天气条件对喀斯特森林优势树种天鹅槭树干液流的影响
2018-04-13袁丛军丁访军刘延惠崔迎春舒德远
袁丛军, 丁访军, 刘延惠, 崔迎春, 舒德远
(贵州省林业科学研究院,贵州贵阳 550005)
水分是林木进行光合、呼吸等生理代谢活动的重要物质基础和先决条件,是影响植物发育和分布的主要环境因素。如何定量估算不同林木尺度的水分利用状况一直是树木生理生态学的热点问题[1]。树木树干液流的测定是量化树木耗水和需水的基础[2],蒸腾作用是森林生态系统向大气输送水分的重要途径[3],是土壤-植物-大气连续体水分循环路径中关键的过程之一,决定着整个树冠的蒸腾耗散量[4]。由于树冠对水分利用的主要动力源于植物蒸腾拉力,因而树干液流量可表征林木蒸腾量[5],已成为研究树木耗水特性及其水分传输机制的重要指标。为了较准确地测量植物蒸腾,国内外许多植物生理学家提出了多种测量方法,刘茜从叶片尺度的角度研究了樟树(Cinnamomumcamphora)叶片蒸腾速率的日变化和季节变化特征[6];王兵等从林木个体角度研究了丝栗栲(Castanopsisfargesii)树干液流蒸腾与气象因子间的关系特征[7];Schipka等从林分角度研究地中海欧洲水青冈林冠冠层蒸腾变化特征[8],其中,Granier热消散式探针法被认为是目前测定乔木蒸腾作用最准确的方法,受到广泛关注[4,9-13]。研究表明,林木自生生长蒸腾耗散与林分所处环境因子密切相关[5,9,12-15],例如土层浅薄和间歇性干旱是限制喀斯特区植物生长与发育的典型生境特征。杨瑞等基于土壤水分差异视角,对黔中喀斯特区的石面、石沟、土面3种不同小生境生长的乡土树种云南鼠刺(Iteayunnanensis)树干液流特征进行研究[9],有关喀斯特区树种树干液流研究较少[10,16],对区域林分或群落尺度的推演缺乏足够的数据支持,开展天气条件对喀斯特森林群落优势树种树干液流影响特征变化研究意义重大。
基于荔波茂兰喀斯特生态系统定位研究站的固定样地的林木液流研究平台,观测试验设计开展监测的主要物种有天鹅槭(Acerwangchii)、丝栗栲(Castnopsisfargesii)、青冈栎(Cyclobalanopsisglauca)、云贵鹅耳枥(Carpinuspubescens)等。本研究仅以该研究区域的常绿阔叶树种——天鹅槭为研究对象,通过同步观测林木树干液流和天气条件变化特征,分析典型天气条件对天鹅槭树干液流影响特征,试图为区域的森林植物群落的常绿落叶阔叶混交林树种液流特征、水分平衡循环提供基础数据,为估测喀斯特森林水分平衡循环和喀斯特退化生态系统植被恢复与重建中树种筛选提供理论基础和依据。
1 材料与方法
1.1 研究区与样木选择
研究区位于贵州省茂兰国家级自然保护区内(25°09′~25°20′ N,107°52′~108°05′ E),平均海拔在550~850 m;保护区面积212. 85 km2,森林覆盖率高达87.3%,属中亚热带南部季风湿润气候,年均气温18.30 ℃,≥10 ℃活动积温达5 767.9 ℃,年均降水量1 320.5 mm,年均相对湿度80%,全年日照时数1 272.8 h,日照率29%[17],属裸露型喀斯特地貌,土壤以黑色石灰土为主,上层浅薄且不连续,地表水缺乏,土体持水量较低,多数地段为中亚热带原生性喀斯特森林,即常绿落叶阔叶混交林。主要优势树种有云贵鹅耳栎(Carpinuspubescens)、朴树(Celtistetrandra)、圆果化香(Platycaryalongipes)、黄连木(Pistaciachinensis)等落叶树种及青冈栎(Cyclobalanopsisglauca)、天鹅槭(Acerwangchii)、腺叶山矾(Symplocosadenopus)、黄梨木(Boniodendronminus)、卫矛(Euonymusalatus)、丝栗栲(Castnopsisfargesii)等常绿树种。
根据样地调查结果,基于观测目的和成本考虑,在固定样地周围选择3株胸径径级大小接近样地天鹅槭平均胸径的林木作为观测样木(表1),鉴于本研究区域的喀斯特小生境特征,样木与样木间具有明显的裸露岩石隔离,各株树根部相互距离5 m以上。

表1 观测样木基本情况
1.2 树干液流与气象因子测定
在2013年1月1日至2014年1月1日期间,选择生长良好、树干通直、无病虫害、径级大小适中、冠幅生长相似的3株天鹅槭作为观测样木;在样木东向距树干基部1.3 m处刮掉树皮,采用Dynamax公司生产的热扩散液流探针(TDP,Dynamax,美国)直接测量树干液流。为了防止太阳辐射、露水和降水等因素对探头及测量的影响,用夹有隔热材料的锡箔包裹探针和树干,最后用胶带密封。
将TDP馈线与数据采集器连接,接通电源,设置数据采集器工作参数,数据采集间隔期为10 min,每隔10 d下载数据。树干液流速率计算公式如下:
K=(dTM-dT)/dT。
式中:dTM为无液流中探针的最大温差,℃;dT为当时测定的温差,℃。dT通过上方和下方热电偶的电压差得到。
平均的茎流速度v(cm/s)和K之间的经验方程为v(cm/s)=0.011 9×K1.231;
茎流速度v转化成茎流率的方程如下:
Fs=As×v×3 600。
式中:Fs是茎流量(cm3/h),As是茎流传导层木材的横截面积(cm2),数据来源于12株天鹅槭的边材面积,建立方程As=0.742 6×D1.807 7(r2=0.876 2)来计算。
采用Campbell自动气象站光合有效辐射、总辐射(W/m2)、空气温度(℃)、空气湿度(%)、风速(m/s)、降水量(mm)和土壤含水量等环境因子,与液流仪同步观测;空气水汽压亏缺(简称VPD,kPa)计算方法参照文献[18-19]。研究区观测年度的主要气象因子指标见表2,典型天气主要气象因子见表3。
1.3 数据处理与分析
选取2013年典型月份代表不同季节,取冬季(1月)、春季(4月)、夏季(7月)、秋季(10月)的连续典型天气(包括不同的天气组合)进行数据分析。

表2 研究区观测年度主要气象因子特征
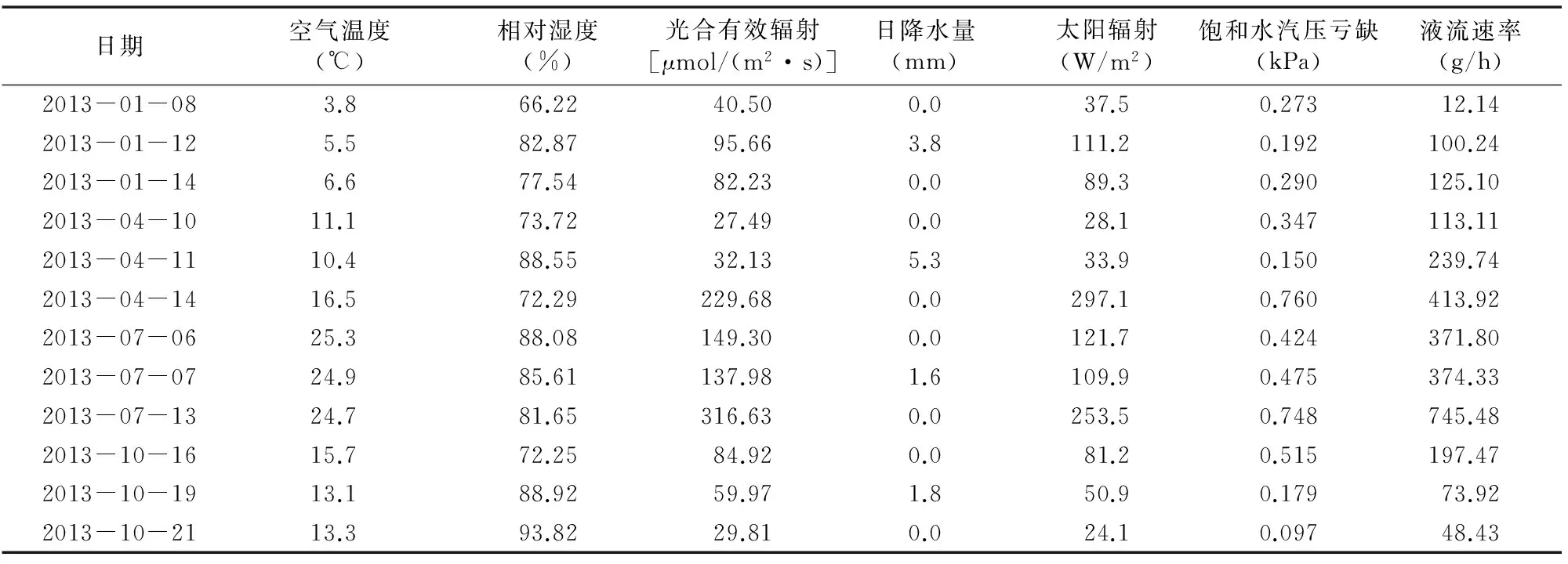
表3 典型天气主要气象因子
以不同季节典型天气下的天鹅槭树干液流日变化为基础,通过选取00:00—03:00、03:00—06:00、06:00—09:00、09:00—12:00、12:00—15:00、15:00—18:00、18:00—21:00、21:00—24:00时间段(Tt)的平均液流量(Fd,g/h)为基础,计算天鹅槭蒸腾量(transpiration,简称TSP)[20-21]:
TSP(cm3)=Fd×Tt。
本研究采用Excel 2013软件进行数据处理与制图,用SPSS 21.0统计软件对优势树种树干液流进行偏相关和回归分析。
2 结果与分析
2.1 天鹅槭树干液流变化特征
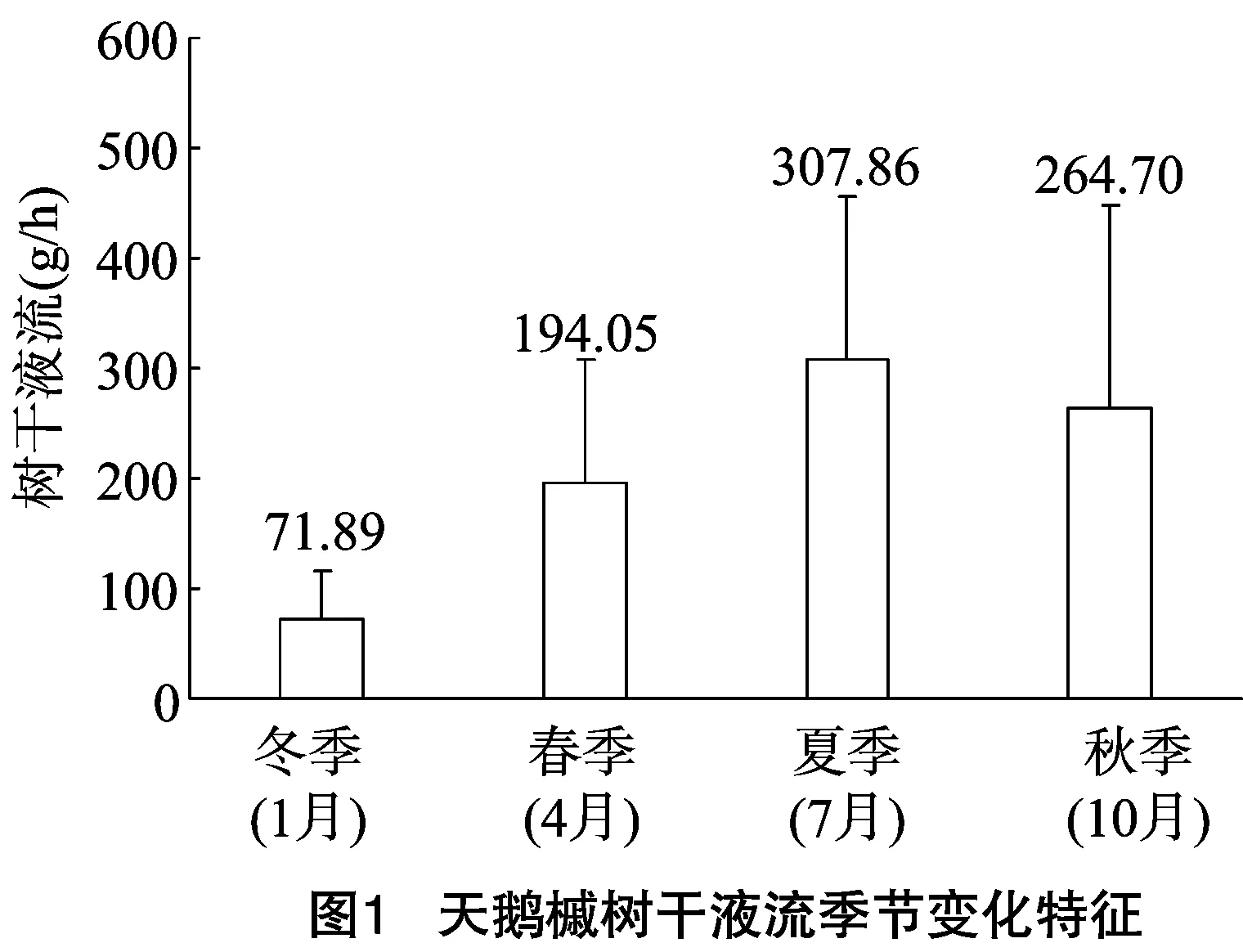
选取2013年4个不同季节时期的树干液流和环境因子数据进行分析(图1)。结果表明,各季节日均液流量差异明显,依次为夏季Fd(307.86 g/h)>秋季Fd(264.70 g/h)>春季Fd(194.05 g/h)>冬季Fd(71.89 g/h);其中,冬季平均液流量与春季、夏季、秋季均表现为明显差异,而秋季与夏季、春季间的平均液流量差异不明显。
本研究为考虑天鹅槭树干液流变化的完整性,各季节选取以降水为中心,降水前、降水后各1 d进行树干液流量日变化(图2)。总体看来,液流量呈现明显的“昼夜变化”节律性,白天液流量明显高于夜间液流量。由图2可以看出,天鹅槭的液流量日变化由于受到环境因子的影响而呈单峰波动曲线。
冬季(1月)液流量整体变化较平缓,其中降水前(1月8日)液流量日变化呈典型的单峰型曲线,液流量在12:00时取得最大值,为75.33 g/h,降水日(1月12日)和降水后(1月14日)无明显变化规律。春季(4月)液流量整体变化波动较大,其中降水前(4月10日)液流量日变化曲线峰型变化不明显,呈现多峰型,在08:00、15:00分别出现峰值,峰值液流量分别为307.00、137.44 g/h;降水日(4月11日)液流量日变化曲线呈现多峰型曲线,在06:00、09:00、13:00、15:00分别出现峰值,液流量分别为614.35、504.32、433.90、293.38 g/h;降水后(4月12日)液流量在06:00呈明显的上升趋势,并分别在13:00、15:00出现液流量的高峰,分别为1 280.07、1 380.70 g/h,这可能由于天鹅槭为适应中午天气强日照,通过叶片作出了一些水分调控策略以应变天气变化。
与冬季、春季不同的是,夏季(7月份)液流量日变化曲线均出现明显的峰,日波动变化极为明显,其中,降水前(7月6日)树干液流06:00开始启动(1.48 g/h),逐渐缓慢增大,到13:00时取得液流量峰值(1 377 g/h),随后逐渐降低,直到 19:00—23:00,仍然保留较高的液流量(134.22 g/h),说明夏季树干夜间仍然保留一定强度液流,以此来补充白天植物过度的水分消耗,弥补体内水分不足。降水日(7月7日)液流量全天均不为0,其值介于16.06~1 260.79 g/h,液流量在14:00取得峰值(1 260.79 g/h);降水后(7月13日)液流量因天气因素,呈明显的双峰型曲线,液流在6:00开始启动(2.04 g/h),随后逐渐上升,至12:00出现第1次峰值(2 328.93 g/h)后,出现缓慢降低后在16:00时出现第2次峰值(2 366.84 g/h)。
树木进入秋季(10月份),生长代谢逐渐变慢,液流量日变化波动变缓。其中,降水前(10月16日)出现明显的双峰型,分别在11:00、14:00时取得最大值,分别为471.79、534.32 g/h;降水日(10月19日)液流量在13:00、15:00取得峰值,分别为261.07、260.95 g/h;降水后(7月13日)液流量因天气因素,无明显峰型变化。
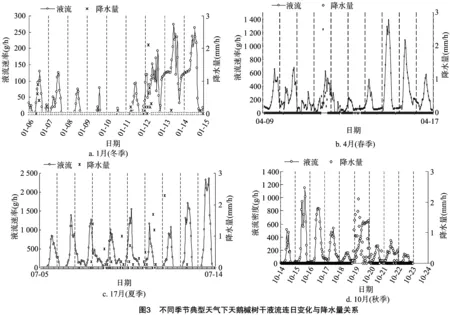
2.2 液流特征对晴-雨交替的响应变化
各季节树干液流特征对晴-雨天气交替变化的连日变化如图3所示。总体看来,冬季、秋季的天鹅槭液流连日变化曲线波动最小,日液流量峰值分别为16.87~273.13、22.65~194.49 g/h;春季天鹅槭液流量连日变化曲线波动次之,日液流量峰值为29.69~312.88 g/h;夏季天鹅槭液流量连日变化曲线波动最小,日液流量峰值为142.80~659.30 g/h。不同季节下的晴天、雨天的天鹅槭液流日变化多呈单峰曲线,而在夏季、秋季部分呈明显的双峰曲线,这可能与研究区夏秋季节特性(10月份时天气条件仍然较好)有关。

3 结论与讨论
通过研究典型季节天气条件下天鹅槭树干液流变化特征,初步取得如下结论:
天鹅槭在各季节液流差异较大,且树干液流量均呈现明显的昼高夜低现象,冬季和秋季树干液流量明显低于春季和夏季的液流量,平均液流量呈现夏季(307.86 g/h)>秋季(264.70 g/h)>春季(194.05 g/h)>冬季(71.89 g/h)。
季节和天气条件(如日降水量)明显影响了树干液流日变化曲线的波动范围和程度,各季节日夜流量峰值变化依次为夏季(142.80~659.30 g/h)、春季(29.69~312.88 g/h)、冬季(16.87~273.13 g/h)、秋季(22.65~194.49 g/h)。
水分在植物体内的状态与传输过程是土壤-植物-大气连续体(SPAC)水分传输理论的核心内容[22],树干液流(tree sap flow)作为SPAC连续体水流路径中一个关键的过程,决定整个树冠的蒸腾量,已成为分析树木耗水特性、研究树木水分传输机制的重要指标而备受关注[4]。研究显示,天鹅槭树干液流速率季节间呈显著差异,其中以夏季日均液流最大,秋季次之,冬季最小,与黄玉清等对岩溶区青冈栎树干液流季节变化结果相似[16]。树干液流日变化均呈“昼高夜低”现象,白天液流变化显著,夜间也存在液流,夜间液流变化缓慢,与马尾松(Pinusmassoniana)、锥栗(Castanopsischinensis)、木荷(Schimasuperba)和广东润楠(Machiluskwangtungensis)相似[24];日变化峰型曲线类型(单峰型、双峰型或多峰型)与季节、天气条件密切相关(图2)。在季节尺度上,冬季液流整体变化较平缓;春季液流整体变化波动较大;夏季液流日变化曲线均出现明显的峰,日波动变化极为明显;秋季生长代谢逐渐变慢,液流日变化波动变缓。
在日变化尺度上,冬季降水前(1月8日)液流日变化曲线呈典型的单峰型曲线,降水日(1月12日)和降水后(1月14日)无明显变化规律。春季降水前和降水日液流日变化曲线峰型变化均不明显,呈现多峰型,降水后(4月12日)液流在06:00呈明显的上升趋势,并分别在13:00、15:00出现液流的高峰值,分别为1 280.07、1 380.70 g/h,表明天鹅槭为了适应中午强日照天气,通过叶片做出了一些水分调控策略以应变天气变化,并有间歇“午休”现象。
夏季降水前、降水日和降水后树干液流因天气因素,呈明显的双峰型曲线。秋季降水前(10月16日)和降水日(10月19日)均出现明显的双峰型,降水后(7月13日)液流因天气因素,无明显峰型变化。可能是由于喀斯特区域的土层浅薄、植物吸收的土壤水分变化剧烈,对异质性和临时性干旱胁迫的立地条件有适应策略,其内在机制有待进一步开展对比试验研究。
参考文献:
[1]王华田. 林木耗水性研究述评[J]. 世界林业研究,2003,16(2):23-27.
[2]龙秋波,贾绍凤. 茎流计发展及应用综述[J]. 水资源与水工程学报,2012,23(4):18-23.
[3]Bernacchi C J,Vanloocke A. Terrestrial ecosystems in a changing environment:a dominant role for water[J]. Annual Review of Plant Biology,2015,66(51):599-622.
[4]赵春彦,司建华,冯起,等. 树干液流研究进展与展望[J]. 西北林学院学报,2015,30(5):98-105.
[5]张雷,孙鹏森,刘世荣. 树干液流对环境变化响应研究进展[J]. 生态学报,2009,29(10):5600-5610.
[6]刘茜. 樟树叶片蒸腾特性及其与生理生态因子关系分析[J]. 中南林业科技大学学报,2009,29(1):1-5.
[7]王兵,郭浩. 影响丝栗栲树干液流速度的环境因子分析[J]. 南京林业大学学报(自然科学版),2009,33(1):43-48.
[8]Schipka F,Heimann J,Leuschner C. Regional variation in canopy transpiration of central European beech forests[J]. Oecologia,2005,143(2):260-270.
[9]杨瑞,肖卫平,喻理飞. 喀斯特区不同生境中云南鼠刺树干液流研究[J]. 中国水土保持科学,2011,9(4):94-97.
[10]张中峰,黄玉清,李先琨,等. 岩溶区青冈栎树干液流特征及其与环境因子关系[J]. 中国岩溶,2008,27(3):228-234.
[11]刘庆新,孟平,张劲松,等. 基于侧柏液流的测定对Granier原始公式系数进行校正[J]. 生态学报,2013,33(6):1944-1951.
[12]凡超,邱燕萍,李志强,等. 荔枝树干液流速率与气象因子的关系[J]. 生态学报,2014,34(9):2401-2410.
[13]丁访军,王兵,赵广东. 毛竹树干液流变化及其与气象因子的关系[J]. 林业科学,2011,47(7):73-81.
[14]池波,蔡体久,满秀玲,等. 大兴安岭北部兴安落叶松树干液流规律及影响因子分析[J]. 北京林业大学学报,2013,35(4):21-26.
[15]王文,朱烨,诸葛绪霞,等. 尾巨桉树干液流特性及其影响因子分析[J]. 水土保持通报,2013,33(3):159-164.
[16]黄玉清,张中峰,何成新,等. 岩溶区青冈栎整树蒸腾的季节变化[J]. 应用生态学报,2009,20(2):256-264.
[17]喻理飞,朱守谦,叶镜中,等. 退化喀斯特森林自然恢复过程中群落动态研究[J]. 林业科学,2002,38(1):1-7.
[18]Campbell G S,Norman J M. An introduction to environmental biophysics[M]. New York:Springer,1977,21(2):104.
[19]王华,赵平,蔡锡安,等. 马占相思树干液流与光合有效辐射和水汽压亏缺间的时滞效应[J]. 应用生态学报,2008,19(2):225-230.
[20]丁筱玲,赵立新,张业民. 植物茎液流速及蒸腾量动态测试仪[J]. 农业工程学报,2000,16(3):46-49.
[21]Hansen S. Daisy,a flexible Soil-Plant-Atmosphere system model[R]. Dept Agric,2002.
[22]黄德卫,张德强,周国逸,等. 鼎湖山针阔叶混交林优势种树干液流特征及其与环境因子的关系[J]. 应用生态学报,2012,23(5):1159-1166.
