乡土风箱果和紫叶风箱果及其杂交种F1叶片的光合功能研究
2018-04-09金微微张会慧滕志远孙广玉
金微微 ,张会慧 ,滕志远 ,孙广玉 ,许 楠 ,3
(1.东北林业大学 生命科学学院,黑龙江 哈尔滨 150040;2.东北农业大学 资源与环境学院,黑龙江 哈尔滨 150030;3.黑龙江省科学院 自然与生态研究所, 黑龙江 哈尔滨 150040)
风箱果Physocarpus amurensisMaxim,蔷薇科风箱果属落叶灌木,树型优美,观赏效果良好[1-2]。由于风箱果花粉活力较弱以及生境的人为破坏等因素导致其分布范围和种群数量日益减少[3-4],目前已经成为我国濒危保护植物之一[5]。紫叶风箱果Physocarpus opulifolius“Diabolo”是我国近年来从北美引入的观赏性花灌木[6],虽然紫叶风箱果与乡土风箱果相比,虽然长势和繁殖具有明显优势,但其抗逆性较差,在我国北方高寒地区露叶栽培中春季返青较慢,且有干稍现象,并且在干旱胁迫下叶片更易失水萎蔫。为获得不但具有紫叶风箱果观赏效果,而且其抗逆能力较强的风箱果品种,黑龙江省森林植物园郁永英等人通过以本地的乡土风箱果(♀)和紫叶风箱果(♂)杂交成功得到88株叶片呈明显紫色的幼苗,自然条件下不但植株长势旺盛,而且冠型整齐一致,表现出明显的杂种优势[7]。
不同品种间植物的光合能力明显不同[8],而杂种优势是生物界普遍存在的生物学现象[9-10],大量的研究均表明杂交种植物的生长、光合能力以及对逆境的适应性均明显优于亲本[11]。叶金山在解释林木杂种优势的“光合优势假说 (Photosynthetic superiority hypothesis)”中认为,林木杂种优势产生的根源就在于杂种与亲本普遍存在的对自然逆境胁迫响应和适应的差异性,而在众多的差异性中又以光合作用的差异性最重要[12]。我们以前的研究中也发现,苏丹草Sorghum sudanense(Piper)Stapf与 高 粱Sorghum bicolor(L.)Moench杂 交获得的新的牧草品种高丹草Sorghum bicolor×S.sudanense的光合能力明显高于苏丹草,并且其在干旱胁迫下的适应能力也明显优于苏丹草[13-14];Campbell等人的研究发现,Ipomopsis aggregate和Ipomopsis tenuttuba的杂交F1代在不同时期的光合能力具有高于亲本的杂种优势[15];Burke等人的研究发现,Iirises fulva和Iirises hexagona的杂交种对逆境的适应性明显优于亲本[16];Whitney等人的研究发现,Helianthus annuus与Helianthus petiolaris杂交后代对逆境的适应能力也较亲本明显提高[17]。为研究3种风箱果叶片的光合能力是否具有杂种优势,本试验从植物的光合能力入手,研究了3种风箱果在自然条件下叶片的光合特性对光强的响应和光合日变化特点,分析了杂种风箱果和2个亲本之间的光合能力之间的差异,拟从光合功能方面揭示杂交风箱果具有杂种优势的内在原因,以期为保护我国的风箱果属植物以及优化外来种的栽培提供一些基础数据。
1 材料与方法
1.1 试验材料与处理
试验于2015年在东北林业大学植物生理实验室进行,此时我国北方地区植物处于旺盛生长期。试验材料为2013年春季黑龙江省森林植物园提供的1年生紫叶风箱果、乡土风箱果和杂交风箱果,其中杂交风箱果为选择乡土风箱果(♀)×紫叶风箱果(♂)F1杂交种枝条进行扦插获得的扦插苗。3种试材株高0.3~0.5 m,主分枝数5~10个。幼苗植于开口直径28 cm、底直径15 cm、高20 cm的塑料花盆中,每盆定植1株,培养基质为草碳土,期间正常浇水及施肥,冬季室外自然越冬。自然环境下培养3 a,待表型稳定后,于2015年8月1日植株旺长期进行光合日变化和光响应曲线的测定。
1.2 测定项目和方法
光合气体交换参数日变化的测定:选择长势相对一致的桑树幼苗主干上的倒数第3~4片完全展开叶,利用Li-6400光合测定系统于上午5:00开始到下午19:00(共计16 h)每2 h测定一次不同品种风箱果叶片的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞间CO2浓度(Ci),各处理均测定3次重复。测定时光合仪叶室垂直对准太阳方向,以保证不同处理间光合有效辐射值的稳定和准确,并用光合测定系统分别记录大气温度和光合有效辐射等参数。然后计算1 d之中光合同化总量,方法参照张会慧等[18]。光合日变化测定时的光合有效辐射和大气温度变化如图1所示,其中,1 d之中光合有效辐射和大气温度的变化均呈单峰曲线变化,其光合有效辐射的峰值出现在中午11:00左右,其最大值大约为 1 500 μmol·m-2s-1,之后逐渐降低,在傍晚19:00时光合有效辐射降低到100 μmol·m-2s-1以下。与光合有效辐射不同,1 d之中大气温度的日变化幅度相对较小,并且出现最高温度的时间较光合有效辐射明显推迟,最高温度的时间点出现在下午14:00左右。
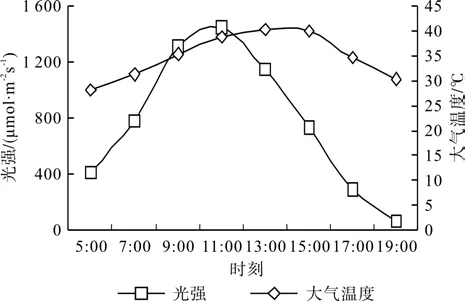
图1 大气温度和光合有效辐射的日变化曲线Fig.1 Diurnal change of PAR and temperature
光合气体交换参数的光响应曲线测定:利用Li-6400光合测定系统,将仪器自带光源分别设定光强(PFD)为1 800、1 500、1 200、1 000、800、500、200、100、50 和 0 μmol·m-2s-1,按照光强由高到低的顺序分别测定不同Cd浓度处理下烟草从上往下数第2片完全展开功能叶的Pn、Gs和Tr,并分别绘制Pn-PFD、Gs-PFD和Tr-PFD响应曲线,用直角双曲线模型求得Pn-PFD响应曲线表观量子效率(Qaqy)、暗呼吸速率(Rd)、光补偿点(Lcp)、光饱和点(Lsp)、光饱和时的净光合速率(Pmax)。
1.3 数据处理方法
运用 Excel(2003)和 SPSS(7.05)软件进行数据统计分析,采用单因素方差分析(One-way ANOVA)和最小显著差异法(LSD)比较不同数据组间的差异。
2 结果与分析
2.1 植株和叶片的表型特征

图2 3种风箱果的长势和叶片颜色差异Fig.2 Growth and leaf color differences of three Physocarpus seedlings
由图2可以看出,紫叶风箱果植株的株高和冠幅度明显高于乡土风箱果,即紫叶风箱果具有明显的生长优势。而杂交风箱果虽然株高不及紫叶风箱果,但其整体长势与紫叶风箱果相差不大,其枝条较粗,长势较旺,长势明显优于乡土风箱果。乡土风箱果和紫叶风箱果杂交获得的F1代杂交风箱果不但在观赏效果上具有紫叶风箱果的特征,而且其生长特性明显优于乡土风箱果。紫叶风箱果叶片呈深紫色,风箱果叶片呈绿色,而乡土风箱果(♀)和紫叶风箱果(♂)杂交获得的F1代杂交风箱果叶色呈二者中间色。紫叶风箱果叶片呈现深裂特征,乡土风箱果叶片裂痕较浅,而杂交风箱果的裂叶特征更接近于紫叶风箱果。杂交风箱果的叶色、叶片形态和植株长势更接近于紫叶风箱果(♂),即受父本遗传的影响较大。
2.2 净光合速率和气孔导度日变化曲线
由图3可以看出,3种风箱果叶片的各光合气体交换参数的日变化曲线基本相似,3种风箱果之间有明显差异。3种风箱果叶片的Pn和Gs均表现为明显的双峰变化曲线,2个峰值分别出现在9:00和15:00,2个参数上午时的峰值均明显大于下午的峰值,特别是Gs这种现象更为明显,Pn和Gs的谷值出现在11:00—13:00之间,即中午时气孔明显关闭,表现出明显的光合午休现象。但是乡土风箱果叶片Pn和Gs在出现光合午休后下午时基本无明显恢复。3种风箱果比较,1 d之中乡土风箱时叶片的Pn和Gs整体低于紫叶风箱果和杂交风箱果,而除9:00和17:00时杂交风箱果叶片的Pn稍高于紫叶风箱果外,其它各时间点杂交风箱果叶片的Pn和1 d之中杂交风箱果叶片的Gs均明显低于紫叶风箱果。
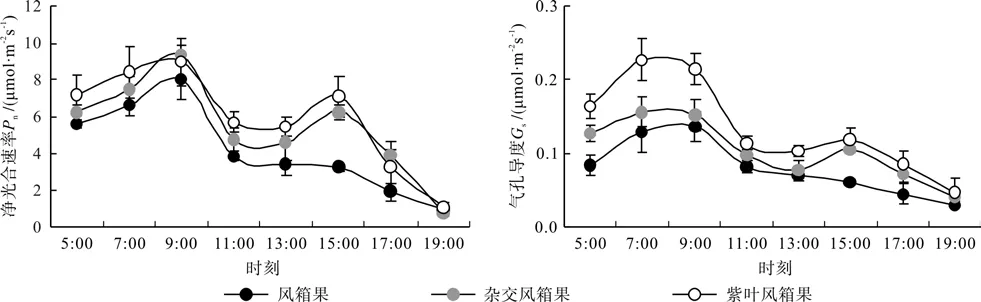
图3 3种风箱果叶片Pn和Gs日变化曲线Fig.3 Diurnal change of Pn and Gs diurnal variation of in leaves of 3 Physocarpus seedlings
2.3 蒸腾速率和胞间CO2浓度日变化曲线
由图4可以看出,3种风箱果叶片的Tr1 d之中也整体呈双峰变化曲线,但其中午时的谷值降低幅度明显小于Pn和Gs,乡土风箱果和紫叶风箱果叶片的Tr谷值均出现在11:00时,而杂交风箱果Tr谷值则出现在13:00时,1 d之中3种风箱果叶片的Tr值在各时间点整体表现为紫叶风箱果>杂交风箱果>乡土风箱果。3种风箱果叶片的Ci在1 d之中的变化幅度较小,上午5:00—9:00时呈降低趋势,但在中午11:00时有小幅增加,随后到13:00时逐渐降低,13:00之后随着时间推移均有小幅度的增加。1 d之中3种风箱时叶片的Ci之间差异也较小,其紫叶风箱果叶片的Ci稍高于乡土风箱果和杂交风箱果,杂交风箱果相对最低。

图4 3种风箱果叶片Tr和Ci日变化曲线Fig.4 Diurnal change of Tr and Ci diurnal variation of in leaves of three Physocarpus seedlings
2.4 1 d之中光合同化总量
由图5可以看出,杂交风箱果和紫叶风箱果叶片1 d之中光合同化总量分别高于乡土风箱果30.79%(P<0.01)和41.23%(P<0.01),均达极显著差异水平,杂交风箱果叶片1 d之中光合同化总量虽然低于紫叶风箱果7.38%(P>0.05),但二者无显著差异。
2.5 净光合速率光响应曲线
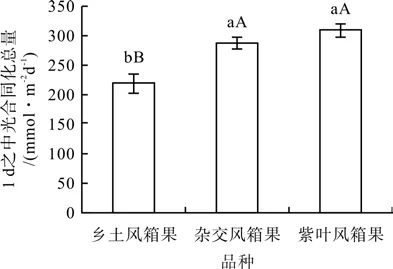
图5 3种风箱果叶片1 d之中光合同化总量Fig.5 Diurnal assimilation amounts in leaves of 3 Physocarpus seedlings
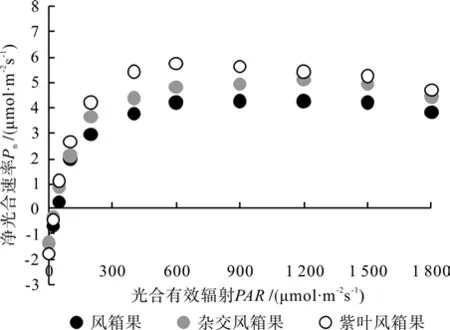
图6 3种风箱果叶片的净光合速率光响应曲线Fig.6 Net photosynthetic rate to illumination intensity in leaves of three Physocarpus seedlings
由图6可以看出,不同光强下3种风箱果叶片的Pn整体表现为紫叶风箱果>杂交风箱果>乡土风箱果,但是在光强增加到900 μmol·m-2s-1后,紫叶风箱果和杂交风箱果叶片的Pn差异程度逐渐变小。通过光响应曲线的拟合分析结果可以看出(见表1),紫叶风箱果叶片的Qaqy和Rd分别高于乡土风箱果32.35%和32.59%,分别高于杂交风箱果25.01%和31.62%,乡土风箱果和杂交风箱果叶片的Qaqy和Rd差异不大。杂交风箱果和紫叶风箱果叶片的Lcp分别较乡土风箱果低13.06%和13.41%,而Lsp则分别较乡土风箱果高31.72%和31.29%,并且Pmax也分别高于乡土风箱果17.33%和27.40%。
2.6 气孔导度和蒸腾速率光响应曲线
由图7可以看出,3种风箱果叶片的Gs和Tr的变化趋势大致相似,不同光强下紫叶风箱果和杂交风箱果叶片的Gs和Tr均明显大于乡土风箱果,在低于 900 μmol·m-2s-1光强下,紫叶风箱果叶片的的Gs和Tr明显高于杂交风箱果,但当光强超过900 μmol·m-2s-1后,杂交风箱果叶片的Gs和Tr增强幅度明显大于紫叶风箱果,并且二者差异幅度也随着光强的增加而变大,即杂交风箱果叶片的Gs和Tr随着光强的增加其增加幅度最大。
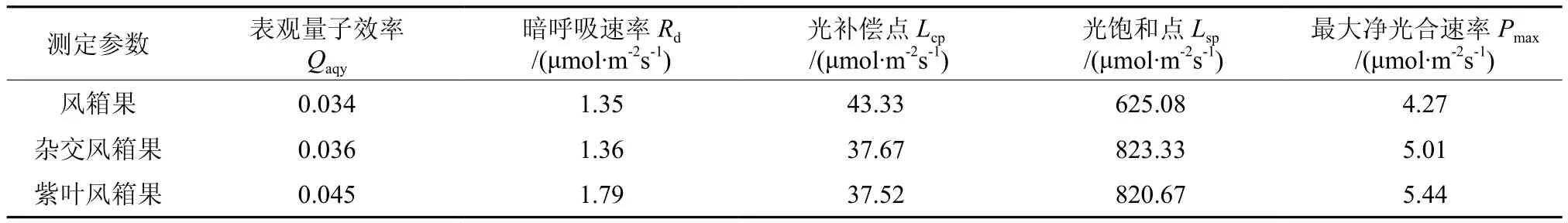
表1 3种风箱果叶片的光合参数Table 1 Photosynthetic parameters in leaves of three Physocarpus seedlings

图7 3种风箱果叶片的孔导度和蒸腾速率光响应曲线Fig.7 Stomatal conductance and transpiration rate to illumination intensityin leaves of three Physocarpus seedlings
3 讨 论
植物的光合能力直接影响着植物同化物的积累,不但直接决定着植物的生长而且与其抗逆能力直接相关[19]。杂种优势是生物界普遍存在的生物学现象,较多的研究发现,杂交种植物的光合能力明显优于亲本。刘元芝等[20]的研究发现,玉米Zea maysL.杂交种净光合速率和气孔导度的光合日变化曲线中各时间段里均明显高于亲本,光合能力表现出明显的杂种优势。在低光强下,杂交泡桐PaulowniaSieb. et Zucc.表现出明显的光合优势[21],周连杰等[22]的研究也发现,小麦Triticum aestivumL.小偃54和8602及其杂交后代小偃81的光合特性也更能适应黄淮地区小麦生育后期光照不足和连阴骤晴的不良气候。许晨璐等[23]的研究也发现,长白落叶松Larix olgensis和日本落叶松Larix kaempferi的杂交种的净光合速率值也显著高于亲本。本试验结果表明,3种风箱果叶片的光合气体交换参数Pn、Gs和Tr均表现为明显的双峰变化曲线,其两个峰值均出现在9:00和15:00,而谷值则出现在12:00左右。说明3种风箱果均存在明显的光合“午休”现象,关于光合“午休”的产生机理,国内外学者进行了广泛的研究,许大全等[24]的研究认为,光合“午休”是由于中午时的强光和空气湿度降低导致叶片气孔的关闭等气孔因素,以及PSⅡ活性的降低和光合作用相关酶活性降低等非气孔因素的共同限制,另外,中午高光强下的光呼吸作用增强也是植物光合“午休”的重要原因之一[25]。本试验中,在中午时的光强达到了 1 500 μmol·m-2s-1,而温度甚至达到了35 ℃以上,说明中午时的高温强光是导致3种风箱果光合“午休”的重要原因之一,高温和强光下,3种风箱果叶片为减少水分的过度蒸腾,气孔导度的降低导致了CO2的供应受到限制。但是本试验中,在中午时3种风箱果叶片的Ci却有小幅度的增加,根据Farquhar等[26]的气孔因素分析结果,认为3种风箱果叶片光合午休的原因还可能与非气孔因素的作用有关,可能是高温强光下光系统活性的降低等原因造成的。这与张晓慧等[27]在马蹄金Dichondra repensForst.上的研究结果相似。3种风箱果比较,1 d之中乡土风箱时叶片的Pn、Gs和Tr整体低于紫叶风箱果和杂交风箱果,并且不同时间段的紫叶风箱果叶片的Pn、Gs和Tr相对高于杂交风箱果,并且紫叶风箱果叶片的Ci在各时间段也稍高于另外2个品种,说明引种紫叶风箱果的光合能力明显高于乡土风箱果品种,表现为株高和冠幅度明显高于乡土风箱果,即紫叶风箱果具有明显的生长优势(图2)。乡土风箱果和紫叶风箱果杂交种不但在观赏效果上具有紫叶风箱果的特征,而且其光合能力也明显高于乡土风箱果品种,并且表现为1 d之中光合同化总量也明显高于乡土风箱果,即杂交风箱果的叶色和光合能力主要受父本紫叶风箱果的影响,这与云南松Pinus yunnanensis与油松Pinus tabuliformis自然杂交产生的同倍体杂种高山松Pinus densata的光合能力表现出明显的杂交优势,并且主要受父系遗传性状的影响的结果相似[28-29]。
植物光合气体交换参数的光响应曲线反映了植物对光能的利用能力[30]。本试验中,不同光强下3种风箱果叶片的Pn整体表现为紫叶风箱果>杂交风箱果>乡土风箱果,即紫叶风箱果的光合能力明显高于乡土风箱果,而2种风箱果杂交F1代风箱果叶片的光合能力较乡土风箱果明显提高,其光合能力介于父本和母本之间,明显高于母本乡土风箱果,这与其光合日变化的结果相符。但是当光强超过900 μmol·m-2s-1后,紫叶风箱果和杂交风箱果叶片的Pn差异程度逐渐变小。并且杂交风箱果叶片的Gs和Tr增强幅度明显大于紫叶风箱果,说明虽然紫叶风箱果叶片的光合能力明显高于杂交风箱果,但是在高光强下,杂交风箱果的光合能力达到与紫叶风箱果接近水平。Qaqy是植物对光能利用效率的重要指标,反映低光强下植物的光合能力[31],Pmax体现了植物潜在的光合能力,Lcp和Lsp反映了植物对光能的利用能力[32-33]。本试验中,杂交风箱果叶片的Qaqy与乡土风箱果相关不大,但明显低于紫叶风箱果,而杂交风箱果叶片的Lcp低于乡土风箱果,而Lsp则明显高于乡土风箱果,说明杂交风箱果在光合能力方面的优势在弱光下并没有明显表现出来,但在强光下的光合能力却较乡土风箱果明显提高,最终表现出杂交风箱果叶片的Pmax明显高于乡土风箱果,其原因可能是由于杂交风箱果叶片中含有一定量的花色素苷,对光具有一定的过滤或衰减作用[34-35],导致其叶片对弱光的捕获能力受到一定的限制,而紫叶风箱果叶片虽然具有较高的花色素苷含量,但对弱光的利用能力却较强,这可能与其特殊的解剖结构或光系统功能有关,这有待更进一步的研究。
4 结 论
引种紫叶风箱果叶片不但具有紫色的观赏效果,而且其光合能力也明显优于乡土风箱果,而乡土风箱果(♀)×紫叶风箱果(♂)F1杂交种的叶色表现、长势和光合能力均受父本遗传特性的影响较大,其光合能力明显优于乡土风箱果(♀),表现为光合日变化的光合午休和1 d之中光合同化总量明显较乡土风箱果提高,并且对光能的利用能力也明显提高,但是杂交风箱果叶片的光合能力仍不及紫叶风箱果(♂)。
参考文献:
[1]Zhang HH, Zhong HX, Sui X,et al.Adaptive changes in chlorophyll content and photosynthetic features to low light inPhysocarpus amurensismaxim andPhysocarpus opulifolius“diabolo” . Peer J, 2016,4(3):e2125
[2]周以良,董世林,聂绍荃.黑龙江树木志[M].哈尔滨:黑龙江科学技术出版社,1986:284-286.
[3]殷东生,魏晓慧,沈海龙.风箱果的花部综合特征及繁育系统研究[J].北京林业大学学报,2016,38(1):67-73.
[4]魏晓慧,殷东生,沈海龙.风箱果种子催芽技术研究[J].植物研究,2013,33(6):709-712.
[5]殷东生,沈海龙,兰士波.风箱果花粉生活力、柱头可授性及传粉者的观察东[J].北京林业大学学报,2010,38(4):80-81,113.
[6]刘晓东,于 晶.紫叶风箱果叶片花色素苷的提取及其稳定性[J].东北林业大学学报,2011,39(2):38-39,81.
[7]郁永英,张华艳,潘 杰,等.风箱果属植物杂交育种[J].东北林业大学学报,2010,38(7):16-18.
[8]董胜君,尹 健,刘明国,等.山杏新品种光合生理特性研究[J].经济林研究,2016,34(2):67-72.
[9]Prasad TK, Anderson MD, Martin BA,et al.Evidence for Chilling-Induced Oxidative Stress in Maize Seedlings and a Regulatory Role for Hydrogen Peroxide[J].The Plant Cell, 1994,6(1):65-74.
[10]於朝广,殷云龙.落羽杉属杂交良种”落羽杉中山302”和”中山杉118”[J].林业科学,2010,46(5):181-182.
[11]吕佳斌,谭晓风,李 泽,等.油桐二代优树及控制授粉子代光合特性的比较研究[J].经济林研究,2016,34(4):7-13.
[12]叶金山,王章荣.杂种马褂木杂种优势的遗传分析[J].林业科学,2002,38(4):67-71.
[13]张会慧,张秀丽,胡彦波,等.高粱-苏丹草杂交种的生长特性和光合功能研究[J].草地学报,2012,20(5):881-887.
[14]胡举伟,张会慧,张秀丽,等.高丹草叶片PSⅡ光化学活性的抗旱优势[J].草业科学,2015,32(3):392-399.
[15]Campell D R, Galen C, Wu C A. Ecophysiology of first second generation hybrids in a natural plant hybrid zone[J]. Oecologia,2005, 144(2):214-225.
[16]Burke J M, Carney S E, Arnold M L. Hybrid fitness in the Louisiana Irises: analysis of parental andF1performance[J].Evolution, 1998,52(1):37-43.
[17]Whitney KD, Randell RA, Rieseberg L H. Adaptive introgression of abiotic tolerance traits in the sun flowerHelianthus annuus[J].New Phytologist,2010,187(1):230-239.
[18]张会慧,张秀丽,胡彥波,等.盐碱地施用农家肥和化肥对桑树生长和叶片光合日变化的影响[J].土壤, 2013, 44(3):444-450.
[19]张会慧,赵 莉,许 楠,等.间作模式下桑树和大豆叶片光合日变化特点[J].经济林研究,2011,29(1):21-26.
[20]刘元芝,张洪钧,徐 侃,等.玉米杂交种及其亲本光合特性的比较研究[J].玉米科学2009,17(3):71-75.
[21]王 楠,李芳东,叶金山.光强对杂交泡桐光合生理和荧光特性的影响[J].生态学杂志, 2015,34(11):3118-3124.
[22]周连杰,谢彦庆,徐 文,等.去除遮阴后自然强光对小偃54和8602及其杂交后代光合特性的影响[J].作物学报,2015(5):813-819.
[23]许晨璐,孙晓梅,张守攻.日本落叶松与长白落叶松及其杂种光合特性比较[J].北京林业大学学报,2012,34(4):62-66.
[24]许大全.光合作用效率[M].上海:上海科学技术出版社, 2002.
[25]Salvucci M E, Crafts-Brandner S J. Inhibition of photosynthesis by heat stress: the activation state of Rubisco as a limiting factor in photosynthesis [Review][J]. Physiologia Plantarum, 2009,44(2):S35.
[26]Farquhar G D, Sharkey T D. Stomatal conductance and photosynthesis[J].Annual Review of Plant Physiology,1982,33(3): 317-345.
[27]张晓慧,干友民,任 婷,等.5份西南野生马蹄金材料的光合特性比较[J].草业科学,2010,27(3):76-81.
[28]毛建丰,李 悦,刘玉军,等.高山松种实性状与生殖适应性[J].植物生态学报,2007,31(2):291-299.
[29]高 琼,王维有,孟景祥,等.油松×云南松杂种与亲本种和高山松的光合特性比较[J].北京林业大学学报,2016,38(2):37-43.
[30]张会慧,张秀丽,胡彦波,等.中碱钠盐胁迫对桑树幼苗生长及光合特性的影响[J].南京林业大学学报,2013,37(1):55-60.
[31]黄 俊,郭世荣,吴 震,等.6个不结球白菜品种光合作用特性的研究[J].西北植物学报,2006,26(6):1183-1189.
[32]刘悦秋,孙向阳,王 勇,等.遮荫对异株荨麻光合特性和荧光参数的影响[J].生态学报,2007,27(8):3457-3464.
[33]刘高慧,李 昆,孙永玉,等.瓣石斛光合特性研究[J].林业科学研究,2014,27(2):265-269.
[34]Chalker-Scitt L. Environmental significance of anthocyanins in plant stress responses[J]. Photochemistry and Photobiology,1999(70): 1-9.
[35]田 野,张会慧,王 娟,等.遮荫对紫叶李叶片花色素苷含量和PSⅡ功能的影响[J].中南林业科技大学学报,2013,33(6): 51-56.
