东北平原典型玉米种植区农业景观植物多样性研究*
——以昌图县为例
2018-04-09边振兴李晓璐
边振兴, 李晓璐, 于 淼
(沈阳农业大学土地与环境学院 沈阳 110161)
农业景观是由集约化农用地和非耕作生境组成的景观镶嵌体[1]。生物多样性是人类赖以生存的物质基础[2], 它决定着景观生态系统的稳定性和复杂性[3]; 景观异质性是生物多样性的前提, 同时也是生态系统生物共生的基础[3]。随着农业集约化进程的持续推进, 农业景观的物质流和能量流发生变化,农业景观生态系统结构和功能单一, 生态系统功能和服务严重缺失, 进而导致生物群落多样性严重下降[4-8]。农业景观生态系统的持续发展离不开生物多样性早已成为共识[9-10]。农业景观中相当多非耕作生物与农业目标生物相伴而生, 它们的生存离不开农业景观所提供的食物和栖息地[11-12]。农田景观中非耕作生物的多样性通过天敌、传粉者等的生物控制、作物授粉等生态系统服务直接影响农田内作物的产量, 是农业景观可持续发展的基础, 对维持区域乃至全球的可持续发展具有重要意义[2]。因此, 农业景观中非耕作生境及其异质性的存在与维持对生物多样性保护具有重要作用[13-15]。欧盟和美国早已将增加景观异质性和非耕作生境构建, 从而提高农业景观中生物多样性列入农业环境保护的重要内容中[16-17]。在保障粮食供给以满足日益增长的人口需求的背景下, 如何权衡农业用地、农业生产实践与保护生物多样性及其提供的生态系统服务已成为人类面临的最严峻挑战[18-21], 且已有研究[22-23]证实可以实现这种双赢的局面。国内的许多研究者从景观、局地和田块等不同水平对农业景观中的生物多样性和生态系统服务等开展了大量研究[24-26], 但生物类群以动物为主, 受“杂草”这一名称和人类对农产品产量追求等诸多负面的影响, 农业景观生态系统中非耕作生境植物多样性的保护及其分布格局、生态过程和生态效应的研究尚未得到足够的关注[27]。而农业景观中非耕作生境中的植物在整个农业景观生态系统服务的发挥和可持续发展中具有极其重要的作用。
东北平原面积达35万km2, 是我国面积最大的平原, 耕地面积约0.20亿hm2, 其中玉米(Zea maysL.)种植面积约 0.15亿 hm2, 占耕地面积的 76.67%,同时也是粮食主产区和重要的商品粮区, 玉米产量占全国总产量的30%左右[28]。随着农业机械的发展,在东北平原区已实现玉米的规模化种植, 而农业景观是区内的主体景观类型, 这种规模化种植方式导致景观均质化, 从而使生物多样性降低, 同时区内人类活动强度和频率的增加显著影响了景观结构,使得农业景观中的生物多样性和生态系统服务受到严重影响。但目前关于区内农业景观中生物多样性的研究较少[29-30]。区内农业景观中最适宜的非耕作生境比例和较为稳定的生境类型, 以及不同生境类型生物物种优势度差异、生物多样性及其与景观异质性之间的关系等都有待研究, 同时这些问题的研究也是探讨区域农业用地、生物多样性保护和生态系统服务维持的前提和基础[31]。为此本研究以辽宁省昌图县为例对东北平原典型玉米种植区农业景观中植物多样性进行调查, 探讨区内不同非耕作比例和生境类型植物物种多样性及不同生境类型植物多样性与景观异质性之间的关系, 为今后农业景观中生物多样性研究提供理论基础, 也为未来区域可持续农业景观构建提供数据支撑。
1 研究区概况
研究区位于辽宁省铁岭市昌图县, 主要河流有辽河、招苏台河等, 主要作物为玉米, 是东北平原典型的玉米种植区之一, 也是全国著名农业大县、重点商品粮生产基地。地理位置为 42°33′~43°29′N,123°32′~124°26′E, 总 面 积 4 317 km2, 耕 地 面 积2 666.67 km2, 其中玉米种植面积为1 500 km2, 占县域面积的34.75%, 占耕地面积的56.25%。区内地势由东部低山丘陵向西部辽河平原过渡, 主要土壤类型为棕壤、黑土、草甸土和风砂土, 属于中温带亚湿润大陆性季风气候, 年平均降雨量约600 mm, 年均温 6~7 ℃。区内以旱作农业景观为主, 农田是最主要的景观类型, 居民点景观类型广泛散落在区域内, 大小不一, 林地景观多呈斑块状散布在农田和居民点周边, 以人工种植的杨树林为主。
2 研究方法
2.1 样地设置和数据的收集
在研究区卫星影像和2011年土地利用现状图的基础上, 运用 1 km×1 km 网格将研究区进行划分,并按照不同非耕作生境比例在研究区内选择了 20个网格作为植物多样性调查区域(图1)。采用法瑞学派的典型样地法对各调查区域内主要非耕作生境(草地、沟渠、果园、林地和田间路等)的植物群落进行调查, 记录样方内植物的物种名、多优度、群集度和各样方的地理坐标等。
野外植物多样性调查在2017年进行, 其中共选择植物多样性调查区域20个(图1), 每个调查区域选择2个主要的非耕作生境类型进行样方布设, 每个非耕作生境类型设置3个样方(图2), 整个研究区共设置调查样方120个(其中草地15个, 沟渠12个, 果园9个, 林地51个, 田间路33个)。经调查发现, 天然林均为单一的樟子松(Pinus sylvestrisvar.mongolicaLitv.)林, 人工林绝大多数均为单一的白杨(Populus tomentosaCarr.)林, 且 20个调查区域内各生境中几乎不存在灌木层, 因此本研究和数据分析中均选取草本层数据。
2.2 数据处理和指标选择
2.2.1 植物物种鉴别与群落指数
采用人工鉴别法[32], 通过查阅植物志、植物学、植物图鉴等[33-37]文献中的植物分类检索表、计算机植物检索数据库, 并结合实地拍摄的植物根、茎、叶、花和果实等主要器官的照片对所采集的植物样株进行鉴别, 鉴别层次为植物科(Familia)、属(Genus)和种(Species)。最终确定出 120个样方中草本植物的科、属、种和数量, 并计算出5种生境中各物种的重要值和植物多样性特征指数。
1)物种多样性指数
选取Shannon-Wiener多样性指数为指标进行分析[38], 计算公式为:
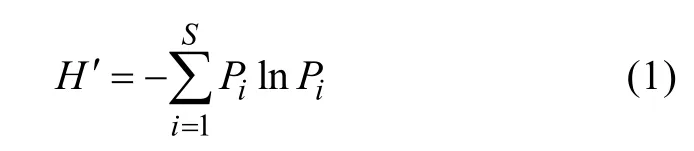
式中:H′为Shannon-Wiener多样性指数;S为样方内物种数;iP为物种i在群落中的重要性, 即物种i的多优度与总多优度和的比值。
2)物种丰富度指数
选取Margalet丰富度指数进行分析, 计算公式为:
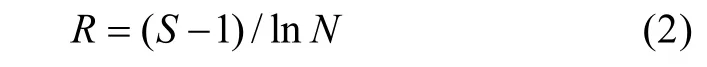
式中:R为 Margalet丰富度指数,S为样方内物种数,N为样方内所有物种的个体总数。
3)物种均匀度指数
选取Pielow均匀度指数为指标进行分析, 计算公式为:

式中:E为 Pielow均匀度指数,H′为 Shannon-Wiener指数,S为样方内物种数。
4)β多样性指数
选取 Whittaker Beta多样性指数来反映生境中群落组成变化的幅度和分化程度[39], 计算公式:

式中: β为Whittaker Beta多样性指数;γ为某取样生境中的总物种数,α为某生境类型所有取样点物种数的平均值。
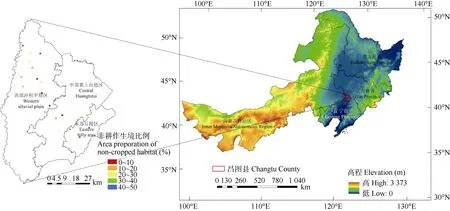
图1 研究区在东北平原的位置及其20个不同非耕作生境比例的植物群落调查区域的分布Fig. 1 Location of the study area in the Northeast China Plain and distribution of 20 investigation areas of plant communities with different area proportions of non-cropped habitats
2.2.2 景观异质性指数
根据已有的研究[40-42]选取了景观水平上的景观异质性表征指数: 蔓延度指数(CONTAG)、香浓多样性指数(SHDI)和香浓均匀度指数(SHEI)。
1)蔓延度指数(contagion index, CONTAG)
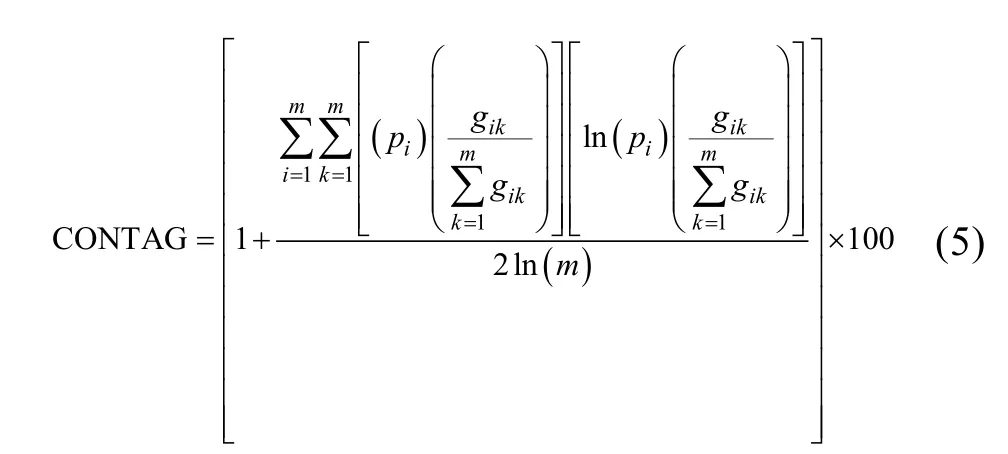
式中:pi为第i类景观斑块所占总面积的比例,m为景观类型总数量,gik为随机两个相邻栅格单元属于类型i和类型k的概率。
2)香浓多样性指数(Shannon diversity index,SHDI)
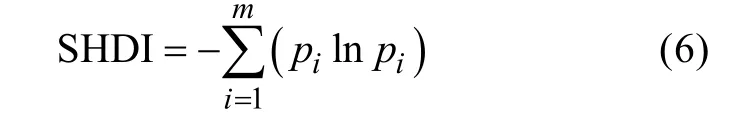
式中:pi为第i类景观所占总面积的比例,m为景观中各种景观类型总数。
3)香浓均匀度指数(Shannon evenness index,SHEI)
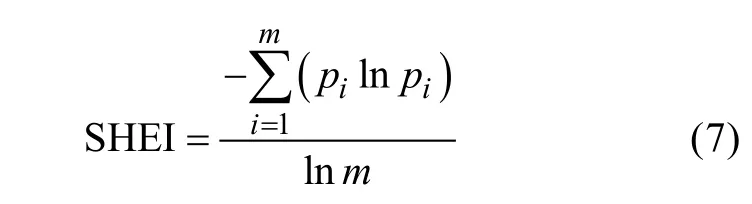
式中:pi为第i种类型景观占总面积的比例,m为出现景观类型的总数量。
2.2.3 统计分析方法
借助于 Fragstats 4.2分别提取植物多样性调查区域所在的20个1 km×1 km网格的3种景观异质性指数, 并运用办公软件Office 2007处理计算野外调查数据。结果分析选择单因素方差分析方法和相关性分析方法, 并运用SPSS 16.0完成, 所有结果分析图均采用Excel和Origin 8.0绘制完成。其中, 按非耕作生境比例统计时, 将同一比例区间内的不同类型生境中植物群落指数作为一个样本数据处理; 按非耕作生境类型统计时, 将同一类型生境中不同非耕作生境比例的植物群落指数作为一个样本数据处理。
3 结果与分析
3.1 不同非耕作生境比例区域的景观异质性
从不同植物群落调查区域来看, 景观异质性与景观内非耕作生境比例具有显著相关性(表1)。随着植物群落调查区域内非耕作生境比例的增加, 景观香浓多样性和香浓均匀度指数增加, 景观斑块形状复杂程度逐渐增加; 蔓延度恰好相反(表1)。表明非耕作生境比例越低, 各景观类型所占面积的比例差异越大, 则出现少数几种景观占优势现象的可能性越大。非耕作生境比例0~10%和40%~50%的植物群落调查区域之间景观斑块复杂程度和主要景观优势度差异较大, 且与其他比例的植物群落调查区域之间存在显著性差异(表1)。
3.2 不同类型非耕作生境的植物群落优势度
农业景观内不同类型非耕作生境中草本层植物调查数据的统计结果(表2)发现, 研究区农业景观中非耕作生境共有129种植物, 分属于35科91属; 其中菊科(Asteraceae)33种, 占总种数的25.58%, 19属,占总属数的20.88%; 禾本科(Gramineae)17种, 占总种数的 13.18%, 15属, 占总属数的 16.48%; 蓼科(Polygonaceae)9种, 占总种数的 6.98%, 2属, 占总属数的 2.20%; 豆科(Leguminosae)8种, 占总种数的6.20%, 7属, 占总属数的7.69%; 其他共62种, 48属。
优势种对群落结构特征具有重要影响, 物种的重要值展现了一个物种在整个区域内的优势度, 物种-累积重要值变化曲线可以反映生境中优势种优势度的强度[31]。不同生境中植物群落主要优势种重要值(表2)分析显示, 各生境中物种的重要值最高的均为禾本科植物, 林地、田间路和草地生境中多花黑麦草的重要值最高, 果园生境中假泽早熟禾的重要值最高, 沟渠生境中菵草的重要值最高, 且长戟叶蓼、矮蒿和绢毛悬钩子具有相似的重要值, 它们分别在不同植物群落调查区域内的沟渠生境中形成优势群落。表明不同生境中植物群落主要优势种组成相似性较高, 但优势种优势度的强度在不同生境中具有一定的差异, 尤其是果园和沟渠生境植物群落优势种优势度最为明显(图3)。推测不同生境之间物种组成具有一定相似性, 但却具有不同的生态和服务功能[31]。
结合物种数目-累积重要值变化曲线发现: 研究区内农业景观中植物群落优势种优势度在果园生境中最明显, 沟渠生境次之, 其他依次为草地生境、田间路生境、林地生境, 且果园和沟渠生境中显著高于草地、田间路和林地生境(图 3)。研究区内农业景观中果园、沟渠、草地和田间路生境中优势种优势度均较为明显, 且物种总数明显低于林地生境。推测在农业景观中, 随着人类活动干扰的增强, 植物群落中物种数会减少, 但优势种的地位会更加突出, 优势种对整个植物群落的控制作用愈加显著[31,38]。
3.3 植物群落调查区域内物种多样性变化特征
3.3.1 不同非耕作生境比例植物群落调查区域内物种多样性特征
调查结果(图4)表明, 研究区农业景观不同非耕作生境比例的植物群落调查区域内植物多样性特征指数存在显著性差异。随着非耕作生境比例的增加,Shannon-Wiener多样性指数和均匀度指数均先增加后减小, 在非耕作生境比例 20%~30%时达到最大值;物种丰富度指数也呈先增大后减小的趋势, 但在非耕作生境比例40%~50%时突增, 且明显高于其他比例, 原因可能是本研究非耕作生境比例40%~50%的植物群落调查区域位于林场周围, 林地景观面积占比接近农田景观面积占比, 从而植物群落调查区域内林地景观的优势度凸显, 进而该比例区间内农业景观中植物物种丰富度指数突增。
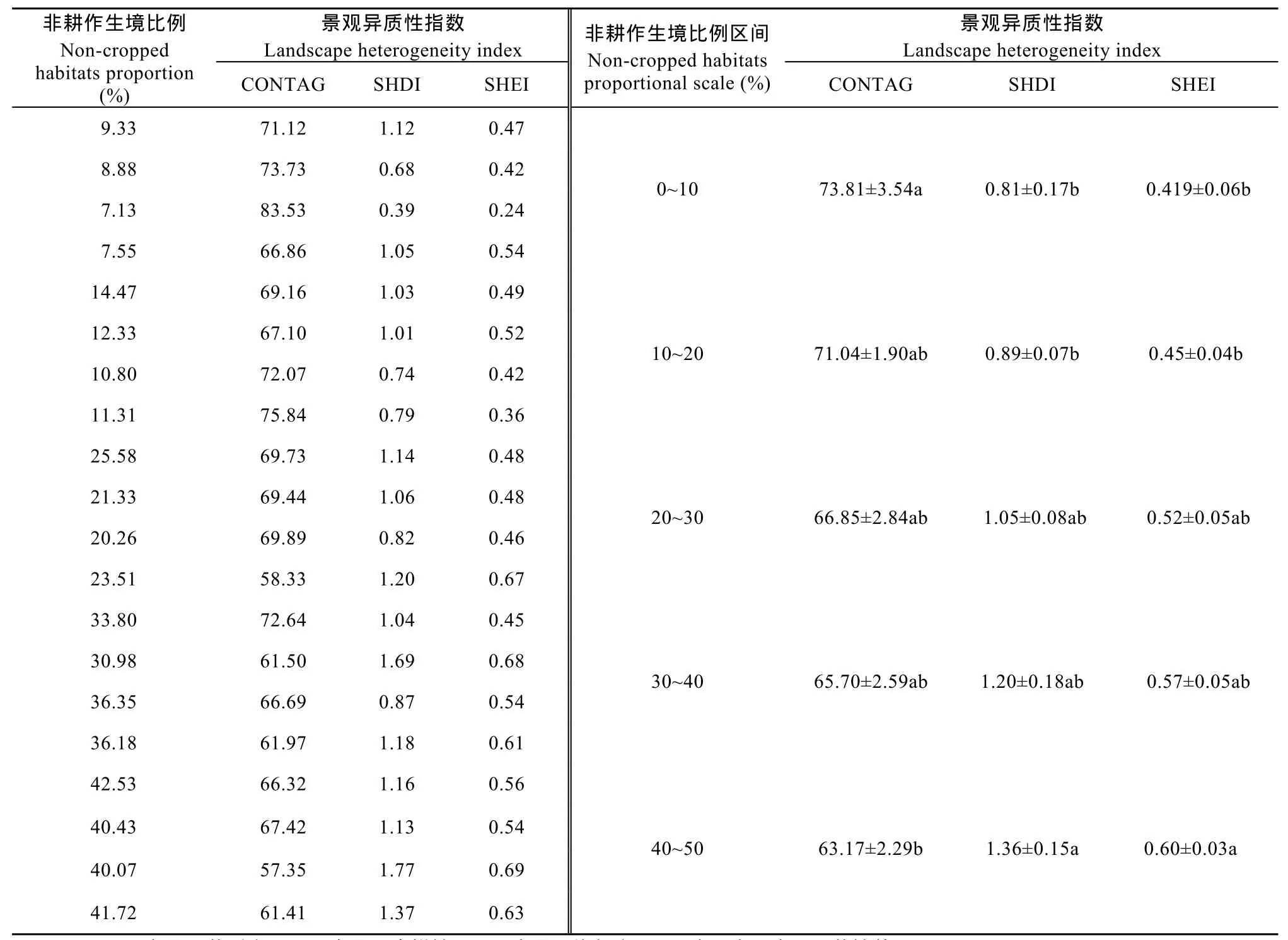
表1 不同非耕作生境比例农业景观的景观异质性指数Table 1 Landscape heterogeneity indexes of agricultural landscapes with different non-cropped habitats proportions
从图 4可以看出, 非耕作生境比例 30%~40%和40%~50%的植物群落调查区域之间的Shannon-Wiener多样性指数、物种丰富度指数和物种均匀度指数均存在较大的变异性(图4), 推测植物多样性特征与植物群落调查区域内生境类型、生境内生态因子异质性和生境外部人类干扰的强度和频度密切相关。综合分析, 非耕作生境比例 20%~30%时, Shannon-Wiener多样性指数、均匀度指数和丰富度指数相对较高且较稳定。因此, 针对植物群落多样性,20%~30%为农业景观中最优的非耕作生境比例。
3.3.2 不同类型非耕作生境植物多样性特征
调查结果(图5)表明: 果园生境Shannon-Wiener多样性指数最高, 草地生境次之, 其余依次为沟渠生境、林地生境、田间路生境, 且果园、草地和沟渠生境之间差异性不显著; 草地生境植物物种丰富指数显著高于其他 4种生境, 依次为草地>林地>沟渠>果园>田间路; 植物群路物种均匀度指数在果园、沟渠、草地、林地和田间路生境中顺次降低, 其中果园和沟渠生境物种均匀度显著高于草地、林地和田间路生境。各指数标准差反映出草地和果园生境的Shannon-Wiener多样性指数、丰富度指数和均匀度指数均存在较大的变异性, 受生境外部人类干扰频率、强度的影响较大。
β多样性指数反映了不同生境的植物群落组成在整个研究区各调查区域间的变化幅度和分化程度。结果(图 5)表明: 林地生境的 β多样性指数显著高于其他4种生境, 田间路生境次之, 沟渠、草地和果园生境顺次降低, 但差异并不显著。这种变化趋势表明研究区农业景观内非耕作生境中的植物群落结构受人类活动强度和干扰频率(田间路生境明显高于沟渠、草地和果园生境)影响十分强烈, 同时受到生境内生态因子异质性(果园和沟渠生境内生态因子异质性较低)影响较大, 其中林地生境内草本层受乔灌层种间差异影响较大。通过β多样性指数标准差反映出不同植物群落调查区域间的草地和田间路生境中β多样性指数存在较大的变异性, 林地、沟渠和果园生境中β多样性指数变异性较小(图5)。

表2 农业景观中不同类型非耕作生境的优势植物种Table 2 Dominant plant species in different non-cropped habitats of agricultural landscape
3.4 不同类型非耕作生境物种多样性与景观异质性的关系
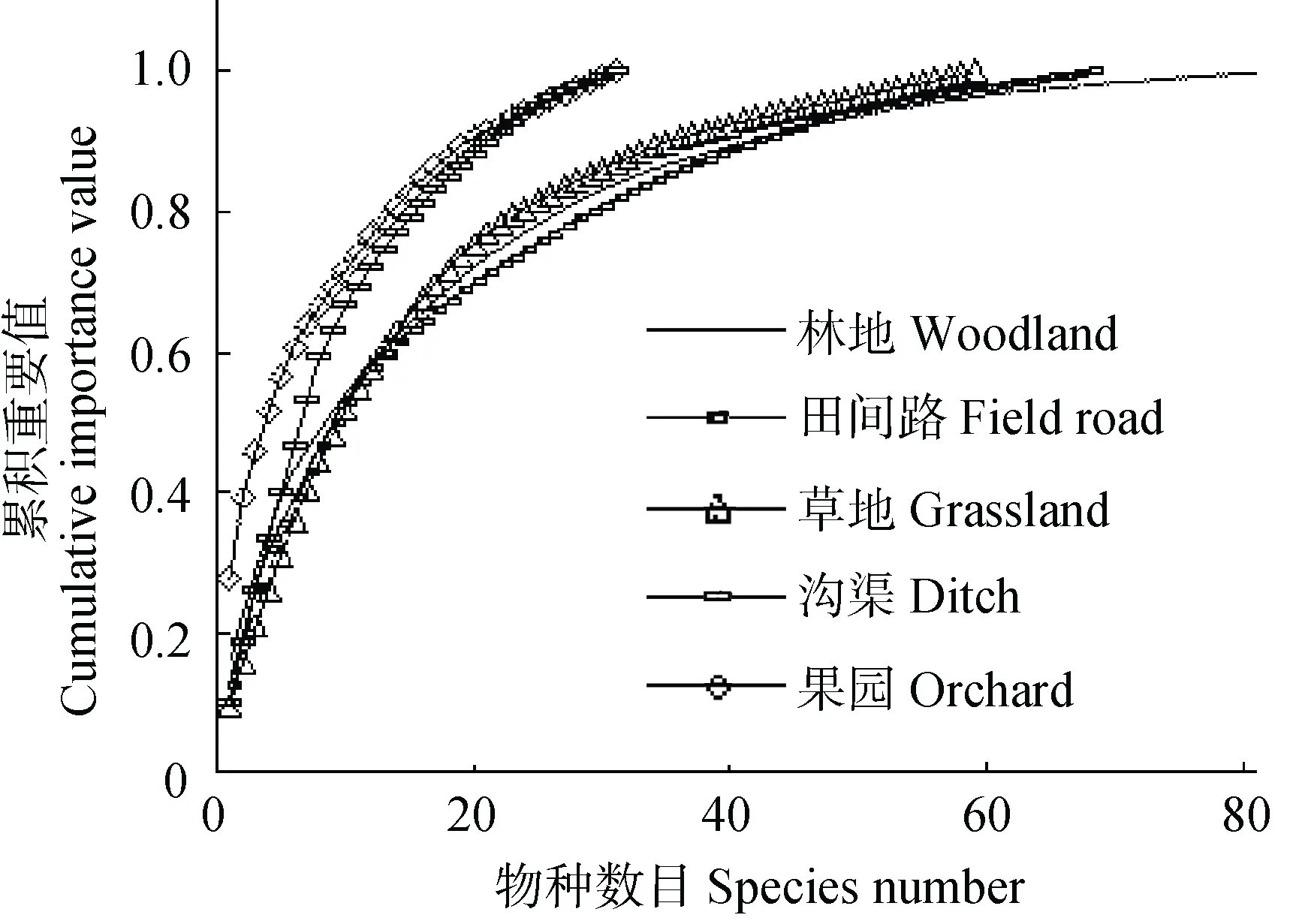
图3 农业景观中不同非耕作生境的植物群落物种-累积重要值变化情况Fig. 3 Variations of cumulative importance values with plant species number in different non-cropped habitats of agricultural landscape
分析结果(表 3)表明: 在草地生境中, 植物群落的Shannon-Wiener多样性、丰富度指数与景观异质性显著相关, 植物群落物种均匀度与景观异质性之间相关性不显著。在果园、沟渠、林地和田间路生境中, 景观异质性与植物群落多样性相关性均不显著。草地生境植物群落物种多样性指数与景观蔓延度指数呈反比, 与景观 Shannon多样性指数和均匀度指数均呈正比, 且植物群落 Shannon-Wiener多样性指数、丰富度指数和均匀度指数与 3种景观异质性指数之间相关程度一致, 相关系数绝对值分别为0.949、0.973和0.771; 其他4种生境中植物多样性指数与景观异质性指数之间相关程度均不相同。
草地生境中景观异质性与植物物种多样性特征指数的相关程度为: 丰富度指数>Shannon-Wiener多样性指数>均匀度指数; 沟渠生境中植物 Shannon-Wiener多样性与景观蔓延度、景观Shannon均匀度指数之间相关程度差异较小, 与景观 Shannon多样性指数相关性较大, 且相关程度为植物Shannon-Wiener多样性指数>均匀度指数>丰富度指数; 果园生境中景观异质性指数与植物多样性指数相关性最强; 林地生境中景观异质性与植物Shannon-Wiener多样指数相关程度差别较小, 其中与景观蔓延度指数、Shannon多样性指数相关性最强的均为植物丰富度指数, 与景观 Shannon均匀度指数相关性最强的为植物Shannon-Wiener多样性指数; 田间路生境中植物丰富度指数与景观 Shannon均匀度指数、Shannon多样性指数相关性最强, 其次为均匀度指数与景观蔓延度指数相关性最强。
不同非耕作生境的景观蔓延度指数与 3种植物物种多样性特征指数之间的相关性为草地和果园>林地>沟渠>田间路, 景观 Shannon多样性指数为草地>沟渠>果园、田间路和林地, 景观 Shannon均匀度指数为果园、草地>沟渠>田间路、林地。综合分析, 景观异质性与植物物种多样性相关性在草地、果园、沟渠、田间路和林地生境中顺次降低, 表明不同生境中干扰类型、方式和强度不同, 会导致景观异质性与物种多样性的相关性和相关程度不同。
4 讨论与结论
研究表明, 研究区内农业景观中非耕作生境比例与景观蔓延度呈反比, 与景观多样性和均匀度均呈正比, 非耕作生境比例逐渐增加, 景观斑块形状复杂程度增加, 各景观类型所占面积的比例差异减小, 同时出现少数几种景观占优势现象的可能性降低。随着非耕作生境比例增加, 植物物种多样性指数和均匀度指数均先增加后减小, 在非耕作生境比例为 20%~30%时达到最大值。非耕作生境比例在0~40%区间内逐渐增加, 植物物种丰富度指数先增加后减小, 在非耕作生境比例为20%~30%达到最大值; 非耕作生境比例高于 40%时, 随着非耕作生境比例的增加, 植物物种丰富度指数又逐渐增加, 且非耕作生境比例为 40%~50%时植物物种丰富度指数高于非耕作生境比例为20%~30%。与此同时, 非耕作生境比例为40%~50%, 景观斑块复杂程度和景观类型占优势程度均出现较大差异, 从而引起植物物种丰富度发生显著变化。本研究结果表明: 农业景观最佳的非耕作生境比例为20%~30%。
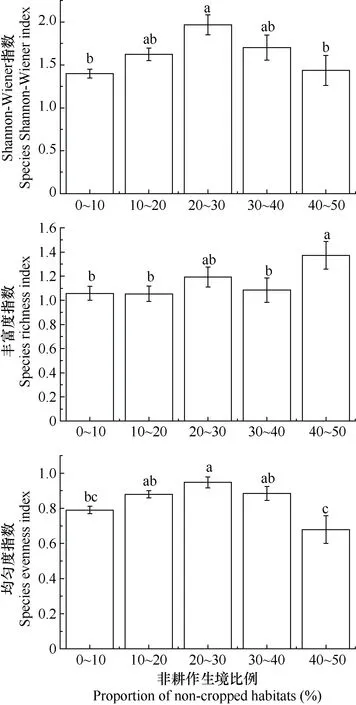
图4 不同非耕作生境比例农业景观的植物群落物种多样性特征指数Fig. 4 Plant diversity characteristic indexes of agricultural landscapes with different non-cropped habitats proportions
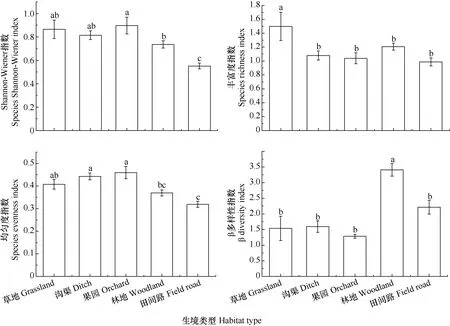
图5 农业景观中不同类型非耕作生境的植物群落物种多样性特征指数Fig. 5 Plant species diversity characteristic indexes of different non-cropped habitats of agricultural landscape
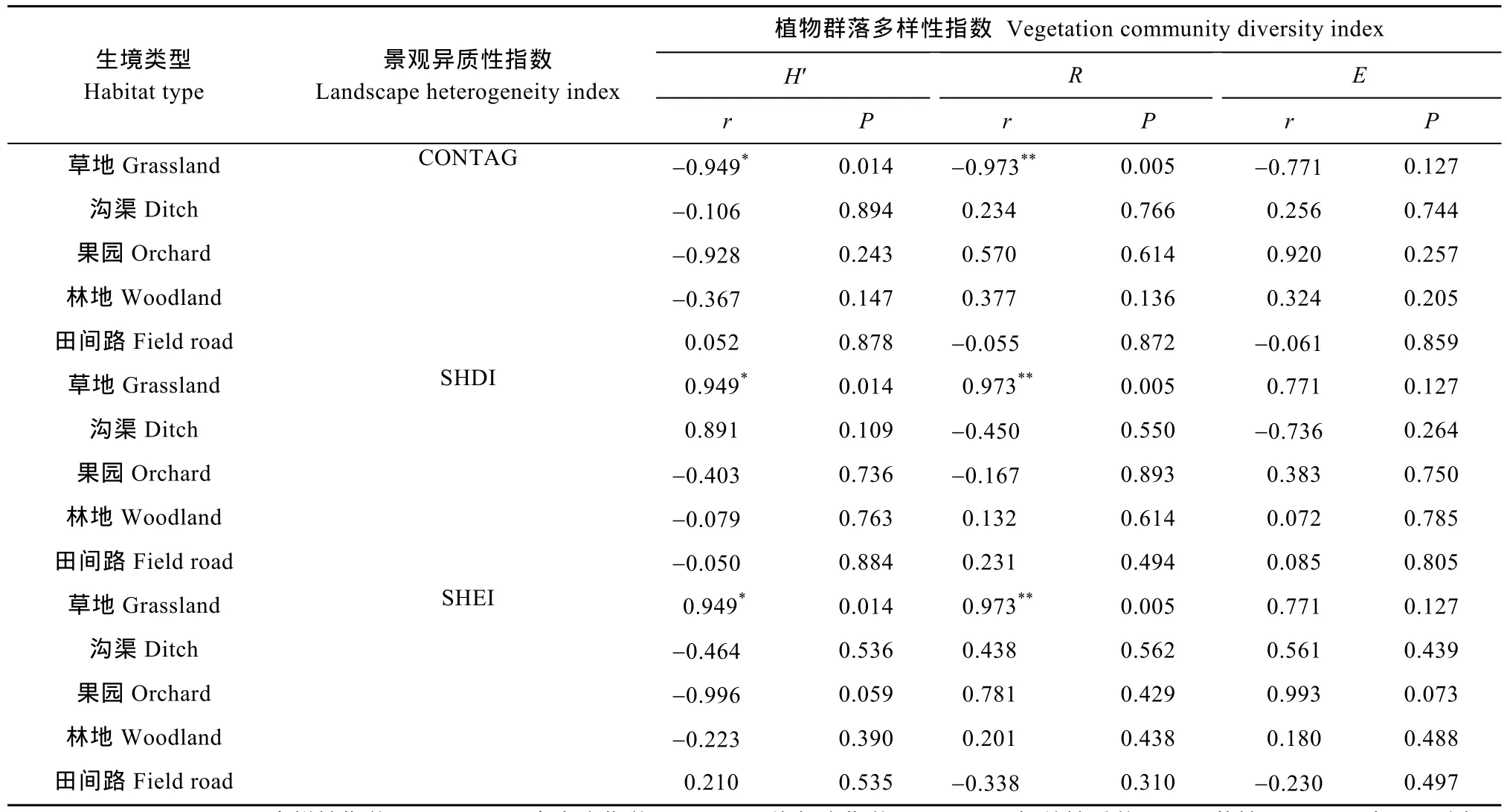
表3 不同类型的非耕作生境中景观异质性与物种多样性相关性Table 3 Correlation between landscape heterogeneity and species diversity in different non-cropped habitats
林地生境中草本植物群落物种丰富度最高, 但受乔灌层影响植物群落优势种类型以禾本科、菊科和莎草科为主; 果园、草地和田间路生境受人类干扰强烈, 植物群落优势种主要以假泽早熟禾和多花黑麦草等耐踩踏和耐药性较强的禾本科植物为主;沟渠生境中的长戟叶蓼是一种湿生植物, 是 5种生境内的特有种, 也是沟渠生境中的植物群落优势种之一, 与其他植物群落优势种之间的优势度差异较小, 因此沟渠对湿生植物物种多样性的保护具有积极作用。研究区内农业景观中植物群落优势种重要值分析表明, 分布于不同生境的植物群落的主要优势种组成具有一定相似性, 但优势种优势度的强度及其承载的生态和服务功能却不相同。物种数目-累积重要值分析表明, 研究区内农业景观中植物群落优势种优势度为: 果园>沟渠>草地>田间路>林地,且果园、沟渠和草地生境中的优势种优势度明显高于田间路和林地生境中的优势种优势度。在农业景观中, 果园、草地和田间路生境受到人类活动干扰强烈, 植物群落中抗药性强和耐践踏的优势种则会通过自然选择, 呈现集群或连片状分布, 对整个植物群落的控制作用愈加显著。
果园、草地和沟渠生境中植物物种Shannon-Wiener多样性指数和均匀度指数显著高于林地和田间路生境,但不同植物群落调查区域内果园和草地生境中存在较大的变异性; 物种丰富度指数依次为草地>林地>沟渠>果园>田间路, 且各植物群落调查区域内草地、果园和田间路生境存在较大变异性。因此, 草地、果园和田间路生境中的植物群落结构受人类干扰频率、干扰强度和生境内生态因子异质性较大, 较为脆弱,不利于物种多样性保护; 沟渠和林地生境中植物物种Shannon-Wiener多样性指数、丰富度指数和均匀度指数均较高, 且较为稳定, 说明沟渠和林地对植物物种多样性保护具有积极作用。
农业景观异质性与不同非耕作生境植物物种多样性相关性为草地>果园>沟渠>田间路>林地, 其中草地生境中植物群落物种 Shannon-Wiener多样性、丰富度指数与景观异质性显著相关。综合不同非耕作生境中植物群落优势种分布情况, 物种 Shannon-Wiener多样性指数、丰富度指数和均匀度指数分布规律分析, 不同干扰类型、方式和强度使得景观异质性与不同植物物种多样性特征指数相关性有所不同。草地生境主要以草本植物群落为主, 每种植物的重要值均小于 1, 且植物群落结构相对单一, 个体数、物种总数和覆盖度也易受影响, 因此植物物种多样性与景观异质性之间相关程度为物种丰富度指数>物种 Shannon-Wiener多样性指数>物种均匀度指数。由于人类对果业产品质量和数量的需求, 果园生境人为干扰和选择强度大, 耐药、抗虫、喜阴的草本植物物种优势度凸显, 从而降低植物物种数, 使得物种多样性指数变化明显, 且与景观异质性间相关性较强。沟渠生境分为两种类型, 其中一种为自然沟渠, 另一种为人工硬化沟渠, 而沟渠的硬化导致部分植物物种直接消失, 故植物覆盖度降低, 均匀度降低, 因此沟渠生境中植物物种多样性与景观蔓延度、景观香浓均匀度指数之间相关程度差异较小,与景观多样性指数差异较大, 表现为物种Shannon-Wiener多样性指数>物种均匀度指数>物种丰富度指数。随着耕作季节的不同, 田间路生境会受到不同程度的踩踏以及除草剂的影响, 植物物种个体数变化明显, 植物群落优势种的生态优势凸显,因此植物物种丰富度指数和均匀度指数与景观异质性相关性较强。林地生境中分布有乔木、灌木和草本, 受植物群落种间竞争和林窗的影响, 草本植物物种数和个体数有较大差别, 从而使得林地生境中物种丰富度和物种Shannon-Wiener多样性指数与景观异质性相关性较强。
草地生境景观结构单一, 果园和田间路生境受到人类干扰最为严重, 景观均质性较强, 植物物种多样性较低, 植物群落优势种生态优势凸显, 不利于物种多样性保护; 沟渠生境较为特殊, 虽然受到人为硬化的干扰, 但植物多样性适中, 且较为稳定,变化幅度和分化程度较低, 更是特有种长戟叶蓼等湿生植物的繁殖、栖息地, 有利于湿生植物的保护;林地生境是动植物的主要栖息地和避难所, 其植物多样性最高, 且最为稳定, 对农田物种多样性保护和提高具有重要意义。物种丰富度指数和物种Shannon-Wiener多样性指数都在1 000 m的尺度下与景观异质性相关性最强[41]。因此, 在平原区景观异质性与农田植物多样性具有密切联系。
昌图县是我国著名农业大县、重点商品粮生产基地, 也是典型的北方波状平原旱作农业地区, 其农业景观是由集约化农用地和非耕作生境组成的景观镶嵌体[1], 生存于其中的非耕作生物均离不开农业景观所提供的食物和栖息地[11-12]。农业景观中的自然、半自然生境(如田间路、林地、果园、草地、沟渠等)为耕作生境中生物提供了丰富的源, 常作为生物的栖息地、繁殖地、食物源、避难所, 对于维持生物多样性有极为重要的影响[43]。深入研究农业景观异质性对生物多样性的影响, 通过权衡生物多样性的生态系统服务与农业生产, 构建可持续的农业景观是当前农业景观管理中亟待解决的科学问题[44]。已有的研究表明, 随着全球变化和人类活动干扰的增强, 农业景观中生物多样性急剧减少[45],道路等线状廊道使得连通性有所下降, 导致栖息于生境内的物种减少[46]。本研究也显示田间路生境中植物物种多样性指数、丰富度指数和均匀度指数均低于其他生境。在农业生产、生物多样性保护和生态系统服务可持续之间的权衡与取舍中, 合理地进行景观结构的优化, 通过对农业景观中非耕作生境的布局和管理, 使其生态服务效应最大化是目前农业生产与景观管理所面临的重大问题。本研究只针对草本植物的多样性特征指数进行了分析, 未考虑乔灌层, 忽略了乔灌层组成和结构对整个生态系统功能和下层草本植物多样性特征指数的影响[47]。研究区内农业景观中, 不同非耕作生境类型和比例下,植物物种优势度和多样性特征指数都存在一定的差异, 在未来的农业结构构建和农业景观物种多样性保护中应该充分考虑生境类型的影响, 同时注意当前农业景观中非耕作生境的合理配置和对物种多样性保护有积极作用的非耕作生境的保护、构建。在今后农业景观中生物多样性保护和农业生态系统的健康可持续发展的进一步研究中, 应该综合地貌、非耕作生境比例和类型、外部干扰强度和频率等多种影响因素对生物多样性、生态服务及农业产量的影响。
参考文献 References
[1] BENNETT A F, RADFORD J Q, HASLEM A. Properties of land mosaics: Implications for nature conservation in agricultural environments[J]. Biological Conservation, 2006,133(2): 250–264
[2] 傅伯杰. 景观多样性分析及其制图研究[J]. 生态学报, 1995,15(4): 345–350 FU B J. Landscape diversity analysis and mapping[J]. Acta Ecologica Sinica, 1995, 15(4): 345–350
[3] 秦向红. 景观异质性与生物多样性关系探讨[J]. 哈尔滨师范大学自然科学学报, 1997, 13(4): 98–102 QIN X H. An approach to relation between biological diversity and landscape heterogeneity[J]. Natural Sciences Journal of Harbin Normal University, 1997, 13(4): 98–102
[4] ROBINSON R A, SUTHERLAND W J. Post-war changes in arable farming and biodiversity in Great Britain[J]. Journal of Applied Ecology, 2002, 39(1): 157–176
[5] FOLEY J A, DEFRIES R, ASNER G P, et al. Global consequences of land use[J]. Science, 2005, 309(5734): 570–574
[6] MEEHAN T D, WERLING B P, LANDIS D A, et al. Agricultural landscape simplification and insecticide use in the Midwestern United States[J]. Proceedings of the National Academy of Sciences of the United States of America, 2011,108(28): 11500–11505
[7] FORMAN R T T, GODRON M. 景观生态学[M]. 肖笃宁,译. 北京: 科学出版社, 1990 FORMAN R T T, GODRON M. Landscape Ecology[M]. Xiao D N, Trans. Beijing: Science Press, 1990
[8] FORMAN R T T. Land Mosaics: The Ecology of Landscapes and Regions[M]. London: Cambridge University Press, 1995
[9] ALTIERI M A. The ecological role of biodiversity in agroecosystems[J]. Agriculture, Ecosystems & Environment, 1999,74(1/3): 19–31
[10] MARSHALL E J P, BROWN V K, BOATMAN N D, et al.The role of weeds in supporting biological diversity within crop fields[J]. Weed Research, 2003, 43(2): 77–89
[11] MIDGLEY G F. Biodiversity and ecosystem function[J].Science, 2012, 335(6065): 174–175
[12] TSCHARNTKE T, KLEIN A M, KRUESS A, et al. Landscape perspectives on agricultural intensification and biodiversity — ecosystem service management[J]. Ecology Letters,2005, 8(8): 857–874
[13] ERNOULT A, ALARD D. Species richness of hedgerow habitats in changing agricultural landscapes: Areαandrdiversity shaped by the same factors?[J]. Landscape Ecology,2011, 26(5): 683–696
[14] GROOT J C J, JELLEMA A, ROSSING W A H. Designing a hedgerow network in a multifunctional agricultural landscape:Balancing trade-offs among ecological quality, landscape character and implementation costs[J]. European Journal of Agronomy, 2010, 32(1): 112–119
[15] MARSHALL E J P, MOONEN A C. Field margins in northern Europe: Their functions and interactions with agriculture[J]. Agriculture, Ecosystems & Environment, 2002,89(1/2): 5–21
[16] JACKSON L, VAN NOORDWIJK M, BENGTSSON J, et al.Biodiversity and agricultural sustainability: From assessment to adaptive management[J]. Current Opinion in Environmental Sustainability, 2010, 2(1/2): 80–87
[17] WHITTINGHAM M J. The future of agri-environment schemes: Biodiversity gains and ecosystem service delivery?[J]. Journal of Applied Ecology, 2011, 48(3): 509–513
[18] GODFRAY H C J, BEDDINGTON J R, CRUTE I R, et al.Food security: The challenge of feeding 9 billion people[J].Science, 2010, 327(5967): 812–818
[19] PHALAN B, ONIAL M, BALMFORD A, et al. Reconciling food production and biodiversity conservation: Land sharing and land sparing compared[J]. Science, 2011, 333(6407):1289–1291
[20] TSCHARNTKE T, CLOUGH Y, WANGER T C, et al. Global food security, biodiversity conservation and the future of agricultural intensification[J]. Biological Conservation, 2012,151(1): 53–59
[21] GOMIERO T, PIMENTEL D, PAOLETTI M G. Is there a need for a more sustainable agriculture?[J]. Critical Reviews in Plant Sciences, 2011, 30(1/2): 6–23
[22] CLOUGH Y, BARKMANN J, JUHRBANDT J, et al. Combining high biodiversity with high yields in tropical agroforests[J]. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(20): 8311–8316
[23] SEUFERT V, RAMANKUTTY N, FOLEY J A. Comparing the yields of organic and conventional agriculture[J]. Nature,2012, 485(7397): 229–232
[24] 刘云慧, 宇振荣, 王长柳, 等. 坝上地区农田和恢复生境地表甲虫多样性[J]. 生态学报, 2011, 31(2): 465–473 LIU Y H, YU Z R, WANG C L, et al. The diversity of ground-dwelling beetles at cultivated land and restored habitats on the Bashang plateau[J]. Acta Ecologica Sinica, 2011,31(2): 465–473
[25] 宇振荣, 谷卫彬, 胡敦孝. 江汉平原农业景观格局及生物多样性研究——以两个村为例[J]. 资源科学, 2000, 22(2):19–23 YU Z R, GU W B, HU D X. On landscape pattern and biodiversity in rural areas of Jianghan Plain — Taking two villages as a case study[J]. Resources Science, 2000, 22(2):19–23
[26] 常虹, 张旭珠, 段美春, 等. 北京密云农业景观步甲群落空间分布格局[J]. 应用生态学报, 2012, 23(6): 1545–1550 CHANG H, ZHANG X Z, DUAN M C, et al. Spatial distribution pattern of carabid assemblage in agricultural landscape of Miyun County, Beijing[J]. Chinese Journal of Applied Ecology, 2012, 23(6): 1545–1550
[27] RUNDLÖF M, EDLUND M, SMITH H G. Organic farming at local and landscape scales benefits plant diversity[J]. Ecography, 2010, 33(3): 514–522
[28] 杨斌, 陈源泉, 隋鹏, 等. 东北平原玉米主产区不同施肥方式对农田生态系统健康的影响[J]. 作物杂志, 2010, (4):77–81 YANG B, CHEN Y Q, SUI P, et al. Comprehensive assessment on ecological health in intensive-cropland under different fertilization measures[J]. Crops, 2010, (4): 77–81
[29] 谷艳芳, 胡楠, 丁圣彦, 等. 开封地区不同土地利用方式农田杂草群落结构及动态[J]. 生态学报, 2007, 27(4):1359–1367 GU Y F, HU N, DING S Y, et al. Community structure and dynamics of agricultural weeds under different land utilization regimes in Kaifeng area[J]. Acta Ecologica Sinica, 2007,27(4): 1359–1367
[30] 汪洋, 王刚, 杜瑛琪, 等. 农林复合生态系统防护林斑块边缘效应对节肢动物的影响[J]. 生态学报, 2011, 31(20):6186–6193 WANG Y, WANG G, DU Y Q, et al. Influence of edge effects on arthropods communities in agroforestry ecological systems[J]. Acta Ecologica Sinica, 2011, 31(20): 6186–6193
[31] 卢训令, 汤茜, 梁国付, 等. 黄河下游平原不同非农生境中植物多样性[J]. 生态学报, 2015, 35(5): 1527–1536 LU X L, TANG Q, LIANG G F, et al. Plant species diversity of non-agricultural habitats in the lower reaches of the Yellow River Plain[J]. Acta Ecologica Sinica, 2015, 35(5):1527–1536
[32] 陈永富, 刘华, 陈巧. 植物鉴别方法研究现状与展望[J].世界林业研究, 2014, 27(4): 18–23 CHEN Y F, LIU H, CHEN Q. Status and prospect for the study of plants identification methods[J]. World Forestry Research, 2014, 27(4): 18–23
[33] 中国科学院中国植物志编辑委员会. 中国植物志[M]. 北京:科学出版社, 2004 Editorial Committee of “Flora Reipublicae Popularis Sinicae”in the Chinese Academy of Sciences. Flora Reipublicae Popularis Sinicae[M]. Beijing: Science Press, 2004
[34] 李书心. 辽宁植物志[M]. 辽宁: 辽宁科学技术出版社,1988: 11 LI S X. Flora Liaoningica Tomus[M]. Liaoning: Liaoning Science and Technology Press, 1988: 11
[35] 傅沛云. 东北草本植物志(第 12卷)[M]. 北京: 科学出版社,1998: 11 FU P Y. Flora Plantarum Herbacearum Chinae Boreali-Orientalis Tom.Ⅻ[M]. Beijing: Science Press, 1998: 11
[36] 马炜梁. 植物学[M]. 北京: 高等教育出版社, 2009 MA W L. Botany[M]. Beijing: Higher Education Press, 2009
[37] 中国科学院植物研究所. 中国高等植物图鉴[M]. 北京: 科学出版社, 1972 Institute of Botany, Chinese Academy of Sciences. Iconographia Cormophytorum Sinicorum Tomus[M]. Beijing: Science Press, 1972
[38] 马克平, 刘玉明. 生物群落多样性的测度方法Ⅰ α多样性的测度方法(下)[J]. 生物多样性, 1994, 2(4): 231–239 MA K P, LIU Y M. Measurement of biotic community diversity Ⅰ α diversity (Part 2)[J]. Chinese Biodiversity, 1994,2(4): 231–239
[39] 陈圣宾, 欧阳志云, 徐卫华, 等. Beta多样性研究进展[J].生物多样性, 2010, 18(4): 323–335 CHEN S B, OUYANG Z Y, XU W H, et al. A review of beta diversity studies[J]. Biodiversity Science, 2010, 18(4):323–335
[40] 邬建国. 景观生态学[M]. 第 2版. 北京: 高等教育出版社.2007 WU J G. Landscape Ecology[M]. 2nd ed. Beijing: Higher Education Press, 2007
[41] 刘栋. 黄河下游平原农业景观非农植物群落特征及其与景观异质性关系研究[D]. 开封: 河南大学, 2013 LIU D. Research on non-crop community characteristics and the relationship of landscape heterogeneity of agricultural landscape in the lower region of the Yellow River Plain[D].Kaifeng: Henan University, 2013
[42] 董翠芳. 景观异质性对植物多样性影响的多尺度分析——以巩义市为例[D]. 开封: 河南大学, 2014 DONG C F. The multi-scale effects of landscape heterogeneity on plant species diversity: A case study of Gongyi City[D].Kaifeng: Henan University, 2014
[43] FAHRIG L, BAUDRY J, BROTONS L, et al. Functional landscape heterogeneity and animal biodiversity in agricultural landscapes[J]. Ecology Letters, 2011, 14(2): 101–112
[44] 宋博, 丁圣彦, 赵爽, 等. 农业景观异质性对生物多样性及其生态系统服务的影响[J]. 中国生态农业学报, 2016, 24(4):443–450 SONG B, DING S Y, ZHAO S, et al. Effects of agricultural landscape heterogeneity on biodiversity and ecosystem services[J]. Chinese Journal of Eco-Agriculture, 2016, 24(4):443–450
[45] BUTCHART S H M, WALPOLE M, COLLEN B, et al.Global biodiversity: Indicators of recent declines[J]. Science,2010, 328(5982): 1164–1168
[46] 董翠芳, 梁国付, 丁圣彦, 等. 不同干扰背景下景观指数与物种多样性的多尺度效应——以巩义市为例[J]. 生态学报,2014, 34(12): 3444–3451 DONG C F, LIANG G F, DING S Y, et al. Multi-scale effects for landscape metrics and species diversity under the different disturbance: A case study of Gongyi City[J]. Acta Ecologica Sinica, 2014, 34(12): 3444–3451
[47] 卢训令, 丁圣彦, 游莉, 等. 伏牛山自然保护区森林冠层结构对林下植被特征的影响[J]. 生态学报, 2013, 33(15):4715–4723 LU X L, DING S Y, YOU L, et al. Effects of forest canopy structure on understory vegetation characteristics of Funiu Mountain Nature Reserve[J]. Acta Ecologica Sinica, 2013,33(15): 4715–4723
