模拟不同初植密度杉木楠木混交林对碳储量的影响
2018-04-19胡靖宇刘静波王丽娟刘苑秋
田 晓,胡靖宇,刘静波,王丽娟,朱 琳,刘苑秋
(1. 江西农业大学,江西 南昌 330045;2.河北旅游职业学院,河北 承德 067000;3.承德石油高等专科学校,河北 承德 067000)
【研究意义】近年来,随着CO2等温室气体浓度的逐渐上升,导致全球气候变暖,已经成为国际社会共同关注的重大环境问题[1]。森林碳储量是陆地生态系统碳储量的重要组成部分,如何精确的测算森林生态系统的碳储量意义重大[2]。但是由于方法和数据的缺乏,目前碳储量的估算具有很大不确定性。因此,精确测算我国森林生态系统的碳储量,正确评价我国森林生态系统在全球碳平衡中的地位和作用,使我国在国际碳贸易的谈判中占主动地位尤为重要。【前人研究进展】杉木(Cunninghmmialanceolata)是我国中亚热带主要造林树种,但研究表明多代连栽导致林分可持续发展能力逐年下降[3]。经营杉木人工林,增加森林的碳储量、保持杉木人工林的长期生产力,成为杉木人工林经营中一个亟待解决的问题。楠木(Phoebebournei)是亚热带常见的阔叶树种,枝叶比较茂盛,冠幅大而厚,林下凋落物多且容易分解,能够增加土壤的养分,因此营造杉木楠木混交林能够改善土壤地力条件,达到丰产、稳定、高效的目的。【本研究切入点】本研究以江西省和福建省杉木、楠木混交林为研究对象,研究了不同立地条件和初植密度下楠木×杉木混交林在未来300年内(6个轮伐期)乔木层、土壤碳储量、生态系统碳储量以及年均固碳量变化趋势,从中筛选出最优的经营方式。【拟解决的关键问题】探索出楠木×杉木混交林可持续性经营策略,同时也为我国森林碳汇的估算提供参考和依据。
1 材料与方法
1.1 标准地设置与数据采集
研究地位于江西省吉安市青原区樟州村、遂川县大坑乡长荣村、遂川县茶盘洲乡溪口村,属于中亚热带季风湿润性气候,年平均温度15.1~18.1 ℃,无霜期277 d ,年平均日照时数1860 h,年均降雨量1511.3 mm,土壤主要是以红壤为主,pH5.6~6。
在研究区域分别设置3块20 m × 30 m的标准地,对标准地的楠木每木检尺,另外每个标准地设置3个样方(灌木样方5个,面积5 m×5 m;草本样方5个,面积1 m×1 m; 枯落物样方3个,面积0.5 m×0.5 m。该实验区灌木主要有黄瑞木(AdinandramellettiiBenth of Hook f.)、细柄阿丁枫(AltingiagracilipesHemsl)等。草本植物主要有铁芒萁(Dicranoperisdichotoma)和狗脊蕨(Woodwardiajaponica)等。对标准地内的灌木及草本植物的地上和地下部分分别进行称重、取样,带回实验室烘干,测其含水率。在每个样地挖取5个土壤剖面,按0~20,20~40 cm分层取样。环刀法测定土壤容重,开氏消煮法测定全氮,重铬酸钾容量法-外加热法测定土壤有机质。调查和实测数据如表1~3所示。
1.2 FORECAST模型简介
将4种数据(土壤养分、林木、林下植被、苔藓)输入到FORECAST模型中,养分可以通过动态的模拟机制进行反馈,便于经营者能够及时的采取适当的经营策略保证立地质量,并且能够获取最大碳储量[5-7]。FORECAST模型最大优点是保留了传统的经典的产量表与复杂的过程模型2种方法的优势,对未来森林的增长趋势的模拟是动态的[8-9]。例如,随时间的推移土壤肥力的下降、气候的变化、病虫害、甚至火灾等环境因素的变化都会在FORECAST模型的模拟中通过反馈机制体现[10-11]。
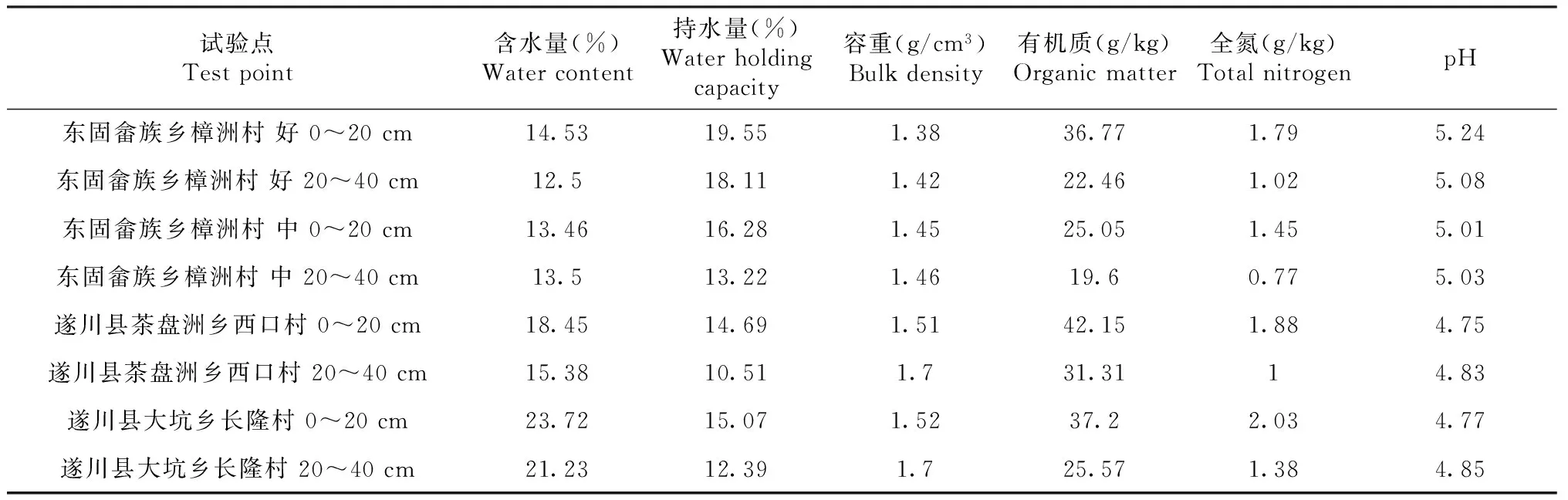
表2 楠木人工林标准地土壤理化性质
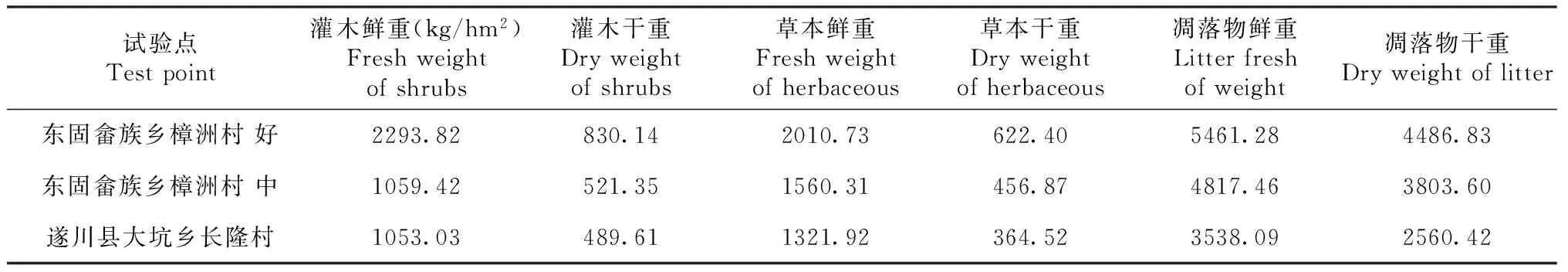
表3 楠木人工林标准地林下植被和凋落物的生物量
FORECAST模型认为,决定森林生态系统碳储量大小有2个因素,一是森林生态系统叶片总量,二是森林生态系统光合效率的大小[12-13]。光合效率大小由该树种叶片中的氮素含量决定,而氮素含量与森林系统物质积累、养分循环状况密切相关,所以决定森林生态系统物质积累、养分循环的综合性指标是氮素含量。因此,FORECAST模型选择了叶氮同化率(FNE)作为它的驱动机制[14]。在实际林分中,由于下层林冠受到上层林冠的遮荫,因此FNE需要修正,修正后叶氮同化率称为遮荫叶氮同化率(SCFNE)[15-16]。遮荫叶氮同化率计算原理是先将生态系统的林冠层沿垂直方向按固定距离划分为若干亚层, 分别计算每个亚层的相对光合效率和叶氮量,最后汇总求和[17-18]。
1.3 情景模拟
为了评估不同立地条件和初植密度下楠木×杉木混交林碳储量变化趋势,将林分的立地指数根据基准年龄的优势木高分别设置为27,21,17。将楠木×杉木混交林初植密度分别设为4000,3000,2500,2000,1600 株/hm2,杉木×楠木混交比例为1︰3。模拟时间300年,每50年1个轮伐期(共6个轮伐期),每个轮伐期内的第10年对混交林进行比例为30 %的间伐,每一个轮伐期内第30年收获杉木, 第50年收获楠木。
1.4 模型的精度检验
模型精度验证所采用的方式主要是将模型模拟数据的数据和实测数据进行比较,不同的学者从不同的角度对楠木×杉木混交林进行了大量的研究,但是关于不同管理措施和立地条件下对楠木×杉木混交林生态系统的碳储量和生物量影响的数据还是比较少,尽可能的收集所有文献中所提到混交林碳储量的数据与模拟数据进行比较。本文所收集的实测数据[19-27]在第一轮伐期内、在好中差3种立地条件下与模拟数据趋势一致(图1)。
2 结果与分析
2.1 碳储总量差异分析
如图2所示,不同初植密度楠木×杉木混交林在好中差3种立地条件下全树和茎杆碳储量由高到低依次为为较好立地、中等立地、较差立地。在较好立地条件下,初植密度为2500株/hm2的林分所积累的碳储量最高,其次是楠木×杉木混交林初植密度为2000、3000、1600、4000株/hm2。在较好立地条件下,初植密度为4000株/hm2楠木×杉木混交林不利于林分碳储量的积累,原因在于:在楠木×杉木混交林中,楠木初期生长速度要比杉木慢,初植密度过大杉木在初期会抑制混交林中楠木的生长。而混交林的密度控制在2500株/hm2能够获得最大碳储量是因为:楠木幼苗稍耐荫,杉木的速生能够在一定程度上为楠木幼苗起到了遮蔽的作用。在中等立地条件下,300年间所积累的全树碳储量和茎干碳储量从高到低以此为3000、2500、4000、2000、1600株/hm2。因为中等立地条件下单体林木的胸径和冠幅要小于较好立地条件下的胸径和冠幅,林分中单体林木之间的竞争强度相对较小,初植密度为2500株/hm2比3000株/hm2的楠木×杉木混交林在360年间所积累的碳储量少2 t/hm2,所以为了获得最大碳储量,中等立地条件下楠木×杉木混交林的初植密度应控制在2500和3000株/hm2之间。在较差立地条件下,300年间所积累的碳储量从高到低以此为4000、3000、2500、2000、1600株/hm2。即林分密度越大碳储量越高。原因在于:立地条件较差,单体林木受到土壤的养分、水分以及阳光等条件的制约,林木的个体材积和冠幅相对较小,林分中单株林木的生长不受其他林木的束缚,即林分密度越大,单位面积的株数越多,单位面积的碳储总量也就越多。
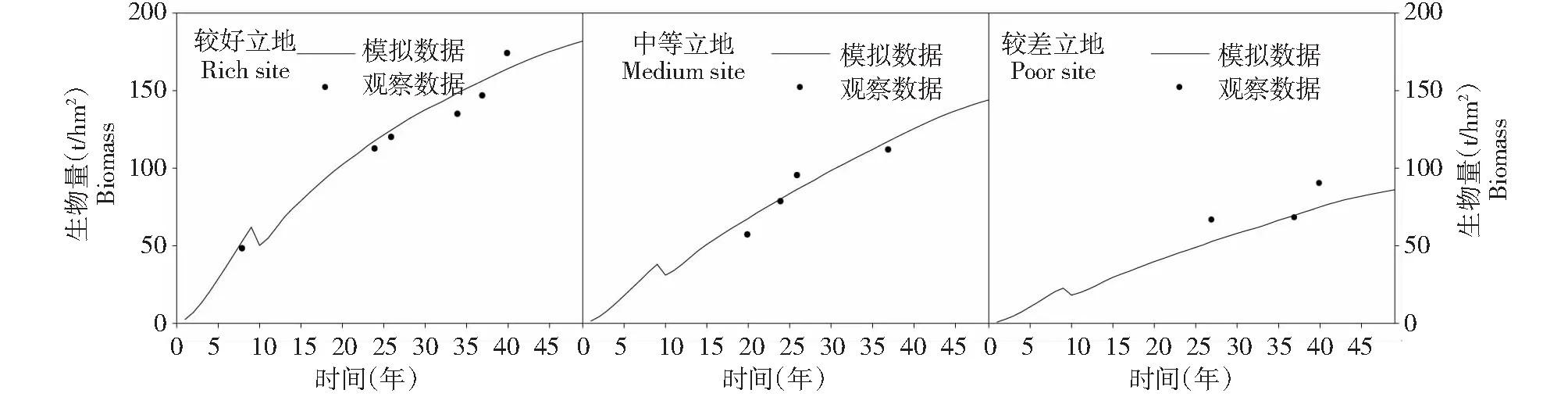
图1 模型精度验证Fig.1 Accuracy verification of model

图2 不同初植密度楠木×杉木混交林在好中差3种立地条件下全树和茎杆300年的碳储量Fig.2 Carbon storage of whole tree and stem of Phoebe bournei × Chinese fir mixed forest in 300 years under various initial planting densities in rich, medium and poor three site conditions

图3 不同立地条件下和初植密度下土壤碳储量(腐殖质+凋落物)Fig.3 Soil organic carbon storage under different site conditions and initial planting densities
2.2 土壤碳储量的变化趋势
从图3可以看出,不同立地条件和初植密度下楠木×杉木混交林在300年的模拟中土壤有机碳库均呈现上升趋势,特别是在较差立地条件下,土壤有机碳库的上升趋势更为明显,笔者用相同的方法模拟杉木纯林、杉木×楠木混交比例3︰1和杉木×楠木混交比例1︰1 3种林分发现:3种林分在模拟时间段中土壤碳储量有不同程度的下降,并且楠木比例越少土壤退化程度越严重[28],由此可见,营造楠木×杉木混交林时,适当增加楠木的比例能够有利于土壤有机碳库的积累,有利于楠木×杉木混交林的可持续经营。
2.3 生态系统碳储量变化趋势
如图4所示,不同初植密度楠木×杉木混交林在好中差3种立地条件下,生态系统碳储量由高到低依次为较好立地、中等立地和较差立地。楠木×杉木混交林生态系统碳储量在6个模拟轮伐期内呈现上升趋势,特别是在较差立地条件下,生态系统碳储量的上升趋势更为明显,由此可见,在较差立地条件下,楠木×杉木混交林对生态系统碳储量的改善作用更为明显。
2.4 净生产力的变化趋势
如图5所示,不同立地条件和初植密度下楠木×杉木混交林NPP由高到低依次为较好立地条件、中等立地条件和较差立地条件。不同初植密度的混交林NPP随着轮作次数的增加呈现上升趋势。初植密度为2500株/hm2的楠木×杉木混交林在第30年收获杉木后,剩下的楠木纯林在20年内NPP最高。不同初植密度的楠木×杉木混交林在每个轮伐期内第30年收获杉木后,剩余的楠木纯林在20年内净生产力都有不同程度的上升趋势,说明楠木30年并未达到成熟期,生物量还有一定的上升空间。

图4 不同立地条件和初植密度生态系统碳储量Fig.4 Ecosystem carbon storage under different site conditions and initial planting densities
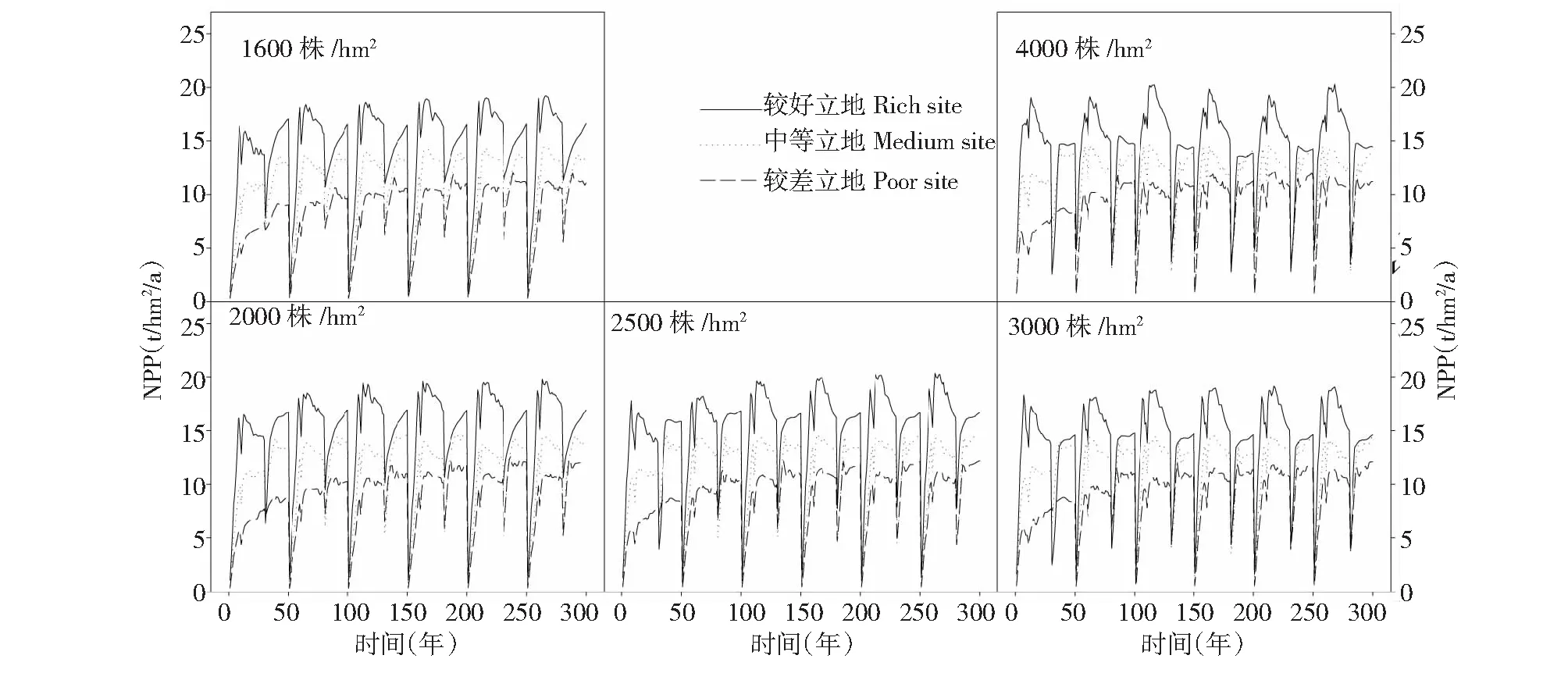
图5 不同初植密度楠木×杉木混交林在300年间净初级生产力Fig.5 Primary productivity of Phoebe bournei × Chinese fir mixed forest under various initial planting densities in 300 years

表4 年均净生产量
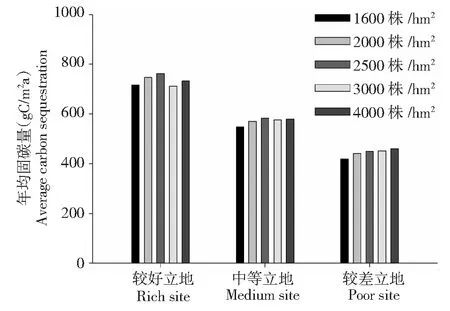
图6 不同立地条件和初植密度下年均固碳量Fig.6 Annual average fixed carbon content under different site conditions and initial planting densities
2.5 年均固碳量差异分析
如表4所示,以中等立地条件为例,5种不同初植密度的楠木×杉木混交林净生产量平均值为12.21t/(hm2·a),而相同方法模拟的楠木纯林在中等立地条件下净生产量平均值为11.47 t/(hm2·a),由此可见:楠木×杉木混交林的年均净生产量要高于相同立地条件下楠木纯林的净生产量。但略低于中亚热带森林净生产量13.5 t/(hm2·a)。在较好立地条件下,初植密度为2500株/hm2的林分年均净生产量最高,年均净生产量为15.26 t/(hm2·a),在较差立地条件下,随着林分密度的增大,年均净生产量呈现上升趋势,初植密度为4000株/hm2年均净生产量最高,年均净生产量为9.81t/(hm2·a)。从年均固碳量的角度来讲,如图6所示,以中等立地条件为例,初植密度为2500 株/hm2的混交林年均固碳量最高,为616.48 gC/(m2·a),这个结果虽然低于中亚热带平均年均固碳量675 gC/(m2·a),但却高于同处亚热带楠木年均固碳量538.38 gC/(m2·a)和杉木年均固碳量348.90 gC/(m2·a),由此可见:楠木×杉木混交林相比于楠木和杉木纯林有较高的固碳能力。
3 结 论
在300年的模拟时间里,不同初植密度多代连栽的楠木×杉木混交林在好、中、差3种立地条件下,碳储总量、土壤碳储量、生态系统碳储量以及年均固碳量都有较大的差异,表现为较好立地条件>中等立地条件>较差立地条件。并且楠木×杉木混交林随着连栽次数的增加碳储总量、土壤碳储量、生态系统碳储量以及净生产力都呈现上升趋势。在较好立地条件下,初植密度为2500株/hm2的林分300年间所积累的碳储量及年均净生产力最高;中等立地条件下楠木×杉木混交林的初植密度控制在2500和3000株/hm2之间,能够获得最大碳储量;在较差立地条件下,楠木×杉木混交林的初植密度应该控制在4000株/hm2能够获得最大碳储量积累。
4 讨 论
4.1 不同初植密度楠木×杉木混交林碳储量的差异
在较好或中等的立地条件下,初植密度为2500株/hm2的楠木×杉木混交林在6个轮伐期内所积累的碳储量最高,这是主要是因为初植密度为2500株/hm2的楠木×杉木混交林在生长初期楠木、杉木生长空间大、养分充足,可以促进单体树木高度的生长,并且杉木生长速度快,而楠木幼年较耐阴,杉木的快速生长能够为楠木起到一定的庇荫作用;其次,10年后又采取30 %的间伐,使得混交林的密度减小,又能促进混交林中树木胸径的生长,由于林隙见光照充足使得灌木草本植物的生长不受影响,有利于维持土壤的地力,从而保证混交林的碳储量的积累。以往在亚热带造林初植密度通常采取3000~3500株/hm2,也有部分国营林场采用4000株/hm2,实际上在较差的立地条件下,由于地力养分不足,单株林木的材积较小,林木间的相互影响不大,初植密度为4000株/hm2混交林有利于碳储总量的积累。在中等或较好立地条件下营造楠木×杉木混交林,由于混交林中林分树种的生长速度不同,如果林分密度过大,会导致林木的生长空间变小,林分相互竞争激烈,生长会受到一定程度的抑制,同时也会影响林下植被的生长,不利于恢复土壤的生产力;如果林分过于稀疏,虽然树木单体生长空间变大,但是混交林中林木总量过少,以及林木枝干间相互整枝作用减弱,都会影响楠木×杉木混交林的碳储总量。综上所述,在中等以上的立地条件下营造楠木×杉木混交林,初植密度为2500株/hm2能够获得最大碳储量。
4.2 FORECAST 模型在针阔混交林模拟中的应用问题
目前,FORECAST 模型在全球许多国家都开始应用,但是更多是应用于针叶纯林方面,近年来,许多学者都认识到针阔混交林无论是在保护生态环境还是在经济效益方面都有巨大优势,所以将FORECAST 模型应用于针阔混交林中意义更为重大,本研究杉木数据是借鉴加拿大著名森林生态学家J.P.(Hamish)Kimmins团队的收集的数据,在此基础上实测的楠木数据加以补充。当然模型自身也有一定的局限性。只考虑了水分的吸收,而没有考虑其对生长的影响,但目前,Kimmins团队正在将水分模型(FORWADY)与FORECAST模型相耦合[17],未来森林生态系统的预测将会更加客观、更加精确。
参考文献:
[1]方精云. 中国森林生产力及其对全球气候变化的响应[J]. 植物生态学报,2000,24(5): 513-517.
[2]冯宗炜,王效科,吴 刚. 中国森林生态系统的生物量和生产力[M]. 北京:科学出版社,1999.
[3]吴丽芝,姚云峰,王 雄,等. 赤峰市森林资源系统可持续发展能力动态评价[J]. 内蒙古大学学报,2014(2):176-183.
[4]田 晓. 基于FORECAST模型楠木人工林碳储量及NPP的研究[D]. 南昌:江西农业大学,2011.
[5]Kimmins J P. Scientific foundations for the simulation of ecosystem function and management in FORCYTE-11[R]. Northwest Region Information Report NOR-X-328,1993:4-10.
[6]Kimmins J P, Mailly D,Seely B. Modelling forest ecosystem net primary production: the hybrid simulation approach used in FORECAST[J]. Ecological Modelling, 1999,122: 195-224.
[7]Cuevas S, Vazquez F M, Torres E, et al. OptimizingCorkoakdehesa forest management using FORECAST ecosystem model[J]. SUBERWOOD,2005,10: 20-22.
[8]Seely B, Welham C, Kimmins J P. Criterion 2: Ecosystem productivity[R]. BC Journal of Ecosystems and Management, 2006, 7(1): 85-91.
[9]Blanco J A.The representation of allelopathy in ecosystem-level forest models[J]. Ecological Modelling, 2007, 209: 65-77.
[10]Blanco J A, Seely B, Welham C, et al. Testing the performance of forest ecosystem model (FORECAST) against 29 years of field data in aPseudotsugamenziesiiplantation[J]. Can.J.For.Res., 2007, 37: 1808-1820.
[11]Kimmins J P. From science to stewardship: Harnessing forest ecology in the service of society[J]. Forest Ecology and Management,2008, 258: 1625-1635.
[12]Welham C, Blanco J A, Kimmins J P. FORCEE helps to manage mixedwoods[R]. FORREX Forest Research Extension Partnership,2008.
[13]Kimmins J P. From science to stewardship: Harnessing forest ecology in the service of society[J]. Forest Ecology and Management, 2008, 258: 1625-1635.
[14]Wang S L,Chen C Y. Ecology of forest litter[M]. Science Press.2010.
[15]Fang-Fang Yang,Yue-Lin Li,Guo-Yi Zhou,et al. Dynamics of coarsewoody debris and decomposition rates in an old-growth forest in lower tropical China[J]. Forest Ecology and Management, 2010 (259): 1666-1672.
[16]Blanco J A.The representation of allelopathy in ecosystem-level forest models[J]. Ecological Modelling, 2007, 209: 65-77.
[17]Blanco J A. The legacy of forest management: analyzing its influence on tree growth with ecosystem Models[M]. Nova Science Publishers,2010.
[18]田 晓,胡靖宇,刘苑秋,等. 森林生态系统经营的新模式:FORECAST模型[J]. 林业调查规划,2010,35(6):18-25.
[19]彭龙福. 不同林分密度楠木人工林生物量初步研究[J]. 福建林业科技,2008,35(4):15-23.
[20]孙祥水. 间伐对楠木杉木混交林生长影响的研究[J]. 亚热带农业研究,2008,4(3):184-187.
[21]林亦曦. 在杉木伴生下的闽楠人工林生产力研究[J]. 福建林业科技,2007,34(4):38-41.
[22]吴载璋. 楠木杉木混交林生长效应研究[J]. 福建林学院学报,2005,25(2):142-46.
[23]邱盛樑. 楠木不同混交造林模式的生长效果比较[J]. 林业科技开发,2001,15(1):26-27.
[24]陈淑容. 不同立地因子对楠木生长的影响[J]. 福建林学院学报,2010,30(2):157-160.
[25]郑金兴,刘小飞,高 人,等. 福建南平35 a生楠木林生态系统碳库及分配[J]. 亚热带资源与环境学报,2009,4(4):59-65.
[26]马明东,江 洪,刘跃建,等. 楠木人工林生态系统生物量、碳含量、碳储量及其分布[J]. 林业科学,2008,44(3):34-39.
[27]彭龙福. 35年生楠木人工林生物量及生产力的研究[J]. 福建林学院学报,2003,23(2): 128-131.
[28]田 晓,刘苑秋,魏晓华,等. 模拟楠木杉木人工混交林不同混交比例对净生产力和碳储量的影响[J]. 江西农业大学学报,2014,36(1):122-130.
