干旱胁迫对不同苦荞品种苗期生长和根系生理特征的影响
2018-04-08路之娟张永清
路之娟,张永清,2*,张 楚
(1 山西师范大学 生命科学学院,山西临汾 041004;2 山西师范大学 地理科学学院,山西临汾 041004)
随着人们生活需求由温饱型向营养型乃至保健型的转化,苦荞的“三降一疗两通”食疗营养功能日益受到消费者的青睐[1-2]。苦荞(FagopyrumtataricumL.)为蓼科(Polygonaceae)双子叶植物,是起源于中国西南部的一种古老的杂粮作物。其含有丰富的矿物质、维生素、脂肪、蛋白质和人体所需的微量元素[3-4],更具备其他植物所不具有的黄酮类化合物,使其药食兼用的作用更为突出,因此常被称为“五谷之王”、“三降食品”。苦荞生育期短,耐冷凉、耐瘠薄、适应性强,在黄土高原地区具有明显的区位优势和生产优势,是当地特色农业发展的重要杂粮作物。
相关研究表明,全球因水源匮乏而导致的减产超过了其他自然灾害造成减产数量的总和[5]。山西素有“十年九旱”之说,干旱是影响该省农业高产稳产的重要逆境因素[6]。根系是作物感受土壤干旱的原初部位与敏感部位,其数量大小、分布和生理状况等直接影响着作物抗旱性的强弱,因而研究根系对干旱胁迫的生理生态反应,对于揭示作物抗旱的本质更有意义[7]。研究苦荞根系对水分胁迫的响应特征,探明其抗旱机理与调控对策,对实现山西黄土高原丘陵区旱地农业可持续发展及保障粮食安全具有重要的意义。现有关于苦荞的研究多集中于种质资源[8]、农艺性状[9-10]、药用与营养价值[11-12]、栽培技术[13-14]、养分吸收与施肥的增产作用[15-17]、逆境胁迫对苦荞化学成分及生理指标影响等方面[18-22],而对苦荞的抗逆性机理则研究较少,尤其是关于干旱胁迫对苦荞根系生理特性的影响更是鲜有报道。本试验拟以前期筛选的不同耐旱性苦荞品种为材料,通过盆栽控水试验研究不同水分条件下不同耐旱性苦荞品种的苗期生长及根系生理生态状况,比较不同抗旱性品种间的变化差异,探明与苦荞抗旱性密切相关的生理生态指标,并最终为黄土高原苦荞抗旱品种的种植、引种、选育及抗旱机制等方面的研究提供理论依据。
1 材料和方法
1.1 试验材料
试验材料为经前期预实验筛选出的耐旱苦荞品种‘迪庆苦荞’、‘西农9909’和不耐旱品种‘西荞1号’、‘黑丰1号’。4个品种生育期无明显差异,均为105 d左右。
1.2 试验处理
试验于2017年5~6月在山西师范大学塑料大棚内进行。试验采用二因素完全随机设计,因素A为不同耐旱性苦荞品种:‘迪庆苦荞’、 ‘西农9909’、 ‘西荞1号’和‘黑丰1号’;因素B为不同水分处理:正常供水(土壤含水量为田间持水量的65%~75%,CK) 、中度干旱(土壤含水量为田间持水量的45%~55%,MS)和重度干旱(土壤含水量为田间持水量的25%~35% ,SS),共12个处理组合,每个处理重复3次,每天采取称重法浇水。
采用砂培盆栽试验,营养钵大小为16 cm×14 cm,每盆装入1.5 kg细沙,沙子使用前用去离子水洗净、晾干备用。每盆精选籽粒饱满均匀且无病虫害的种子14粒,播后正常供应水分,幼苗2叶1心后间苗,每盆定苗7株。3叶1心后开始进行各水平的水分处理,为了保证试验期间幼苗生长不受养分限制,每盆定量浇灌2.5 L相同浓度的霍格兰营养液,其成分及含量为:Ca(NO3)2820.7 mg·L-1、KNO3505.6 mg·L-1、MgSO47H2O 616.2 mg·L-1、KH2PO4272.2 mg·L-1,微量元素和铁盐同霍格兰标准营养液,每5 d每盆浇灌同等营养液1次,处理20 d后(显蕾期)开始测定相关指标。
1.3 测定指标与方法
1.3.1形态指标叶面积用叶面积仪法测定;株高、最大根长用直接测量法;生物量用烘干后称重法;根系的总长、根总表面积、根体积用英国产 Delta-T SCAN根系分析系统测定;叶片相对含水量(RWC)用称重法测定[23]。
1.3.2生理指标叶绿素含量用分光光度计测定[23];用美国 Opti2science公司生产的 OS52FL调制式叶绿素荧光仪于取样前一天早上(5:00~8:00,弱光环境下) 测定最上一片定型叶的叶绿素荧光参数初始荧光Fo和最大荧光产量Fm,并计算可变荧光Fv(Fm-Fo)、PSⅡ最大光化学量子产量Fv/Fm。超氧化物歧化酶(SOD)活性用蓝四唑法测定[23];过氧化物酶(POD)活性用愈创木酚比色法测定[23];丙二醛(MDA)含量用硫代巴比妥酸法测定[23];可溶性糖含量用蒽酮比色法测定[23];可溶性蛋白质含量用考马斯亮蓝G-250染色法测定[23]。以上指标均重复3次,取平均值。
1.4 品种苗期抗旱性综合评价
采用模糊数学隶属函数法[24]对苦荞品种苗期的抗旱性进行综合评价。设有n个苦荞品种(n=1,...,i), 每个苦荞品种有m个抗旱性指标(m=1,...,j),Dij和Wij分别表示第i个苦荞品种、第j个抗旱指标的隶属度值和权重系数。
(1)标准化转化原始数据:
抗旱指标与抗旱性呈正相关时:
u(xj)=(xj-xmin)/(xmax-xmin)
(1)
抗旱指标与抗旱性呈负相关时:
u(xj)=1-(xj-xmin)/(xmax-xmin),j=1, 2,...,n
(2)
(2)权重系数(Wj)计算:
(3)
(4)
(3)隶属函数值(D)计算:
(5)
式中,u(x)表示数据标准化值,xmax表示第j个指标的最大值,xmin表示第j个指标的最小值,Vj代表标准差系数, 比较品种最终所得抗旱隶属度值。
1.5 数据分析
用 Microsoft Excel 2003和SPSS 19.0 软件对试验数据进行处理分析,采用Duncan’s法进行多重比较,用SigmaPlot 10.0 作图。
2 结果与分析
2.1 干旱胁迫对苦荞幼苗植株幼苗生长的影响
2.1.1地上部生长在干旱胁迫条件下,各苦荞品种苗期株高、茎粗、叶面积及地上部干重均显著降低(P<0.05),且随着胁迫程度的加剧下降幅度也更大(表1)。其中,与正常供水处理(CK)相比,苦荞幼苗株高、茎粗、叶面积及茎叶干重在两个干旱胁迫环境下(MS和SS) 分别平均显著降低了21.25%、17.00%、35.13%和36.62%,即干旱胁迫对上述生长指标的影响表现为茎叶干重>叶面积 >株高>茎粗。同时,不同耐旱型苦荞品种之间相比较,重度(SS)和中度(MS)干旱胁迫下各苦荞品种的上述指标虽均比正常供水处理显著降低,但耐旱品种的平均降幅均小于不耐旱品种,且差异显著(P<0.05)。以上结果说明耐旱性苦荞品种在干旱环境中表现出更强的地上部生长优势,受干旱胁迫的影响相对较小,适应性更强。
2.1.2根系生长在干旱胁迫条件下,各苦荞品种幼苗的根系体积和根系表面积、根系干重均比正常水分处理(CK)显著降低(P<0.05),且随着干旱胁迫程度的增加,降幅也随之加大;与耐旱性品种相比较,不耐旱品种根系相关指标降低的幅度更大(表 2)。在两个干旱环境中,耐旱品种根系体积、根系表面积和地下部干重的平均降幅分别为31.00%、21.50%和31.25%,不耐旱品种的相应降幅分别为51.50%、47.50%和42.75%。同时,与CK处理根冠比相比,仅耐旱品种‘迪庆苦荞’在重度胁迫(SS)下显著上升了7.00%,其余品种在两种干旱胁迫下均无显著变化。另外,与CK主根长相比,两种耐旱型品种在中度干旱胁迫(MS)下显著降低,而在SS下显著增加,但不耐旱品种则在2种干旱胁迫下均显著下降,且在SS胁迫下降幅更大。可见,干旱条件下,耐旱型苦荞在一定程度上可通过促进主根伸长来增加根系吸收范围,而不耐旱品种适应性较弱,表现为主根长长度持续降低;干旱胁迫使‘迪庆苦荞’苦荞根冠比有所升高,其通过增加根冠比增强了干旱适应性。
2.2 干旱胁迫对苦荞幼苗叶片生理指标的影响
2.2.1叶绿素含量和叶片相对含水量由表3可知,4个苦荞品种叶片叶绿素含量在两个干旱胁迫 (MS、SS)下均比CK显著下降(P<0.05)。其中,不耐旱品种‘黑丰1号’叶绿素含量降幅最大,分别达31.00%和58.00%,而品种‘西荞1号’次之,耐旱品种‘西农9909’和‘迪庆苦荞’重度胁迫下降幅均仅为25%。叶片相对含水量同叶绿素含量的变化趋势相似,‘迪庆苦荞’、‘西农9909’、‘西荞1号’和‘黑丰1号’在中度胁迫下分别比CK下降2%、6.00%、19.00%和19.00%,在重度胁迫下分别降低了22.00%、25.00%、30.00%和29.00%。可见,耐旱品种叶绿素含量和叶片含水量降幅小于不耐旱品种,其在干旱环境中仍能保持相对较高的叶绿素含量和含水量,从而保证自身有较强的光合能力,同时较高的含水量维持了体内正常的生理代谢活动,表现出较强抗旱性。
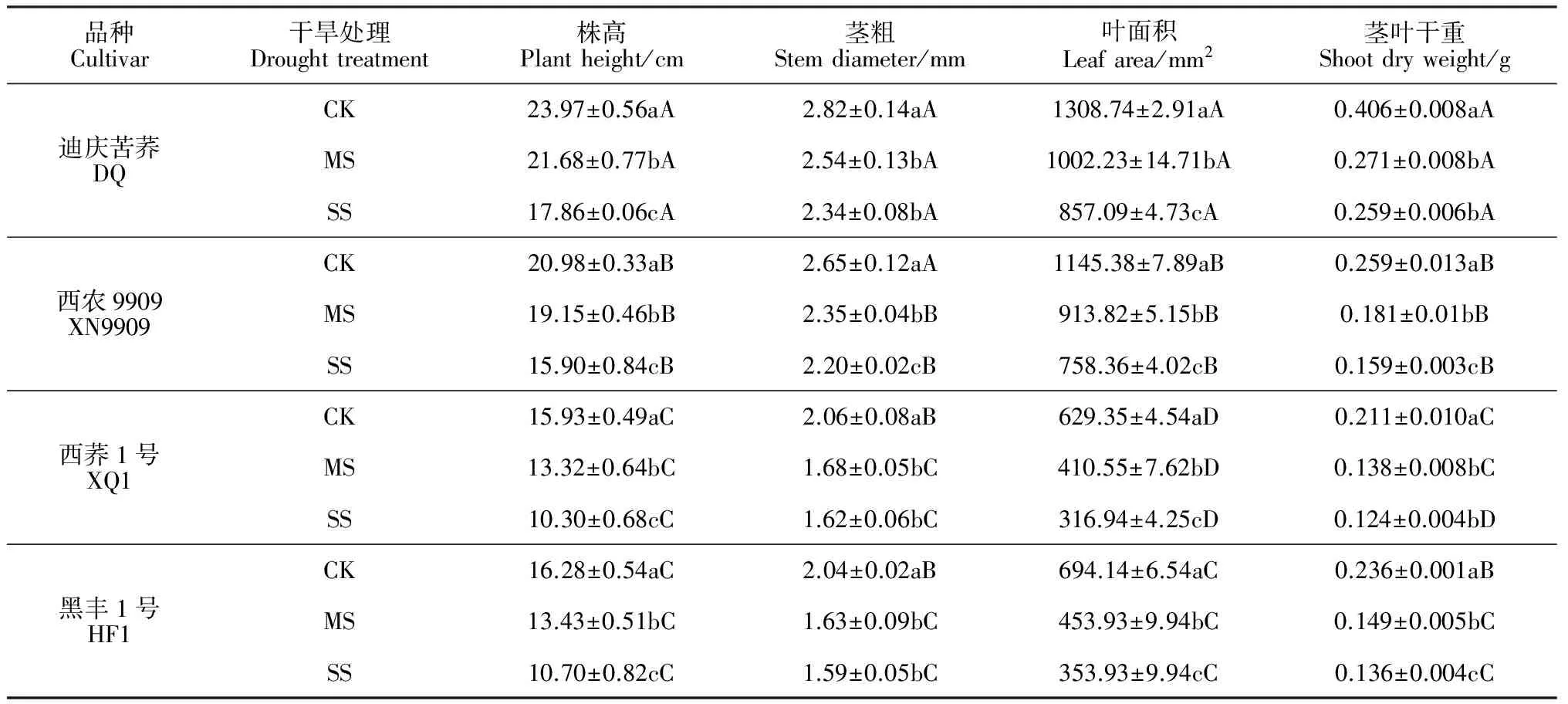
表1 水分胁迫条件下苦荞苗期地上部植株生长的变化
注:CK.对照;MS.中度胁迫;SS.重度胁迫;表内数据为平均值±标准误;同列数据后不同大、小写字母分别表示品种间或处理间差异达0.05显著水平(P<0.05)。下同
Notes: CK. Normal irrigation; MS. Moderate water stress; SS. Heavy water stress. Values are mean ± SE. Different capital or normal letters in the same column indicate significant difference among different cultivars or treatments at 0.05 level. The same as below
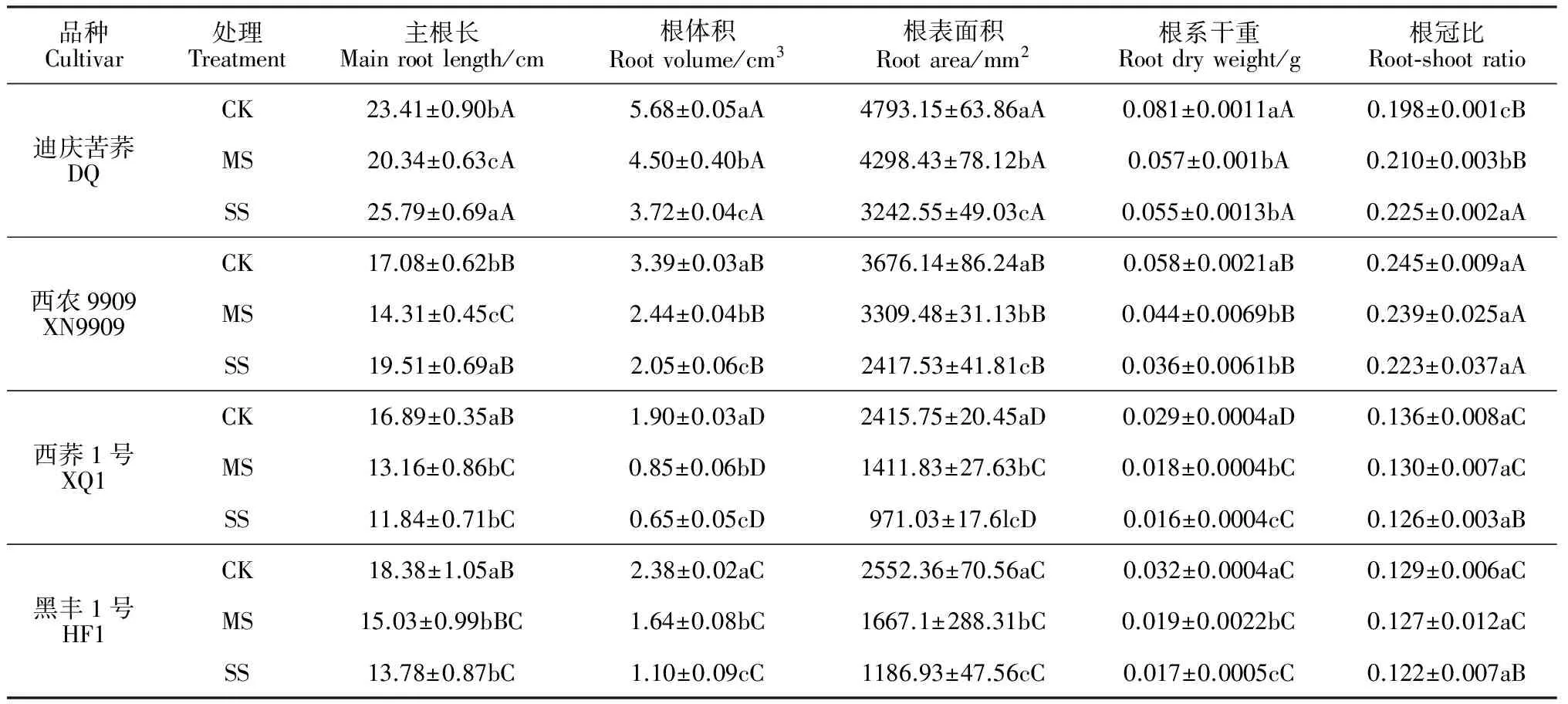
表2 水分胁迫对苦荞苗期根系形态指标的影响
2.2.2叶片荧光参数各苦荞品种叶片的初始荧光产量(F0)干旱胁迫下均比CK有不同程度的增加,且胁迫越重增加幅度越大,不耐旱品种增幅在两种干旱胁迫下均达到显著水平,耐旱品种在重度胁迫下也达到显著水平(P<0.05);在相同程度胁迫下,耐旱品种F0增幅小于不耐旱品种(表3);同时,干旱胁迫使各品种苦荞Fm、Fv/Fm显著降低,胁迫程度越重降幅越大,且不耐旱品种降幅大于耐旱品种。说明干旱胁迫对各品种光合系统Ⅱ(PSⅡ)反应中心均造成不同程度破坏,但耐旱品种反应中心受破坏程度较小,抗旱能力更强。
2.3 干旱胁迫对苦荞幼苗根系生理指标的影响
2.3.1根系活力干旱胁迫使各品种苦荞苗期根系活力显著下降,且随着胁迫程度的增大下降更明显,而耐旱品种的根系活力降幅又小于不耐旱品种(表4)。其中,与CK处理相比,耐旱品种根系活力在2个干旱胁迫 (MS和SS)下平均降幅是22.50%,而不耐旱品种则平均降低26%。可见,耐旱苦荞品种根系活力明显高于不耐旱品种,对干旱的耐性更强,吸收能力更好。
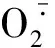
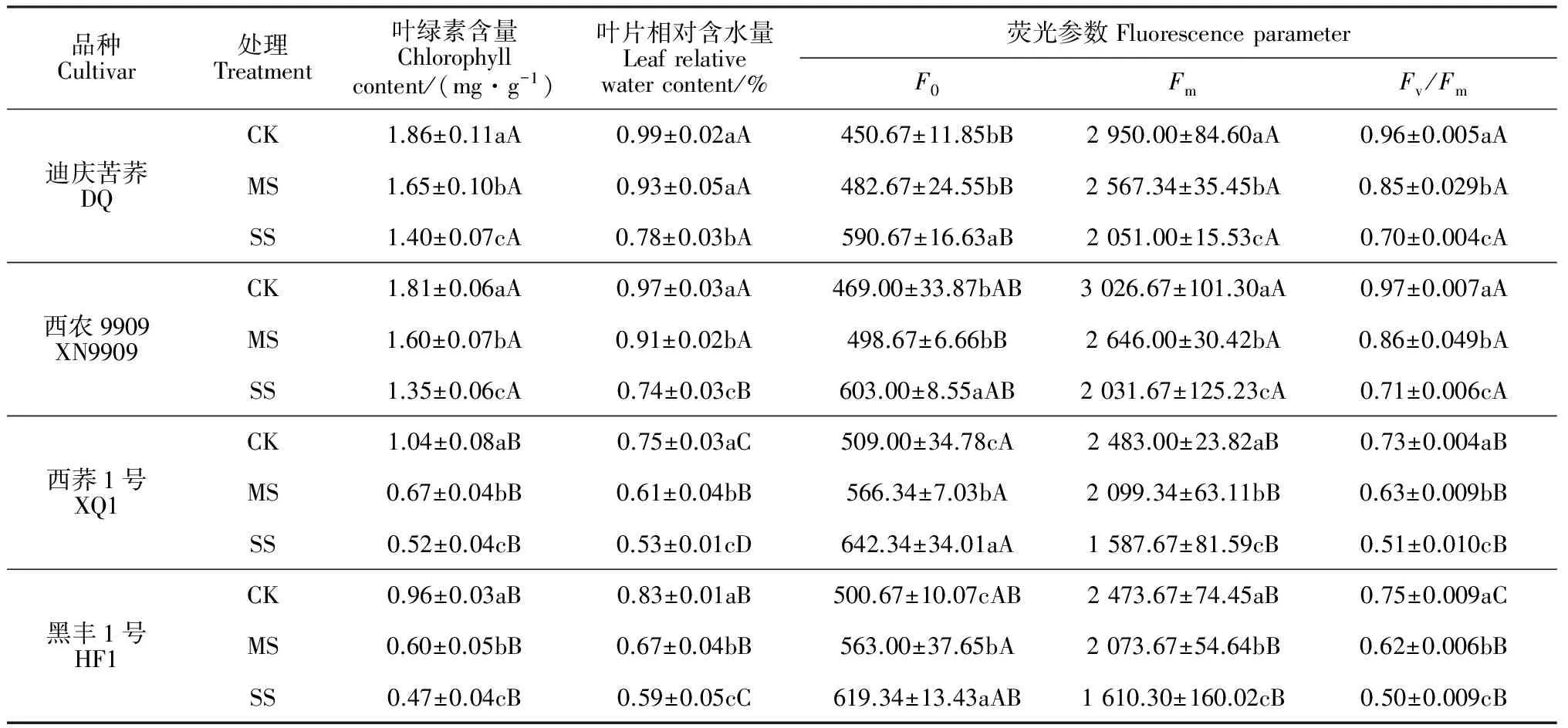
表3 水分胁迫对苦荞苗期叶片叶绿素含量、叶片相对含水量、荧光参数的影响
2.3.3渗透调节物质含量植株在逆境胁迫下会通过累积渗透调节物质降低渗透势来适应环境,而可溶性糖、可溶性蛋白和游离脯氨酸作为植物体内重要的渗透调节物质均可作为衡量植物抗旱性的指标。表5显示,干旱胁迫对各品种苦荞苗期根系可溶性糖、可溶性蛋白及游离脯氨酸含量均有不同程度影响,可溶性糖及游离脯氨酸含量显著升高,而可溶性蛋白含量显著降低。其中,在中度和重度干旱胁迫下,根系可溶性蛋白含量降幅最大的品种均是‘西荞1号’,降幅分别达16%和26%,随后依次是品种‘黑丰1号’、 ‘西农9909’和‘迪庆苦荞’,两个耐旱品种降幅显著小于两个不耐旱品种;在两个干旱胁迫下,耐旱品种的平均可溶性糖含量和游离脯氨酸含量分别较CK处理显著提高71%和86%,而不耐低旱品种分别显著提高21%和42%,耐旱品种的增幅均明显高于不耐旱品种。由此表明,在干旱胁迫环境下,耐旱苦荞品种能够比不耐旱品种保持更强的渗透调节能力,表现出更好的适应性。
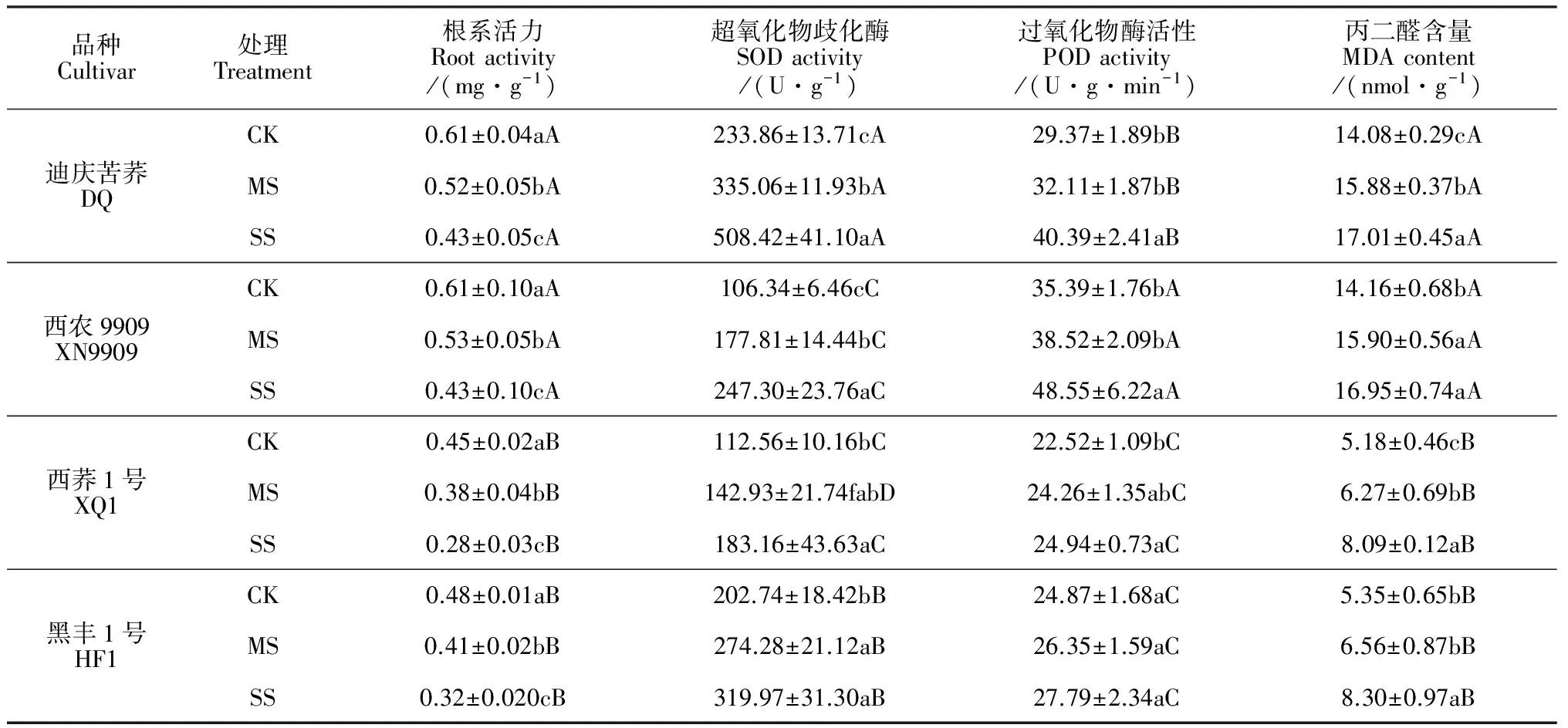
表4 干旱胁迫对不同苦荞苗期根系活力、SOD和POD活性及MDA含量的影响
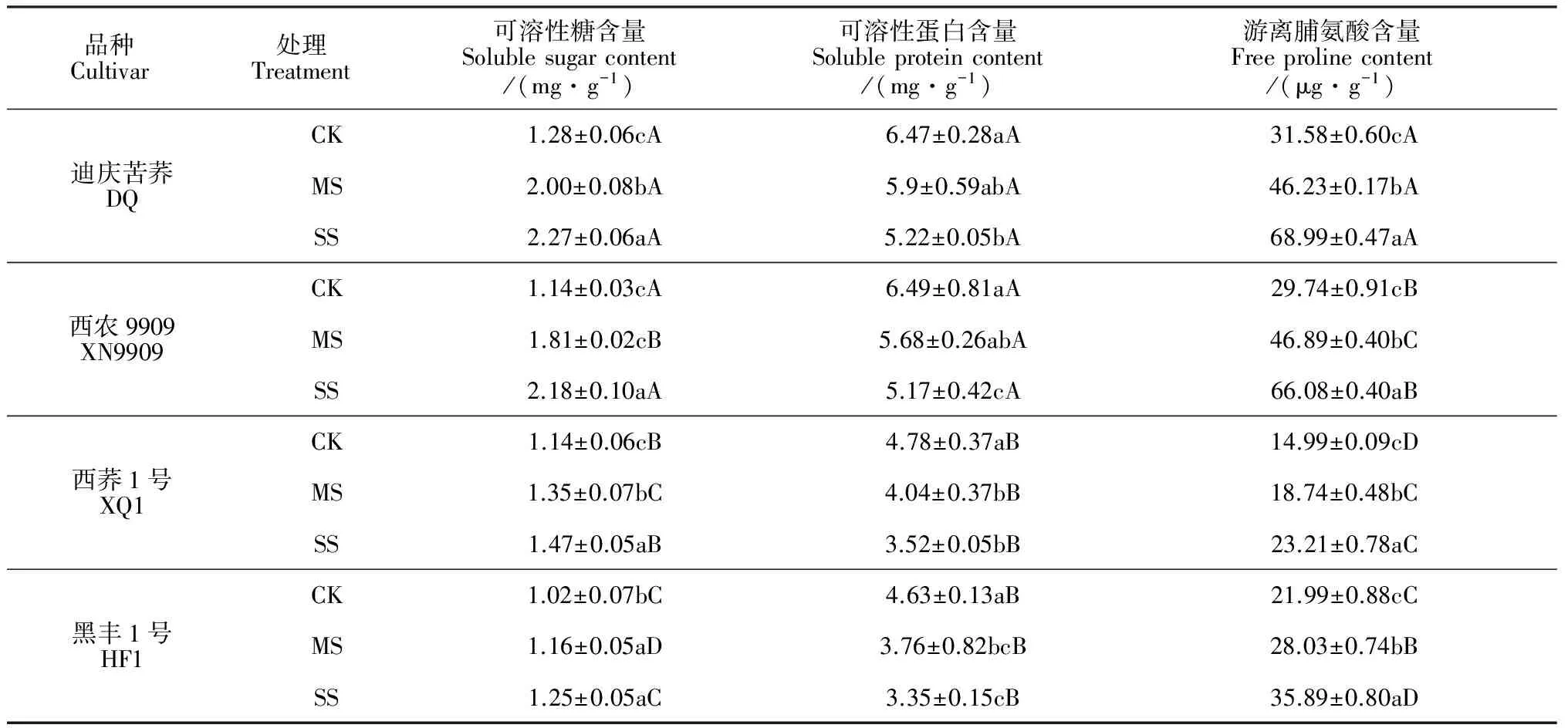
表5 干旱胁迫对不同苦荞脯氨酸、可溶性糖和可溶性蛋白含量的影响

表6 各基因型苦荞的综合指标值、权重、D值及综合评价
2.4 各苦荞品种的综合指标值、权重、D值、综合评价及其与相关性
由抗旱性综合评价公式(1)、(2)、(3)、(4)、(5)可得出表6相关指标值,其中‘迪庆苦荞’的综合评价值(D值)最高,耐旱能力最强,其它品种由强到弱依次为‘西农9909’、 ‘黑丰1号’和 ‘西荞1号’,这与上述各品种相关生长、生理生化指标分析所得结论基本一致。
另外,进一步把21个单一指标与综合评价值(D值)做相关分析(表7)表明,苦荞苗期株高、地下部干重以及根系SOD活性和蛋白质含量与D值呈显著正相关,根系脯氨酸含量和可溶性糖含量与D值呈极显著正相关。说明这6个指标对苦荞品种苗期抗旱性能力强弱的表达较为敏感,在苦荞苗期抗旱性测定工作中可有选择性地测定上述指标,使鉴定工作简化。
3 讨 论
3.1 不同抗旱性苦荞幼苗对干旱胁迫的形态响应
水是植物发生的最初环境,也是植物生长发育的必要条件。前人的研究表明,水分胁迫会影响形态指标,影响明显或降低最多的品种往往抗逆性最差[26-27]。本研究表明,干旱胁迫对苦荞幼苗地上部的影响明显大于对根系的影响,并且在一定程度上干旱胁迫可促进其根系增长,且耐旱性最好品种的根冠比在一定程度上有所升高这是苦荞形态指标对干旱胁迫的一种响应机制;不同耐旱性苦荞品种对干旱胁迫的响应存在较大差异,耐旱品种受干旱胁迫的影响较小,其地上部农艺性状较不耐旱品种表现出明显优势,对干旱环境适应性更强。
干旱胁迫下苗期根系形态直接与水分含量的多少相关,较好的根系形态是植物水分吸收的重要前提。本研究得出,耐旱型苦荞在干旱环境中主根生长得到促进,适应性较好,说明干旱环境下耐旱品种植株通过增大根系与土壤的接触面积来获取水分和养分,这与张永清等[28]对几种谷类作物根土系统的研究结果相似。此外,两个耐旱苦荞品种根系体积和根系表面积等指标受干旱环境影响程度低于不耐旱品种,由此表明耐旱品种在干旱环境中可保持较完整的根系形态来适应缺水环境。
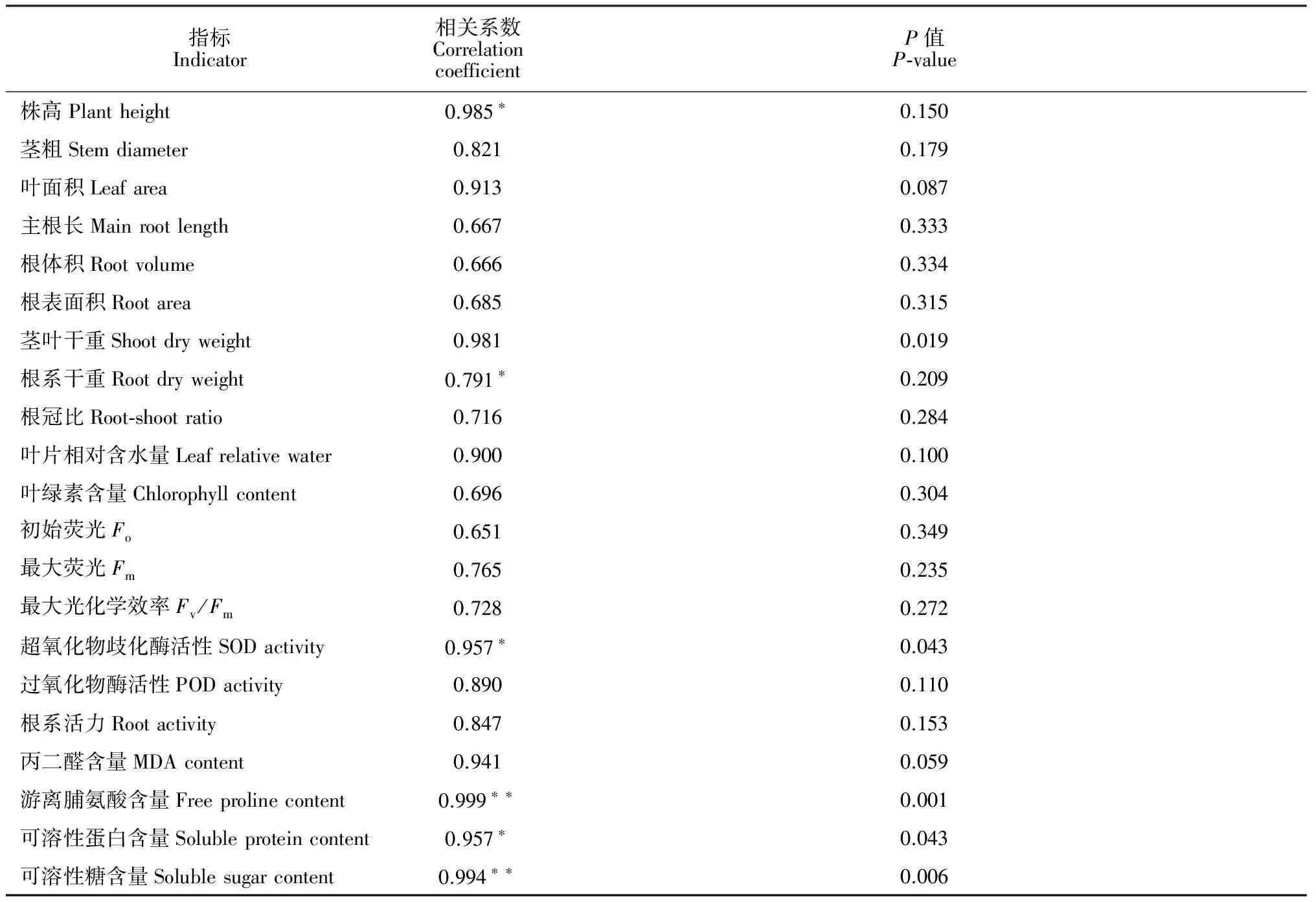
表7 苦荞幼苗单一指标与综合评价值(D值)的相关系数
注:*和**分别表示相关性达到0.05和0.01显著水平
Note:*and**indicate significant correlation at 0.05 and 0.01 level, respectively
3.2 不同抗旱性苦荞幼苗对干旱胁迫的生理响应
胁迫环境可导致植株形态学参数和生理生化指标发生改变[29]。首先,从叶片来看,本次研究发现,耐旱苦荞品种幼苗叶片叶绿素含量、相对含水量、Fm及Fv/Fm在干旱胁迫下降幅度均小于不耐旱品种,且其Fo有所升高,说明耐旱品种在干旱环境中叶绿素含量及捕获激发能的效率相对较高, PSⅡ反应中心受破坏程度较小,光合能力较强,适应性较好。其次,从根系来看,本研究表明干旱胁迫下苦荞苗期根系活力、可溶性蛋白质含量显著下降,根系SOD和POD活性以及MDA、可溶性糖及游离脯氨酸含量显著升高,且不同耐旱性品种对干旱胁迫的响应存在明显差异。在干旱胁迫环境中,耐旱品种的根系SOD、POD活性增幅较大,能够较大程度地解除或减缓毒性O-和H2O2作用对植株造成的过氧化伤害,降低细胞膜受伤程度,增强抗性;同时,其根系可溶性糖、游离脯氨酸等有机物质含量的增幅也较大,可大幅度提高根系的渗透调节能力,降低渗透势,并起到延缓植株衰老的作用。由此可知,干旱逆境下耐旱品种提高保护酶的能力较强,可快速清除有害成分,缓减膜脂过氧化作用,从而降低MDA含量,可更好地适应干旱环境,这是其耐旱胁迫的重要生理机制之一;而耐旱苦荞品种在旱胁迫下仍具有较高的根系活力及蛋白质含量也保证了其在干旱环境表现出较强的生存能力。
3.3 不同抗旱性苦荞的综合指标值及其与权重的相关性
作物的抗旱性是由多因素、多机制共同作用的复杂综合性状,不同作物的抗旱机制存在着差异[30],用单一指标很难全面、精准地反映各品种的耐旱性强弱,所以,用多个指标去综合评价作物的耐旱性才比较可靠。本试验通过隶属函数法计算出苦荞幼苗各指标的耐旱系数[μ(xi)]并将其作为评价苦荞单项耐旱能力大小的指标,并采用主成分分析法得出各苦荞品种综合抗旱能力(D值),将参试的4个品种进行耐旱性排序。μ(xi)为隶属函数法处理后的抗旱系数,将其作为抗旱性筛选指标能够消除不同基因型苦荞间固有的生物学和遗传学特性差异;D值是一个无量纲的纯数,所以各品种间的抗旱性差异具可比性。再通过对D值与各指标之间的关系相关性分析,筛选出对苦荞耐旱性影响较为显著的指标,进一步得到快速鉴别苦荞耐旱能力的指标。本研究结果表明:4个苦荞品种根据D值的排序为‘迪庆苦荞’> ‘西农9909’> ‘黑丰1号’> ‘西荞1号’,与之前单一指标分析的结果基本一致。同时通过相关性分析发现,苦荞幼苗株高、地下部干重以及根系SOD活性和可溶性蛋白含量与D值呈显著正相关,根系脯氨酸含量和可溶性糖含量与D值呈极显著正相关,说明这6个指标在苦荞对干旱逆境的感受最为敏感,可作为苦荞苗期快速、准确的抗旱性鉴定指标。此外,作物抗旱能力不仅与抗旱生理生化指标有关, 还与干旱胁迫发生的时期有密切关系。本研究只对苦荞苗期形态、生理指标进行了综合性探讨,今后需考虑其他时期各指标对苦荞干旱的响应变化,从而得出更全面可靠的结论。
综上所述,干旱胁迫对苦荞苗期各指标均有显著影响,本研究在前期预实验结果的基础上,通过干旱胁迫处理和隶属函数法与主成分分析进一步确定出4个参试品种的耐旱等级差异,其从强到弱依次为‘迪庆苦荞’、‘西农9909’、‘黑丰1号’和‘西荞1号’;同时,苗期株高、地下部干重以及根系SOD活性、可溶性蛋白含量、脯氨酸含量和可溶性糖含量这6个指标对干旱胁迫反应较敏感,可作为苦荞苗期抗旱性的快速鉴定指标。
参考文献:
[1]张雄. 黄土高原小杂粮生产可持续发展研究[D]. 陕西杨陵:西北农林科技大学2003: 20-30.
[2]万燕, 韦爽, 贾晓凤,等. 荞麦抗旱性研究进展[J].作物杂志, 2015,(2): 23-26.
WAN Y, WEI S, JIA X F,etal. Advances in drought resistance of buckwheat[J].Crops, 2015,(2):23-26.
[3]ZHOU X, HAO T, ZHOU Y L. Relationships between antioxidant compounds and antioxidant activities of tartary buckwheat during germination[J].JournalofFoodScienceandTechnology, 2015,52(4):2 458-2 463.
[4]HUANG X Y, ZELLER F J, HUANG K F. Variation of major minerals and trace elements in seeds of tartary buckwheat[J].GeneticResourcesandCropEvolution, 2014,61(3): 567-577.
[5]汤章城. 植物对水分胁迫的反应和适应性__抗逆性的一般概念和植物的抗涝性[J]. 植物生理学通讯, 1983,(3): 24-29.
TANG Z C. The reaction and adaptability of plants to water stress[J].PlantPhysiologyCommunications, 1983, (3): 24-29.
[6]李鑫, 张永清, 王大勇, 等. 水氮耦合对红小豆根系生理生态及产量的影响[J]. 中国生态农业学报, 2015,23(12): 1 511-1 519.
LI X, ZHANG Y Q, WANG D Y,etal. Effects of coupling water and nitrogen on root physio-ecological indices and yield of adzuki bean[J].ChineseJournalofEco-Agriculture, 2015,23(12): 1 511-1 519.
[7]张永清, 苗果园. 水分胁迫条件下有机肥对小麦根苗生长的影响[J]. 作物学报, 2006,32(6): 811-816.
ZHANG Y Q, MIAO G Y. Effects of manure on root and shoot growth of winter wheat under water stress[J].ActaAgronomicaSinica, 2006,32(6): 811-816.
[8]徐笑宇, 方正武, 杨璞, 等. 苦荞遗传多样性分析与核心种质筛选[J]. 干旱地区农业研究, 2015,1(33): 268-277.
XU X Y, FANG Z W, YANG P,etal. Genetic diversity analysis of tartary buckwheat and selection of core collections[J].AgriculturalResearchintheAridAreas, 2015,1(33): 268-277.
[9]汪灿, 胡丹, 杨浩, 等. 苦荞主要农艺性状与产量关系的多重分析[J]. 作物杂志, 2013,6(11): 18-22.
WANG C, HU D, YANG H,etal. Genetic diversity of 104 tartary buckwheat accessions[J].Crops, 2013,6(11): 18-22.
[10]李瑞国, 高冬丽, 柴岩, 等. 苦荞资源农艺性状因子聚类分析[J]. 干旱地区农业研究, 2007,25(6):80-84.
LI R G, GAO D L, CHAI Y,etal. Factor and cluster analysis of tartary buckwheat resources[J].AgriculturalResearchintheAridAreas, 2007,25(6): 80-84.
[11]刘琴, 张薇娜, 朱媛媛, 等.不同产地苦荞籽粒中多酚的组成、分布及抗氧化性比较[J]. 中国农业科学, 2014,47(14): 2 840-2 852.
LIU Q, ZHANG W N, ZHU Y Y,etal. Comparison of the constitutions, distribution, and antioxidant activities of polyphenols from different varieties of tartary buckwheat seed produced from different regions of China[J].ScientiaAgriculturaSinica, 2014,47(14): 2 840-2 852.
[12]QIN P Y, WANG Q, SHAN F,etal. Nutritional composition and flavonoids content of flour from different buckwheat cultivars[J].InternationalJournalofFoodScience&Technology, 2010,45(5): 951-958.
[13]葛维德, 赵阳, 刘冠求. 播种期对苦荞主要农艺性状及产量的影响[J]. 杂粮作物, 2009,29(1): 36-37.
GE W D, ZHAO Y, LIU G Q. Effects of sowing time on main agronomic characters and yield of tartary buckwheat[J].RainFedCrops, 2009,29(1): 36-37.
[14]万丽英. 播种密度对高海拔地区苦荞产量与品质的影响[J]. 作物研究, 2008,22(1):42-44.
WAN L Y. Effect of planting density on yield and quality of tartary buckwheat in high altitude region[J].CropResearch, 2008,22(1): 42-44.
[15]田秀英, 李会合, 王正银. 施硒对苦荞N, P, K营养元素和土壤有效养分含量的影响[J]. 水土保持学报, 2009,23(3): 112-115.
TIAN X Y, LI H H, WANG Z Y. Effect of Se application on contents of nitrogen, phosphorus and potassium in tartary buckwheat and content of available nutrients in soil[J].JournalofSoilandWaterConservation, 2009,23(3):112-115.
[16]宋毓雪, 郭肖, 等. 不同氮磷钾肥料处理对苦荞籽粒充实度及产量的影响[J]. 浙江农业学报, 2014,26(6): 1 568-1 572.
SONG Y X, GUO X,etal. Effects of different N,P,K treatments on the yield and plumpness of tartary buckwheat[J].ActaAgriculturaeZhejiangensis, 2014,26(6): 1 568-1 572.
[17]张强, 李艳琴. 基于矿质元素的苦荞产地判别研究[J]. 中国农业科学, 2011,44(22):4 653-4 659.
ZHANG Q, LI Y Q. The origin discrimination of tartary buckwheat based on the mineral elements[J].ScientiaAgriculturaSinica, 2011,44(22): 4 653-4 659.
[18]LIM J H, PARK K J,etal. Effect of salinity stress on phenolic compounds and carotenoids in buckwheat (FagopyrumesculentumM.) sprout[J].FoodChemistry, 2012,135(3): 1 065.
[19]胡丽雪. 苦荞幼苗对铅铝胁迫的生理生化反应及其矫治[D]. 贵阳:贵州师范大学, 2014.
[20]董新纯, 赵世杰, 郭珊珊, 等. 增强UV-B条件下类黄酮与苦荞逆境伤害和抗氧化酶的关系[J]. 山东农业大学学报(自然科学版), 2006,37(2):157-162.
DONG X C, ZHAO S J, GUO S S,etal. Role of flavonoids on stress in jury and antioxyd ative enzymes inFagopyrumtataricumseedlings under enhanced UV-B radiation[J].JournalofShandongAgriculturalUniversity(NaturalScience), 2006,37(2): 157-162.
[21]陈鹏, 张德玖, 李玉红, 等. 水分胁迫对苦荞幼苗生理生化特性的影响[J]. 西北农业学报, 2008,17(5): 204-207.
CHEN P, ZHANG D J, LI Y H,etal. Effect of drought stress during seedling stage on physiological and biochemical traits of tartary buckwheat[J].ActaAgriculturaeBoreali-OccidentalisSinica, 2008,17(5): 204-207.
[22]YAO Y, XUAN Z, LI Y,etal. Effects of ultraviolet-b radiation on crop growth, development, yield and leaf pigment concentration of tartary buckwheat (Fagopyrumtataricum) under field conditions[J].EuropeanJournalofAgronomy, 2006,25(3): 215-222.
[23]张志良, 瞿伟菁, 李小方.植物生理学实验指导(第4版)[M]. 北京: 高等教育出版社, 2009: 32-227.
[24]许自成, 黎妍妍, 毕庆文, 等. 湖北烟区烤烟气候适生性评价及与国外烟区的相似性分析[J]. 生态学报, 2008,28(8): 3 832-3 838.
XU Z C, LI Y Y, BI Q W,etal. Evaluation on climatic feasibility of tobacco-growing areas in Hubei and its similarity to that of foreign tobacco-growing areas[J].ActaEcolSin, 2008,28(8): 3 832-3 838.
[25]闫江艳, 张永清, 冯晓敏, 等. 干旱胁迫及复水对不同黍稷品种根系生理特性的影响[J]. 西北植物学报, 2012,32(2): 348-354.
YAN J Y, ZHANG Y Q, FENG X M,etal. Effect of drought stress and rewatering on physiological characteristics of roots in different proso millet varieties[J].ActaBotanicaBoreali-OccidentaliaSinica, 2012,32(2): 348-354.
[26]郑盛华, 严昌荣. 水分胁迫对玉米苗期生理和形态特性的影响[J].生态学报, 2006,26(4):1 138-1 143.
ZHENG S H, YAN C R. The ecophysiological and morphological characteristics of maize in seeding stage under water stress[J].ActaEcologicaSinica, 2006,26(4): 1 138-1 143.
[27]王贺正, 李艳, 马均, 等. 水稻苗期抗旱性指标的筛选[J]. 作物学报, 2007,33(9): 1 523-1 529.
WANG H Z, LI Y, MA J,etal. Screening indexes of drought resistance during seedling stage in rice[J].ActaAgronomicaSinica, 2007,33(9): 1 523-1 529.
[28]张永清. 几种谷类作物根土系统的研究[D]. 山西太谷: 山西农业大学, 2005.
[29]张美俊, 乔治军, 等. 不同糜子品种对低氮胁迫的生物学响应[J]. 植物营养与肥料学报, 2014,20(3): 661-669.
ZHANG M J, QIAO Z J,etal. Biological response of different cultivars of millet to low nitrogen stress[J].JournalofPlantNutritionandFertilizer, 2014,20(3): 661-669.
[30]邵惠芳, 陈征, 等. 两种烟草幼苗叶片对不同强度干旱胁迫的生理响应比较[J].植物生理学报, 2016,52(12): 1 861-1 871.
SHAO H F, CHEN Z,etal. Physiological responses of two tobacco cultivar leaves to different drought stresses during seedling stage[J].PlantPhysiolJ, 2016,52(12): 1 861-1 871.
