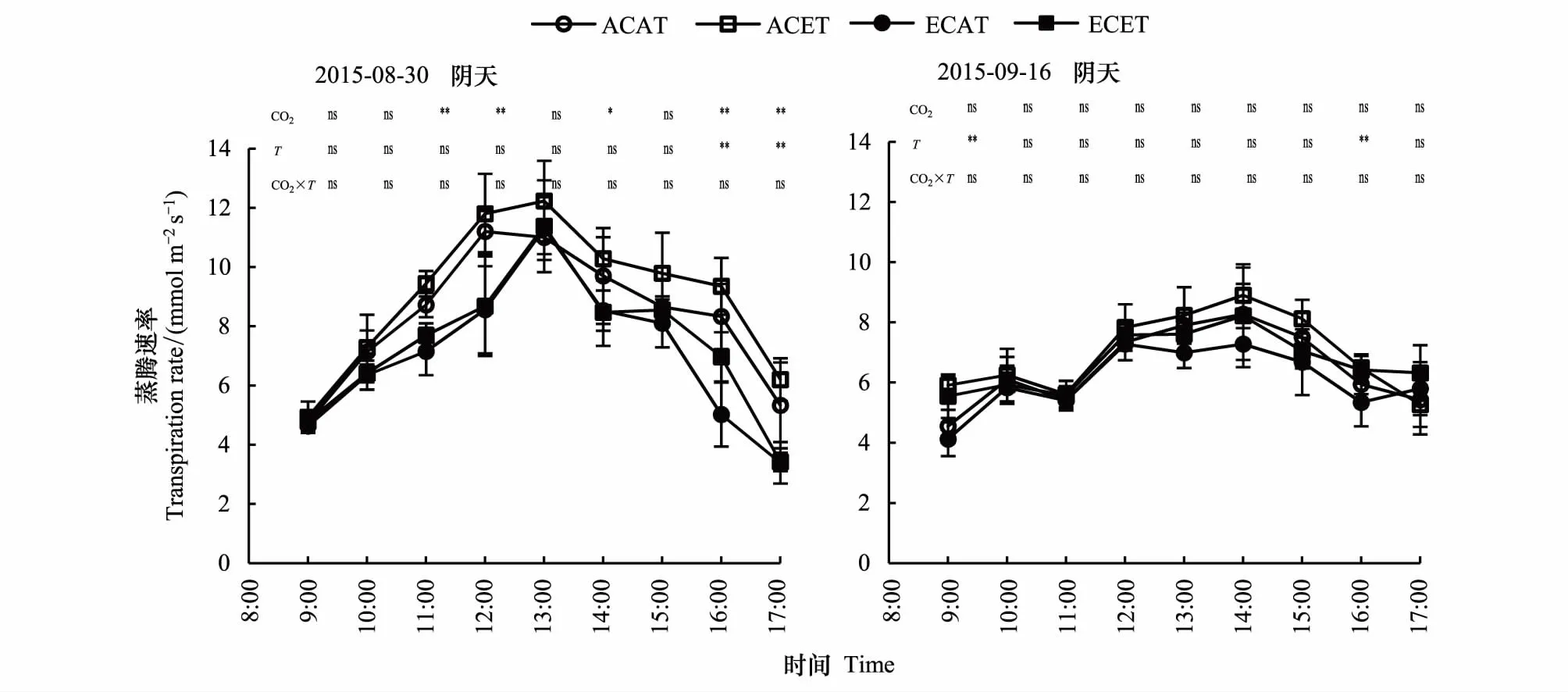
图5 CO2浓度和温度升高的不同天气下水稻蒸腾速率的日变化Fig.5 Diurnal variation of rice transpiration rate of elevated CO2 concentration and temperature between cloudy and sunny days
2.2.5CO2浓度和温度升高的不同天气下水稻水分利用率的日变化
由图6可知,CO2浓度升高显著提高了不同天气下水稻水分利用率(WUE),增温降低了不同天气下水稻WUE,二者交互作用不明显。不同处理,水稻WUE日变化均值对CO2浓度和温度的响应均达到显著水平。阴天水稻WUE 的ECAT比ACAT、ACET和ECET提高了50.9%、87.1%、19.7%,晴天提高了51.7%、96.3%、23.3%(表2)。
CO2浓度和温度升高,水稻WUE阴天的日变化表现为随时间变化先上升后下降的趋势,而晴天表现为下降的趋势。同一处理,晴天水稻WUE日变化均值显著高于阴天的,ACAT、ACET、ECAT和ECET均值分别增加了101.5%、85.5%、104.1%和92.2%。
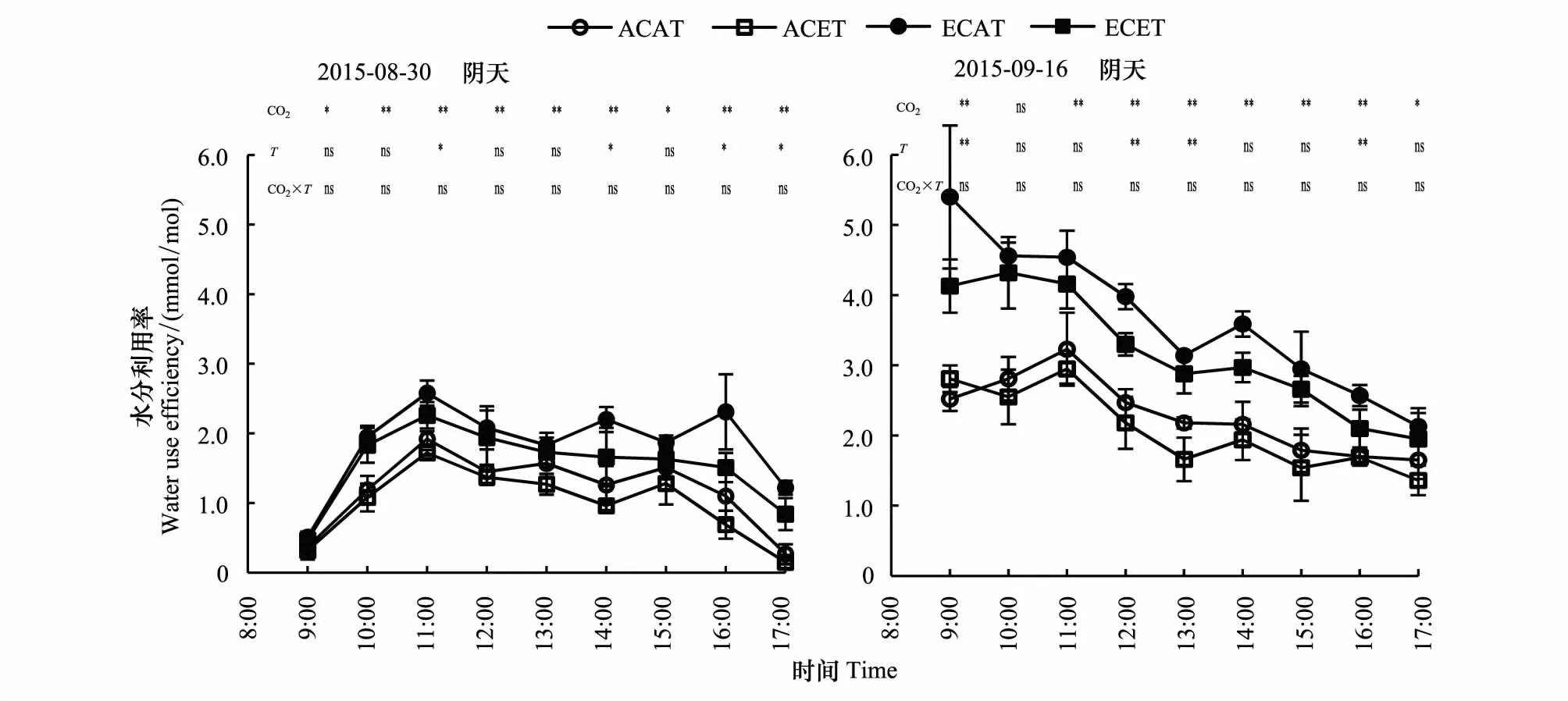
图6 CO2浓度和温度升高的不同天气下水稻水分利用率的日变化Fig.6 Diurnal variation of rice water use efficiency of elevated CO2 concentration and temperature between cloudy and sunny days
2.3 CO2浓度和温度升高的不同天气下水稻光合特征参数的相关性
不同环境因素对水稻光合特征的影响差异明显,具体表现在与水稻光合特征参数相关性不同,光合特征参数间的相关性亦不同(表3)。天气状态(W)是PAR和温度共同作用的结果,与PAR极显著相关。PAR与水稻Pn、Gs、Ci、Tr、WUE成不同程度正向相关。CO2浓度与水稻Pn、Ci、WUE向相关,而与Gs和Tr负相关。温度与Pn、Gs、Ci、WUE负相关,其中与Gs和WUE达到显著水平。
光合参数中,Pn与Gs、Ci、Tr、WUE正显著相关。Ci与Gs、Tr负显著相关。这些相关分析表明,PAR、CO2浓度和温度通过调解水稻光合参数Gs、Ci、Tr、WUE的变化,进而影响水稻Pn。
表3CO2浓度和温度升高的不同天气下水稻剑叶光合特征参数相关关系
Table3RelationshipsamongricephotosynthesischaracteristicparametersofelevatedCO2concentrationandtemperaturebetweencloudyandsunnydays
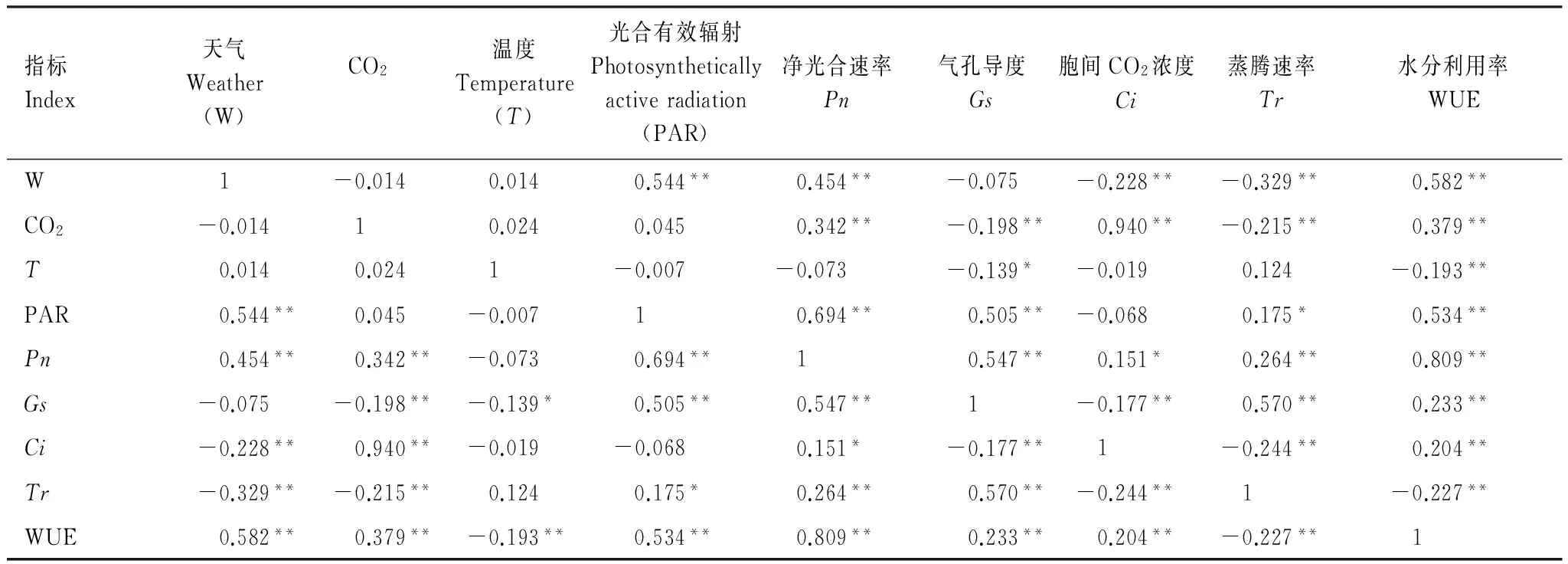
指标Index天气Weather(W)CO2温度Temperature(T)光合有效辐射Photosyntheticallyactiveradiation(PAR)净光合速率Pn气孔导度Gs胞间CO2浓度Ci蒸腾速率Tr水分利用率WUEW1-0.0140.0140.544**0.454**-0.075-0.228**-0.329**0.582**CO2-0.01410.0240.0450.342**-0.198**0.940**-0.215**0.379**T0.0140.0241-0.007-0.073-0.139*-0.0190.124-0.193**PAR0.544**0.045-0.00710.694**0.505**-0.0680.175*0.534**Pn0.454**0.342**-0.0730.694**10.547**0.151*0.264**0.809**Gs-0.075-0.198**-0.139*0.505**0.547**1-0.177**0.570**0.233**Ci-0.228**0.940**-0.019-0.0680.151*-0.177**1-0.244**0.204**Tr-0.329**-0.215**0.1240.175*0.264**0.570**-0.244**1-0.227**WUE0.582**0.379**-0.193**0.534**0.809**0.233**0.204**-0.227**1
自由度 Degree of freedom,df=45;*和** 分别表示在P<0.05和P<0.01水平上差异显著
3 讨论
水稻光合的测定多选择稳定的人工PAR或晴朗无云的天气条件,很少考虑阴天PAR大幅降低的情况。但江都FACE试验点2015年水稻生育期137d(6月17日至10月26日)中,晴天、多云或阴天、雨天数分别为30d、73d、34d[16],其中多云或阴天天数占水稻生育期53.3%。2015年水稻齐穗期阴、雨天多,晴天少。我们在测定了CO2响应曲线后,齐穗期已无晴天可期,就利用阴天观测了光合日变化。比较水稻阴天(齐穗期)和晴天(灌浆中期)的光合日变化对高CO2浓度响应时,发现高CO2浓度条件下水稻Pn与对照比的增幅,阴天显著低于晴天,说明较低的PAR减弱高CO2浓度对水稻光合的正效应。由于工业发展大气中气溶胶等颗粒物增加导致我国大气PAR不断下降[3- 6],已经严重影响到水稻光合作用。为更好地评估未来情景下水稻光合生产力变化,不同天气条件下水稻光合日变化对高CO2浓度和高温的响应差异值得进一步研究。
本研究表明,水稻阴天和晴天Pn日变化分别为单峰和双峰曲线。阴天Pn峰值出现在13:00,而后随PAR的下降而降低,未发生光合“午休”;晴天Pn在12:00和14:00两个峰值间出现了明显的下降,产生了光合“午休”。周宁等[13]在本FACE平台的研究并未发现水稻的光合“午休”现象,可能与其测定间隔较长(2h)有关。对于叶片光合“午休”的机理存在不同的观点。许大全等[14]研究认为,自然条件下晴天中午过饱和光强抑制及呼吸作用的增强,是植物光合作用效率降低的主要原因。李霞等[17]研究发现,中午强光高温引起PSⅡ反应中心可逆失活影响到ATP和NADPH同化力的产生,进而发生Fv/Fm值下降,这可能是叶片Pn中午降低的主要生理原因之一。但也有研究认为[18],“午休”的发生与维持主要由气孔限制引起,非气孔限制在光合“午休”中所占的比例远低于气孔限制。本试验南粳9108剑叶根据光响应曲线数据得到其饱和PAR值在1300mol m-2s-1左右,晴天中午PAR 超过了其饱和值,导致叶片气孔关闭,Gs降低,是光合“午休”现象产生的主要原因,其机理需进一步研究。
通常水稻齐穗期或灌浆初期光合能力最强,本试验所测的阴天(齐穗期)Pn却低于晴天(灌浆中期),表明低PAR大大削弱了水稻光合能力。通过PAR与光合参数相关性的分析,PAR与Gs正显著相关,由此可以推测阴天PAR下降,导致水稻剑叶Gs关闭。气孔限制是阴天Pn下降的主要原因。阴天Pn的下降不利于水稻生产力的提高,因此,从环境改善或品种改良等方面提高阴天水稻的Pn,值得深入研究。
4 结论
水稻光合日变化特征与CO2浓度、温度、PAR等环境因素紧密相关。在本研究中,阴天因PAR比晴天低,导致了水稻Pn的下降了37.1%—72.0%,晴天水稻光合日变化发生了光合“午休”,阴天并未发生此现象。不同天气下,CO2浓度升高对水稻光合作用促进幅度不同,阴天和晴天水稻Pn分别增加了21.6%—38.8%和38.6%—58.4%,阴天降低了CO2增加对水稻光合作用的正效应;温度升高表现出抑制水稻光合作用的趋势,但未改变CO2浓度对水稻光合作用的促进。Gs在水稻光合作用发挥着重要调解作用,当CO2浓度增加、温度升高或PAR大于水稻饱和光强时,水稻通过Gs降低以适应环境变化。
参考文献(References):
[1]IPCC (Intergovernmental Panel on Climate Change). Climate change 2007: the Physical science basis ∥ Solomon S, Qin D, Manning M, Chen Z, Marquis M, Averyt K, Tignor M, Mille H L, eds. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, United Kingdom, New York: Cambridge University Press, 2007: 996.
[2]Wild M. Global dimming and brightening: a review. Journal of Geophysical Research: Atmospheres, 2009, 114(D10): D00D16.
[3]Che H Z, Shi G Y, Zhang X Y, Arimoto R, Zhao J Q, Xu L, Wang B, Chen Z H. Analysis of 40 years of solar radiation data from China, 1961—2000. Geophysical Research Letters, 2005, 32(6): L06803.
[4]郭梁, 胡波, 戴君虎, 许建初. 近五十年北京板栗始花物候对光合有效辐射变化的响应. 植物分类与资源学报, 2014, 36(4): 523- 532.
[5]白建辉. 光合有效辐射在大气中的衰减. 环境科学学报, 2010, 30(2): 302- 313.
[6]宣守丽, 石春林, 金之庆, 曹宏鑫, 魏秀芳, 王晶晶. 长江中下游地区太阳辐射变化及其对光合有效辐射的影响. 江苏农业学报, 2012, 28(6): 1444- 1450.
[7]景立权, 赖上坤, 王云霞, 杨连新, 王余龙. 大气CO2浓度和温度互作对水稻生长发育的影响. 生态学报, 2016, 36(14): 4254- 4265.
与此同时,安全问题也不可忽视,“人-机-物”全面互联,极大地扩展了网络空间的边界和功能,也打破了工业控制系统传统的封闭格局,使得系统安全问题大量暴露出来。企业的数据重要性毋庸置疑,因此,针对安全问题的产品已经越来越多,从硬件到软件都有很好的保护措施。
[8]Wang D R, Bunce J A, Tomecek M B, Gealy D, Mcclung A, Mccouch S R, Ziska L H. Evidence for divergence of response inIndica,Japonica, and wild rice to high CO2× temperature interaction. Global Change Biology, 2016, 22(7): 2620- 2632.
[9]Figueiredo N, Carranca C, Trindade H, Pereira J, Goufo P, Coutinho J, Marques P, Maricato R, de Varennes A. Elevated carbon dioxide and temperature effects on rice yield, leaf greenness, and phenological stages duration. Paddy and Water Environment, 2015, 13(4): 313- 324.
[10]Cai C, Yin X Y, He S Q, Jiang W Y, Si C F, Struik P C, Luo W H, Li G, Xie Y T, Xiong Y, Pan G X. Responses of wheat and rice to factorial combinations of ambient and elevated CO2and temperature in FACE experiments. Global Change Biology, 2016, 22(2): 856- 874.
[11]谢立勇, 姜乐, 冯永祥, 赵洪亮, 王惠贞, 林而达. FACE条件下CO2浓度和温度增高对北方水稻光合作用与产量的影响研究. 中国农业大学学报, 2014, 19(3): 101- 107.
[12]赖上坤, 庄时腾, 吴艳珍, 王云霞, 朱建国, 杨连新, 王余龙. 大气CO2浓度和温度升高对超级稻生长发育的影响. 生态学杂志, 2015, 34(5): 1253- 1262.
[13]周宁, 沈士博, 景立权, 王云霞, 朱建国, 杨连新, 王余龙. 自由空气中CO2浓度和温度增高对粳稻叶片光合作用日变化的影响. 生态学杂志, 2016, 35(9): 2404- 2416.
[14]许大全, 徐宝基, 沈允钢. C3植物光合效率的日变化. 植物生理学报, 1990, 16(1): 1- 5.
[15]刘钢, 韩勇, 朱建国, 冈田益己, 中村浩史, 吉本真由美. 稻麦轮作FACE系统平台Ⅰ. 系统结构与控制. 应用生态学报, 2002, 13(10): 1253- 1258.
[16]江都历史天气预报查询. http://www.tianqihoubao.com/lishi/jiangdu.html.
[17]李霞, 焦德茂, 戴传超, 王守海, 吴爽, 李成荃. 转育PEPC基因的杂交水稻的光合生理特性. 作物学报, 2001, 27(2): 137- 143.
[18]王焘, 郑国生, 邹琦. 干旱与正常供水条件下小麦光合午休及其机理的研究. 华北农学报, 1997, 12(4): 48- 51.
[19]翁晓燕, 蒋德安. 生态因子对水稻Rubisco和光合日变化的调节. 浙江大学学报: 农业与生命科学版, 2002, 28(4): 387- 391.
[20]Matsui T, Namuco O S, Ziska L H, Horie T. Effects of high temperature and CO2concentration on spikelet sterility in indica rice. Field Crops Research, 1997, 51(3): 213- 219.
[21]Baker J T, Allen Jr L H. Effects of CO2and temperature on rice: a summary of five growing seasons. Journal of Agricultural Meteorology, 1993, 48(5): 575- 582.
[22]Chen C P, Sakai H, Tokida T, Usui Y, Nakamura H, Hasegawa T. Do the rich always become richer? Characterizing the leaf physiological response of the high-yielding rice cultivar takanari to free-air CO2enrichment. Plant & Cell Physiology, 2014, 55(2): 381- 391.
[23]张黎萍, 荆奇, 戴延波, 姜东, 曹卫星. 温度和光照强度对不同品质类型小麦旗叶光合特性和衰老的影响. 应用生态学报, 2008, 19(2): 311- 316.
