干旱风沙区立地条件、季节变化对柠条灌丛“虫岛”效应的影响
2018-04-11郗伟华刘任涛刘佳楠常海涛罗雅曦
郗伟华, 刘任涛, 刘佳楠, 赵 娟, 常海涛, 罗雅曦, 张 静, 马 继
(1.山西师范大学 生命科学学院, 山西 临汾041004;2.宁夏大学 西北土地退化与生态恢复国家重点实验室培育基地, 银川 750021)
近年来,由于沙地灌丛具有“资源岛”功能[1],并能够产生“虫岛”现象[2],沙地灌丛的生态效应研究已开始引起广泛关注。不同立地条件灌丛林地由于土壤表层流沙覆盖情况、微地形等环境因素的差异性,导致土壤性质和地表植被群落组成也产生深刻变化[3-5]。土壤动物是沙地生态系统中的重要组成部分,其产卵、孵化、交配、取食及行为活动将会受到因立地条件不同而导致其群落结构空间分布存在差异性[3-4]。同时,伴随季节变化而引起的土壤温度、水分等环境因素的变化,也将影响到土壤动物的生活史过程和生物生态适应条件[6]。因此,随着季节变化,不同立地条件沙地灌丛对土壤动物空间分布的影响亦可能不同,结果可能将产生不同的“虫岛”效应[7]。所以,研究不同立地条件沙地灌丛“虫岛”效应的季节分布特征,对于沙地灌丛生态学研究、人工林管理利用及防沙治沙均具有重要的理论与实践意义。
目前,关于沙地灌丛和土壤动物关系的研究,在国内外开展了较多。Pen-Mouratov等[8]研究了以色列Negev沙漠中丛枝霸王(Zygophyllumdumosum)灌丛和土壤线虫群落分布的关系,发现灌丛下土壤湿度和有机质含量对土壤线虫群落及其营养功能群组成具有显著影响。Doblas-Miranda等[9]研究了地中海干旱系统中灌丛生境差异对大型无脊椎动物组成的影响及其季节变化,发现灌丛生境能够维持较高的物种丰富度、个体数和生物量。Blaum等[10]研究了萨瓦那草原灌丛覆盖程度对节肢动物多样性的影响,指出了节肢动物中存在着对不同灌丛生境的指示种。Zhao等[2]对科尔沁沙地小叶锦鸡儿和黄柳灌丛和土壤动物空间分布关系进行研究,提出了灌丛“虫岛”术语,并揭示了沙地灌丛“虫岛”效应的形成机理。刘任涛[11]对沙地灌丛的“肥岛”和“虫岛”关系、形成过程、特征及其与生态系统演替的关系进行了总结。刘任涛等[12]在宁夏盐池毛乌素沙地南麓开展了沙地柠条灌丛“虫岛效应”随林龄变化的研究。在此基础上,刘任涛等[13]比较了科尔沁沙地和毛乌素沙地灌丛“虫岛”效应特征,并且Liu等[14]总结出了沙地灌丛“虫岛”概念、分析方法与模型构建,为沙地灌丛土壤动物生态学研究提供了模式。综合分析表明,沙地灌丛“虫岛”的研究内容,成为沙地灌丛“沃岛效应”的重要补充[11]。但是,关于不同立地条件下沙地灌丛“虫岛”效应研究,报道较少。关于立地条件下沙地灌丛“虫岛”效应的季节分布特征,尚不清楚。
鉴于此,在宁夏盐池依据有无流沙覆盖土壤立地条件选择柠条灌丛林地为研究对象,于春、夏和秋季分别在灌丛内外样点中调查地面节肢动物群落结构特征,采用“相对相互作用强度指数”(Relative interaction intensity index,RⅡ),比较分析不同立地条件下3个季节沙地灌丛“虫岛”特征,旨在丰富沙地灌丛“沃岛效应”研究内容,为沙地生境人工林建设与管理、沙漠化防治和退化生态系统恢复提供依据。
1 研究地区与研究方法
1.1 研究区概况
研究区位于宁夏盐池县境内东北部(37°49′N,107°30′E,海拔1 348 m)。该区域属于中温带半干旱区,年平均气温7.7℃,最热月(7月)平均气温22.4℃,最冷月(1月)平均气温-8.7℃;≥10℃年积温2 751.7℃。年降水量为280 mm,主要集中在7—9月,占全年降水量的60%以上,且年际变率大,年蒸发量2 710 mm。年平均风速2.8 m/s,冬春风沙天气较多,每年5 m/s以上的扬沙达323次。
研究区地带性土壤类型为灰钙土。研究样地包括2种立地条件:(1) 有流动沙地覆盖的人工柠条林地,林带间距为7~8 m,灌丛高度为1.30~1.50 m,密度13~14 株/100 m2;灌丛间为裸沙生境,几无草本分布。(2) 邻近地区且在背风方向相距60~70 m的无流沙覆盖人工柠条林地,林带间距为10~12 m,灌丛高度为0.50~0.60 m,密度8~10株/100 m2;灌丛间有较多草本植物分布。主要植物包括柠条锦鸡儿(Caraganakoushinskii)、猪毛菜(Salsolacollina)、猪毛蒿(Artemisiascoparia)、山苦荬(Ixerischinensis)、白草(Pennisetumcentrasiaticum)、胡枝子(Lespedezabicolor)和牛枝子(Lespedezapotaninii)等。
1.2 试验设计与取样调查
选择有沙和无沙覆盖土壤地表两种土壤立地条件柠条林地为研究对象,二者相距60 m左右。每种立地条件有3个重复样地,每个样地选择长势良好且比较一致的4株灌丛进行标记,分别在标记灌丛下和灌丛外布置1个调查样点,灌丛外样点距离灌丛下样点5 m左右。共得到(4个灌丛外+4个灌丛下)×3重复样地×2个立地条件=48个样点。采用国际通用的陷阱诱捕法采集地面节肢动物,具体方法见参考文献[12]。
于2012年春季(5月初)、夏季(7月底)、秋季(9月底)布设取样,每次持续时间均为14 d(每隔3 d检查收集1次,避免采集标本的损失)。春季,由于有流沙覆盖柠条林地的一个重复样地的诱捕器遭到破坏,结果有沙覆盖柠条林地共得到16个取样点。3个季节共获得有效取样点136个。试验结束后将收集到的地面节肢动物标本带回实验室进行鉴定分类。节肢动物标本分类鉴定主要依据《中国土壤动物检索图鉴》[15]、《昆虫分类》[16]和《宁夏贺兰山昆虫》[17]等。
1.3 数据处理
在每个季节的每个样地中,分别将标记灌丛内外相似生境的4个采样点数据进行合并计算,得到地面节肢动物个体数(个体数/4陷阱)和类群数。在此基础上,计算地面节肢动物Shannon指数和优势度指数。
Shannon指数:
H=-∑PilnPi
(1)
优势度指数:
D=-∑(Pi)2
(2)
式中:Pi=ni/N,ni为第i类群动物的个体数;N为群落内动物的总个体数。
利用Sorenson指数[18]分析2种立地条件柠条林地面节肢动物群落相似性。
Sorenson指数(SS):
SS=2c/(a+b)
(3)
式中:c为共有类群数;a,b分别为各沙地生境的类群数。
“相对互相作用强度指数”(Relative interaction intensity,RⅡ)能够在种群和群落2个水平上能够表达灌丛“虫岛”的量化特征[13-14],故本文采用RⅡ来分析灌丛的“虫岛”强度。
相对互相作用强度指数:
RⅡ=(S-O)/(S+O)
(4)
式中:S为测定的灌丛下量值;O为测定灌丛外量值。
公式的生态学意义[13-14]包括:(1) 当灌丛内外均有节肢动物分布且量值相同(即:A=B)时,RⅡ=0,表示节肢动物对灌丛内外微生境存在广适性。(2) 当灌丛内外均有节肢动物分布且灌丛内数值A大于灌丛外B(即:A>B)时,即RⅡ>0,表示灌丛“虫岛”具有正效应。(3) 当灌丛内有节肢动物分布而灌丛外无节肢动物分布(即:A≠0,B=0)时,RⅡ=1,表示节肢动物对灌丛内微生境具有特定选择性而产生灌丛“虫岛”正效应。(4) 当灌丛内外均有节肢动物分布且灌丛内数值A小于灌丛外B(即:B>A)时,RⅡ<0,表示节肢动物更喜于在灌丛外微生境中生存而表现出灌丛“虫岛”负效应。(5) 当灌丛外存在节肢动物分布而灌丛内无分布(即:B≠0,A=0)时,RⅡ=-1,表示节肢动物对灌丛外微生境具有特定选择性而表现出灌丛“虫岛”负效应。
采用SPSS 15.0软件进行统计分析。采用非参数检验—独立样本t检验方法,分析同一季节不同立地条件沙地灌丛“虫岛”分布的差异性。利用单因素方差分析和多重比较,分析季节变化、立地条件对沙地灌丛“虫岛”分布影响。采用描述统计计算各指标的平均值和标准误。
2 结果与分析
2.1 土壤动物群落组成与数量特征
从表1可以看出,3个季节两种立地条件柠条林地生境共获得地面节肢动物11目38科41个类群。其中,有沙覆盖柠条林地在春季、夏季和秋季分别获得14,24,14个类群,平均个体数分别为107,96,61只/4陷阱;无沙覆盖柠条林地在春季、夏季和秋季分别获得21,21,17个类群,平均个体数分别为154,67,65只/4陷阱。在春季、夏季和秋季,两种立地条件柠条林地地面节肢动物共有类群数分别有10,15,11个,相似性指数分别为0.57,0.67,0.71。
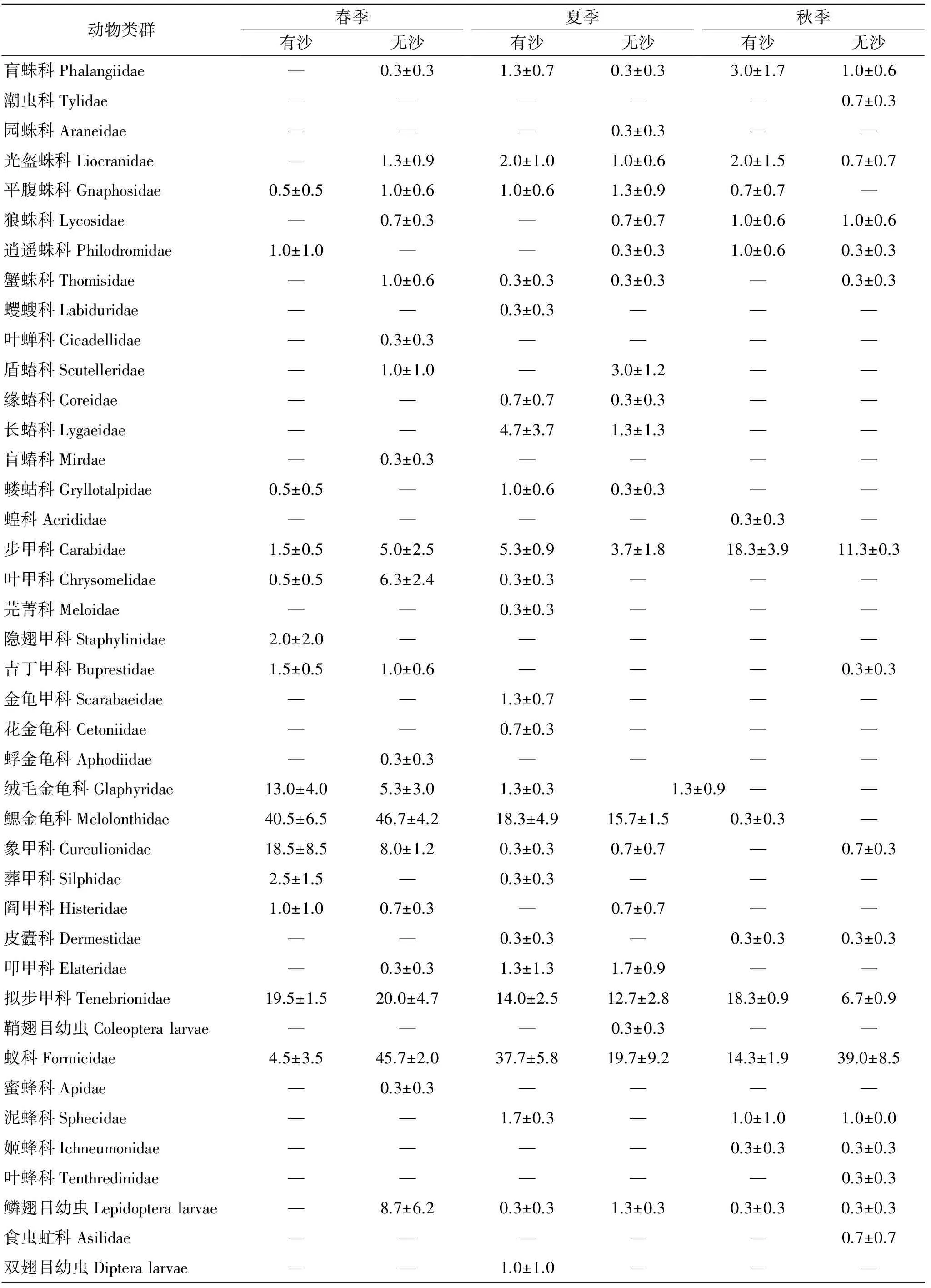
表1 地面节肢动物类群个体数(平均值±标准误) 只/4陷阱
注:“—”表示不存在节肢动物活动,有沙=有流沙覆盖土壤地表,无沙=无流沙覆盖土壤地表。
2.2 基于类群水平的灌丛“虫岛”比较
从表2可以看出,3个季节2种立地条件生境中,只有11个节肢动物类群表现出了“虫岛”特征。其中,盾蝽科、叶甲科、吉丁甲科、葬甲科只在单季节单个样地生境中出现“虫岛”特征,表现为盾蝽科出现在夏季无沙覆盖柠条林地中(RⅡ值<0),叶甲科出现在春季无沙覆盖柠条林地中(RⅡ值>0),吉丁甲科(RⅡ值<0)和葬甲科(RⅡ值>0)出现在春季有沙覆盖柠条林地中。象甲科在春季2种立地条件中均呈现出“虫岛”特征,而且两种立地条件间灌丛“虫岛”RⅡ无显著差异性(p>0.05),RⅡ值>0;泥蜂科在夏季有沙柠条林地和秋季无沙柠条林地中才呈现出“虫岛”特征,而且RⅡ值>0;鳞翅目幼虫在春季和夏季的无沙覆盖柠条林地中呈现出“虫岛”特征,而且RⅡ值>0。绒毛金龟科在春季2种林地条件生境以及夏季有沙覆盖柠条林地生境中均呈现出“虫岛”特征。鳃金龟科类群在春季和夏季2种立地条件中均呈现出灌丛“虫岛”特征,而且两种立地间灌丛“虫岛”RⅡ值均无显著差异性(p>0.05),但在春季无沙覆盖柠条林地呈现出正效应(RⅡ>0),而在春季有沙覆盖林地以及夏季2种立地条件生境中均呈现出负效应(RⅡ<0)。
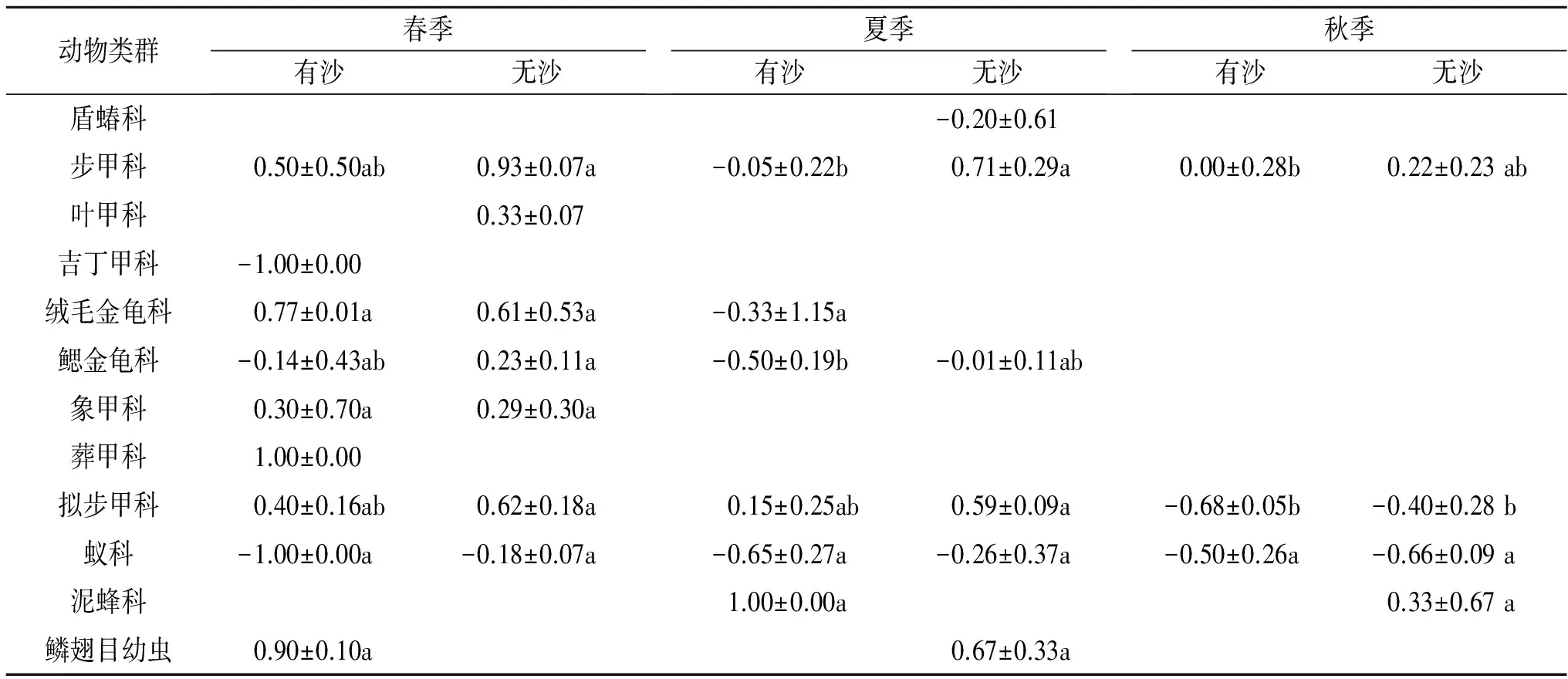
表2 不同立地条件节肢动物类群灌丛“虫岛”效应随季节的变化
注:不同小写字母表示显著差异(p<0.05),有沙=有流沙覆盖土壤地表,无沙=无流沙覆盖土壤地表。
由表2可知,步甲科、拟步甲科和蚁科在3个季节的每一种立地条件中均呈现出“虫岛”特征,但具体分布情况又存在较大差异性。在春季和秋季,步甲科类群表现为两种立地条件生境间灌丛“虫岛”RⅡ值无显著差异性(p>0.05),但是在夏季,步甲科类群表现为无沙覆盖柠条林地灌丛“虫岛”RⅡ值显著高于有沙覆盖柠条林地(p<0.05),而且有沙覆盖柠条林地步甲科类群灌丛“虫岛”呈现出负效应,即RⅡ<0。拟步甲科和蚁科类群灌丛“虫岛”RⅡ值分布则与步甲科类群的情况完全不同,表现为在春、夏和秋季,两种立地条件生境间RⅡ值均无显著差异性(p>0.05),但同时又表现作用方向的差异性。两种立地条件生境下,拟步甲科类群灌丛“虫岛”在春和夏季均呈现出正效应(RⅡ>0),而在秋季表现为负效应(RⅡ<0)。两种立地条件生境下,蚁科类群灌丛“虫岛”在3个季节总是表现为负效应,即RⅡ<0。
2.3 基于群落水平的灌丛“虫岛”比较
从图1可以看出,只有夏季且只有节肢动物个体数灌丛“虫岛”RⅡ值在两种立地条件生境间存在显著差异性,表现为无沙覆盖柠条林地显著高于有沙覆盖林地(p<0.05),而且有沙覆盖柠条林地节肢动物个体数灌丛“虫岛”RⅡ值<0。节肢动物个体数灌丛“虫岛”RⅡ值在春季和秋季两种立地条件间均无显著差异性(p>0.05)。并且,节肢动物丰富度、Shannon指数和优势度指数灌丛“虫岛”RⅡ值在春季、夏季和秋季两种立地条件间亦均无显著差异性(p>0.05)。但是,灌丛对节肢动物丰富度、Shannon指数和优势度指数的“虫岛”RⅡ值作用方向随着立地条件和季节变化而存在一定差异性。
由图1可知,两种立地条件生境节肢动物个体数灌丛“虫岛”在春季和秋季表现为相反的作用方向,即春季两种立地条件生境灌丛对节肢动物个体数均呈现出正效应(RⅡ>0),而在秋季均呈现出负效应(RⅡ<0)。
在春季,节肢动物丰富度灌丛“虫岛”表现为两种立地条件生境均呈现出正效应(RⅡ>0);到了夏季,节肢动物丰富度灌丛“虫岛”表现为有沙覆盖柠条林地呈现出负效应(RⅡ<0),而在无沙覆盖柠条林地均呈现出正效应(RⅡ>0);但到了秋季则完全相反,节肢动物丰富度灌丛“虫岛”表现为有沙覆盖柠条林地呈现出正效应(RⅡ>0),而在无沙覆盖柠条林地均呈现出负效应(RⅡ<0)。基于Shannon指数和优势度指数的灌丛“虫岛”作用方向则完全相反,表现为节肢动物Shannon指数在3个季节的两种立地条件生境中均呈现出正效应(RⅡ>0),而节肢动物优势度指数在3个季节的两种立地条件生境中均呈现出负效应(RⅡ<0)。
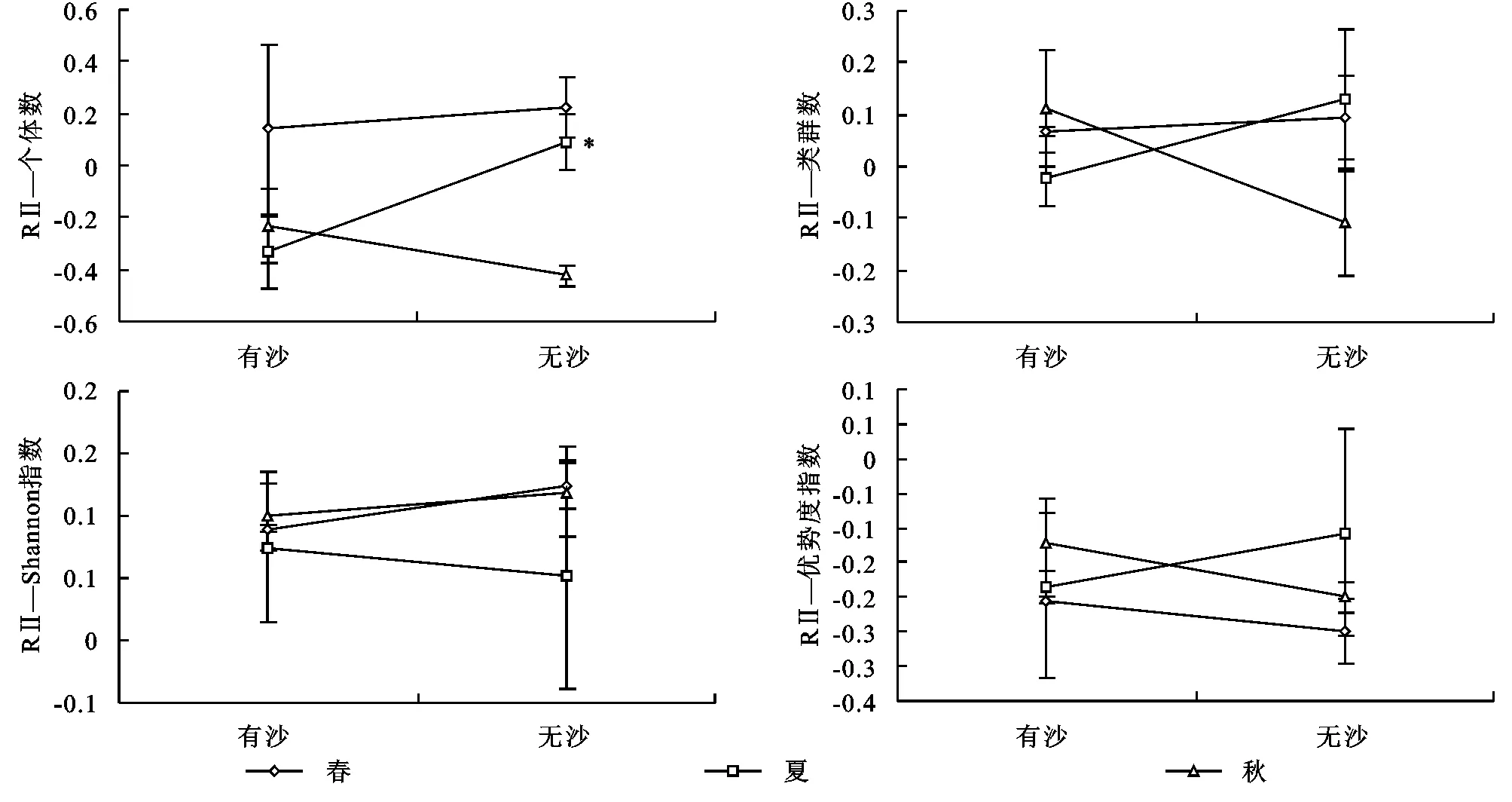
注: *表示p<0.05,有沙=有流沙覆盖土壤地表,无沙=无流沙覆盖土壤地表。
图1基于群落指数的2种立地条件柠条灌丛“虫岛”效应(RⅡ)比较
3 讨论与结论
在宁夏盐池半干旱农牧交错区土地发生退化、沙化地段,定居于沙地生境中的灌丛呈现出特殊的生境“岛屿”,为节肢动物的生存、繁殖提供了理想的栖息地,在流动沙地的固定与恢复过程中可能起到“种源”(节肢动物扩散源)作用,成为沙化草地生态系统食物网结构构建、物质循环与能量流动过程中不可或缺的一环[11]。本研究中,两种土壤立地条件柠条林地的地面节肢动物群落相似性指数随着季节更替而逐渐增加,达到中等相似水平,反映了基于地面节肢动物类群组成的相似性和柠条林地背景的相似性。但是,由于两种立地条件柠条林地的土壤表层流沙覆盖情况、微地形等各种立地条件的差异,导致不同立地条件中灌丛林的适应性生长特征不同[19],进而影响到土壤结构、植被组成存在差异性,直接导致不同土壤立地条件灌丛内外地面节肢动物呈现出不同的空间分布[3-4]。本研究中,盾蝽科灌丛“虫岛”出现在夏季无沙覆盖柠条林地中,叶甲科灌丛“虫岛”出现在春季无沙覆盖柠条林地中,吉丁甲科和葬甲科灌丛“虫岛”则出现在春季有沙覆盖柠条林地中,说明不同季节不同立地条件生境中,不同节肢动物类群表现出了相似或相反的灌丛“虫岛”分布特征,反映了地面节肢动物对随季节改变的不同立地条件生境的选择性和适应性[20]。
已有研究表明,灌丛内外微生境的差异性导致节肢动物的空间分布差异性,导致不同节肢动物类群的灌丛“虫岛”作用方向和作用强度[12,21]。本研究中,在春季,无沙覆盖柠条林地中鳃金龟科类群主要分布在灌丛内微生境中,而在有沙覆盖林地中鳃金龟科类群主要分布在灌丛外微生境中。说明立地条件对灌丛内外的空间分布产生了深刻影响,这与不同立地条件不同节肢动物类群的生活史过程、生态生物学特性以及对微生境的强烈选择性和适应性特征密切相关[16-17,20]。春季是鳃金龟科成虫越冬后的发生盛期[22],无沙覆盖灌丛内和有沙覆盖灌丛外土壤环境诸如土壤孔隙度、土壤透水、透气性等条件较好,更有利于鳃金龟科幼虫取食孵化和定居[2,16]。
随着季节更替,立地条件对节肢动物类群的灌丛“虫岛”分布亦产生显著影响。例如,步甲科类群灌丛“虫岛”RⅡ值表现为春季和秋季两种立地条件生境间无显著差异性,但是在夏季则呈现出显著差异性,并且灌丛“虫岛”从正效应变为负效应。一方面说明灌丛“资源岛”效应能够为步甲科类群提供充足的食源条件,吸引这些类群个体数前来定居生存[2],特别是在夏季无沙覆盖的柠条林地中表现出了较高灌丛“虫岛”强度。同时,也说明在随着季节条件的变化灌丛内外微生境的改变对步甲科类群个体数分布亦产生了深刻影响,在有沙覆盖柠条林地中更多的步甲科类群选择在灌丛外生存[21]。在夏季有沙覆盖柠条林地中,灌丛下土壤温度较低、水分条件较好吸引较多的节肢动物类群包括捕食性动物类群前来定居生存,在有限的食源条件下产生竞争力,进而导致这些移动能力较强的步甲科类群更多选择在灌丛外活动、生存[23]。
但是,拟步甲科和蚁科类群灌丛“虫岛”RⅡ值分布则与步甲科类群的情况完全不同。在春季和夏季,两种立地条件拟步甲科类群灌丛“虫岛”均呈现出正效应,而在秋季均表现为负效应。一方面说明立地条件对灌丛内外拟步甲科类群个体数分布影响较小,同时也说明拟步甲科类群个体数在灌丛内外的空间分布更多受到季节变化的调控[9]。拟步甲科是荒漠草原生态系统重要的碎屑性节肢动物类群[24],春季和夏季灌丛下积累较多的枯枝落叶吸引拟步甲科前来定居,而经过秋季,灌丛外枯枝落叶的增多又导致更多的拟步甲科个体前来取食,这些均有利于荒漠草原区地面枯落物的分解和物质循环过程[5]。蚁科类群灌丛“虫岛”受立地条件和季节变化的影响较小,均呈现负效应。一方面说明蚂蚁类群在荒漠草原区分布的广布性,同时也说明更多的蚂蚁类群个体数则主要分布在灌丛外微生境中,这表征了灌丛结构特征及其灌丛下植被分布不利于或阻碍了蚂蚁群居性活动,导致蚂蚁更喜于在灌丛外裸露生境中筑巢、生存[25-26]。
在季节变化和立地条件的双重影响下,地面节肢动物群落水平的灌丛“虫岛”亦呈现出不同的表现形式和强度[13]。本研究中,只有夏季且只有节肢动物个体数灌丛“虫岛”RⅡ值在两种立地条件生境间存在显著差异性。在夏季,节肢动物个体数灌丛“虫岛”RⅡ值表现为无沙覆盖柠条林地显著高于有沙覆盖林地,并且从灌丛“虫岛”的正效应变为负效应,这与步甲科类群个体数的灌丛“虫岛”RⅡ值分布情况相似。步甲科是荒漠草原区的优势类群之一[24],其个体数分布表征了地面节肢动物群落的个体数量特征。根据岛屿生物地理学理论,干旱风沙区生境中灌丛斑块充当土壤动物“生境岛屿”的关键性角色[27]。当夏季有沙覆盖柠条林地土壤生境条件易受风蚀等外界干扰情况下,无沙覆盖林地灌丛内土壤生境条件,包括温度、水分、养分对土壤动物的存活、定居和繁殖就具有更为突出的作用[2],基于地面节肢动物个体数的灌丛“虫岛”效应表现为无沙覆盖林地显著高于有沙覆盖林地,这说明了无沙覆盖林地灌丛对地面节肢动物个体数具有较强的保育效应,有利于生物多样性保护与生态系统恢复过程[28]。但是,在春季和秋季,节肢动物个体数灌丛“虫岛”表现出相反的作用方向。春季两种立地条件生境中节肢动物个体数主要分布在灌丛下微生境中,这与灌丛的“资源岛”效应密切相关[2]。但是在秋季,两种立地条件生境中节肢动物个体数主要分布在灌丛外微生境中,这与拟步甲科类群个体数在灌丛内外的分布情况相似。
两种立地条件节肢动物丰富度、Shannon指数和优势度指数灌丛“虫岛”均受到季节改变的影响较大。在春季,两种立地条件生境灌丛对节肢动物丰富度呈现出聚集作用,这与节肢动物个体数在灌丛内外的分布情况相似,主要原因在于灌丛“资源岛”效应吸引较多的节肢动物类群在灌丛下定居、存活,表征了较为明显的灌丛“虫岛”特征[2]。到了夏季,无沙覆盖柠条林地灌丛下微生境中分布有较多的节肢动物类群数,而有沙覆盖柠条林地灌丛外微生境中分布有较多的节肢动物类群数,并且秋季和夏季两种立地条件表现出了相反的灌丛“虫岛”作用方向,这与前人在科尔沁沙地[2]和黑河流域戈壁生态系统[28]中对灌丛微生境中土壤动物的研究结果相悖。这说明在调查灌丛微生境中土壤动物空间分布时,需要同时考虑立地条件和季节变化的双重因素。但是,基于Shannon指数和优势度指数的灌丛“虫岛”作用方向则完全相反,并且不受立地条件和季节变化的影响,反映了灌丛对地面节肢动物多样性具有正效应,有利于地面节肢动物多样性恢复和维持,这与Zhao[2]和李锋瑞[5]等的研究结果相吻合。
综合分析表明,立地条件和季节变化导致了不同节肢动物类群的灌丛“虫岛”作用方向和作用强度产生差异性。立地条件对节肢动物个体数灌丛“虫岛”的影响较大,并受到季节变化的调控。节肢动物多样性指数灌丛“虫岛”效应主要受到季节变化的影响,而受到立地条件的影响较小。灌丛对地面节肢动物多样性具有正效应,有利于地面节肢动物多样性恢复和维持。因此,在研究立地条件对灌丛“虫岛”效应及其对生物多样性分布影响的情况下,需要考虑地面节肢动物群落结构的季节分布特征。
参考文献:
[1]苏永中,赵哈林,张铜会.科尔沁沙地几种灌木半灌木对土壤肥力的影响[J].应用生态学报,2002,13(7):802-806.
[2]Zhao H L, Liu R T. The‘bug island’effect of shrubs and its formation mechanism in Horqin Sand Land, Inner Mongolia[J]. Catena, 2013,105:69-74.
[3]董炜华,李金霞,刘继亮.沙丘和丘间低地地面节肢动物群落比较研究[J].干旱区资源与环境,2016,30(1):157-161.
[4]董炜华,殷秀琴,顾卫,等.农牧交错带不同土地类型土壤动物生态特征研究:以内蒙古卓资山为例[J].干旱区地理,2008,121(5):63-70.
[5]李锋瑞,刘继亮,刘长安,等.土地覆被变化与管理对土壤动物群落演变的耦合效应[J].中国沙漠,2012,32(2):59-69.
[6]刘任涛,朱凡.半干旱区草地生境中大型土壤动物群落季节动态[J].生态环境学报,2013,22(1):72-77.
[7]Dunning J B, Danielson B J, Pulliam H R. Ecological processes that affect populations in complex landscapes[J]. Oikos, 1992,65(1):169-175.
[8]Pen-Mouratov S, Rakhimbaev M, Barness G, et al. Spatial and temporal dynamics of nematode populations underZygophyllumdumosumin arid environments[J]. European Journal of Soil Biology, 2004,40(1):31-46.
[9]Doblas-Miranda E, Sánchez-Pinero F, Gonzalez-Megías A. Different microhabitats affect soil macroinvertebrate assemblages in a Mediterranean arid ecosystem[J]. Applied Soil Ecology, 2009,41(3):329-335.
[10]Blaum N, Seymour C, Rossmanith E, et al. Changes in arthropod diversity along a land use driven gradient of shrub cover in savanna rangelands:Identification of suitable indicators[J]. Conservation Biology, 2009,18(5):1187-1199.
[11]刘任涛.沙地灌丛的“肥岛”和“虫岛”形成过程、特征及其与生态系统演替的关系[J].生态学杂志,2014,33(12):3463-3469.
[12]刘任涛,朱凡.基于群落与种群水平的沙地柠条灌丛“虫岛效应”随林龄的变化[J].应用与环境生物学报,2015,21(4):689-694.
[13]刘任涛,王少昆,周娟.科尔沁和毛乌素沙地灌丛“虫岛”效应比较[J].中国沙漠,2015,35(6):1599-1606.
[14]Liu R T, Pen-Mouratov S, Steinberger Y. Shrub cover expressed as an‘arthropod island’in xeric environments[J]. Arthropod-Plant Interactions, 2016,10(5):393-402.
[15]Yin W Y. Pictorial keys to soil animals of China[M]. Beijing: Science Press, 2000.
[16]郑乐怡,归鸿.昆虫分类[M].南京:南京师范大学出版社,1999.
[17]王新谱,杨贵军.宁夏贺兰山昆虫[M].银川:宁夏人民出版社,2010.
[18]刘任涛,赵哈林,刘继亮.黄河兰州段人工林大型土壤动物群落结构[J].土壤学报,2009,46(3):179-182.
[19]蒋齐,李生宝,潘占兵,等.人工柠条灌木林营造对退化沙地改良效果的评价[J].水土保持学报,2006,20(4):23-27.
[20]任国栋,于有志.中国荒漠半荒漠的拟步甲科昆虫[M].河北保定:河北大学出版社,2000.
[21]刘继亮,李锋瑞,刘七军,等.黑河流域干旱荒漠土壤动物群落组成与多样性的季节变异[J].中国沙漠,2010,30(2):342-349.
[22]韩国君,张文忠,韩国辉,等.黑绒鳃金龟生物学特性研究[J].吉林林业科技,2002,31(6):15-17.
[23]Sanders N J, Gotelli N J, Sarah E W, et al. Assembly rules of ground-foraging ant assemblages are contingent on disturbance, habitat and spatial scale[J]. Journal of Biogeography, 2007,34:1632-1641.
[24]刘任涛,郗伟华,朱凡.宁夏荒漠草原地面节肢动物群落组成及季节变化特征[J].草业学报,2016,25(6):126-135.
[25]Crist T O, Guertin D S, Wiens J A, et al. Animal movements in heterogeneous landscapes: An experiment with Eleodes beetles in shortgrass prairie[J]. Functional Ecology, 1992,6(5):536-544.
[26]Stapp P. Microhabitat use and community structure of darkling beetles(Coleoptera:Tenebrionidae)in shortgrass prairie:effects of season, shrub cover and soil type[J]. American Midland Naturalist, 1997,137(2):298-311.
[27]Garner W, Steinberger Y. A proposed mechanism for the formation of‘fertile islands’in the desert ecosystem[J]. Journal of Arid Environments, 1989,16(3):257-262.
[28]Liu J L, Li F R, Liu C, et al. Influences of shrub vegetation on distribution and diversity of a ground beetle community in a Gobi desert ecosystem[J]. Biodiversity Conservation, 2012,21(10):2601-2619.
