The relaxin peptide family – potential future hope for neuroprotective therapy? A short review
2018-04-04MariusNistorMartinSchmidtRenSchiffner
Marius Nistor, Martin Schmidt, René Schiffner
1 Department of Neurology, Jena University Hospital, Friedrich Schiller University, Jena, Germany
2 Orthopaedic Department, Jena University Hospital, Friedrich Schiller University, Jena, Germany
3 Institute for Biochemistry II, Jena University Hospital, Friedrich Schiller University, Jena, Germany
Introduction
After its discovery in the 1920s, the peptide hormone relaxin was assumed to be a primarily reproductive hormone.Present in all mammals, relaxin mediates the necessary cardiovascular adaptions during pregnancy, such as increased renal blood flow and systemic vasodilation (Wilkinson et al., 2005; Callander and Bathgate, 2010). During the decades since relaxin’s discovery, though, the relaxin family peptides have proven to be more complex than initially suspected.With the identification of the relaxin-3 peptide in 2002,humans are now known to possess seven peptides belonging to the relaxin family (relaxin-1, relaxin-2, relaxin-3,insulin-like peptide (INSL) 3, INSL4, INSL5 and INSL6)(Rosengren et al., 2006; Callander and Bathgate, 2010). Most of the circulating relaxin in the human body, relaxin-2, is expressed by the RLN2 gene. Currently, vasodilatory, angiogenic, anti-apoptopic, anti-fibriotic and anti-inflammatory effects can be linked to relaxin, both in males and females(Sarwar et al., 2017). Relaxin acts through a number of signal transduction pathways, for example via generation of cyclic adenosine monophosphate (cAMP), nitric oxide (NO),cyclic guanosine monophosphate (cGMP), or activation of mitogen-associated protein kinases (MAPKs). Its vasodilatory effects can especially be traced back to interaction with the nitric oxide system, and all of the three subtypes(endothelial, inducible and neuronal) of NO synthases(NOS) (Sarwar et al., 2017). The varied effects of relaxin-2 suggest differing relaxin receptor densities which might be the cause for region-speci fic control of various signal transduction pathways. RXFP1 and RXFP2 represent the known relaxin-2-activated receptors (Bathgate et al., 2013). These receptors are involved in various aspects of the remodeling of cerebral parenchymal arterioles (Chan and Cipolla, 2011;Chan et al., 2013), as well as in the reduction of vascular resistance, and in an increased blood flow in renal and systemic small arteries in humans and rats (Debrah et al., 2006;Conrad and rars Shroff, 2011); these effects, however, often require longer timescales. Area-speci fic distribution of RXFP1-mRNA expression and relaxin binding sites of the rat brain are well documented (Ma et al., 2012).While relaxin-2 has roughly nanomolar affinities for both receptors (Halls et al., 2015), the affinity is about an order of magnitude higher for the RXFP1 receptor.
While the last years have seen major advances in uncovering the relaxin family’s effects and their underlying mechanisms, many basic concepts and especially potential therapeutic bene fits remain uncertain (Callander and Bathgate,2010; Sarwar et al., 2017).
In recent years, relaxin-2’s therapeutic potential has been mainly investigated in the context of cardiovascular diseases. Animal models initially presented promising results of improved healing and protection against necrotic cell death of cardiomyocytes in ischemia-reperfusion injury, as well as partly reduced infarction size in myocardial ischemia models (Sarwar et al., 2017). Relaxin-2’s physiologic properties, which include reduced proliferation of fibroblasts and pro-inflammatory cytokines as well as vasodilation,therefore seem to recommend the peptide hormone as a therapeutic drug in cardiac illnesses (Ghosh et al., 2017).The RELAX-AHF (international multicenter phase III)study, which enrolled 1,161 patients with acute heart failure(AHF), initially reported positive results of reduced 180-day mortality and a reduction of dyspnea within the first 5 days of treatment (Ghosh et al., 2017). The succeeding RELAX-AHF-2 study, a global multicenter phase III study with 6,600 patients, was supposed to provide further details on the therapeutic potential of serelaxin, but ultimately failed to meet its endpoints of reducing cardiovascular mortality and preventing the worsening of heart failure (No authors listed, 2017).
While relaxin-2’s therapeutic effects have primarily been investigated in a cardiac context, the very same properties suggest a potential neuroprotective effect of relaxin-2 as well.
Current Research on Relaxin in a Neuropathophysiological Context
Few studies so far have specifically examined relaxin hormones as potential neuroprotective agents. Nonetheless, the few studies that did reported mainly favorable results and the substance’s general properties furthermore appear to warrant more in-depth research into its potential neuroprotective properties.
Wilson et al. (2005) investigated the potential neuroprotective effects of relaxin-2 in a rat stroke model. Thirty minutes before a middle cerebral artery occlusion (MCAO), rats received an intracortical injection of relaxin-2. A 2,3,5-triphenoltetrazolium chloride (TTC) stain subsequent to brain removal four hours post-intervention revealed that relaxin-2 pre-treated animals exhibited a signi ficantly reduced infarct size as compared to saline-injected control animals (Wilson et al., 2005). At this point, Wilson et al. already hypothesized that the neuroprotective effect that could be observed in relaxin-2-treated rats might be due to NO-mediated vasodilation and subsequently improved collateral perfusion through nearby vasculature. An alternative explanation considered estrogen receptor activation through relaxin-2 as the cause of the observed neuroprotection, since relaxin-2 was known to activate estrogen receptors in the uterus and preceding studies had demonstrated that estrogen pre-treatment similarly reduced ischemia-related infarct sizes (Wilson et al., 2005).
Following up on these early results and investigating their initial assumptions, Wilson et al. published another study on relaxin-2 in the following year (Wilson et al., 2006). While basically the same study design (MCAO, brain removal after 4 hours and TTC-staining) was employed, a number of new experimental groups were added, namely combined relaxin-2 and estrogen injection, relaxin-2 and estrogen receptor antagonist injection (ICI 182,780), as well as intravenous injection of relaxin-2 as well as relaxin-2 in combination with an endothelial NOS (eNOS) inhibitor (l-NIO) (Wilson et al.,2006). These additional experimental groups revealed that relaxin-2’s neuroprotective effects are eNOS related and not dependent on estrogen receptor activation. While combined relaxin-2 and estrogen injection revealed an additive (insigni ficant) decrease of infarct size, the injection of an estrogen receptor antagonist did not abolish relaxin-2’s effects — unlike eNOS inhibition, which signi ficantly reduced the neuroprotective effect of relaxin-2 (Wilson et al., 2006). Exclusive relaxin injection again demonstrated a reduction of infarct size in the rat stroke model, both through intracortical and intravenous administration (Wilson et al., 2006).
A more recent study by Bergeron et al. (2015) again confirmed relaxin-2’s neuroprotective effect and furthermore explored in a more in-depth way the underlying mechanisms. While also utilizing a rat stroke model, Bergeron et al. (2015) employed both relaxin-2 and relaxin-3 receptor expression through real-time PCR and assessed the animals’ brains for infarct size. Furthermore, the study employed a wide range of experimental groups that varied in the manner of the MCAO intervention (permanent or transient), the time point of relaxin administration (before or after intervention) and the additional injection of receptor antagonists. Both relaxin-2 and relaxin-3 provided neuroprotection during permanent MCAO, regardless of the time of administration. In con firmation with Wilson et al.’s earlier study, eNOS inhibition abolished these effects.Relaxin receptor (Rxfp) densities varied: Rxfp1 expression was signi ficantly higher than Rxfp3 expression in the cortex,while astrocytes revealed higher Rxfp3 expression (Ma et al.,2006; Bergeron et al., 2015). Interestingly, relaxin-2 could not reduce infarct volume during transient MCAO, while relaxin-3 was again successful in reducing infarct size as compared to saline-treated control animals (Bergeron et al.,2015). As an explanation, Bergeron et al. (2015) refer to the promiscuous binding of relaxin-3 to both Rxfp1 and Rxfp3,while relaxin-2 only activates Rxfp1 (and Rxfp2, albeit this was not subject of this study). The authors therefore propose, that Rxfp1’s vasodilatory effects (triggered by both relaxin-2 and relaxin-3) are responsible for the neuroprotective effects during permanent MCAO, while the neuroprotective effects of relaxin-3 during the transient MCAO(basically an ischemia-reperfusion injury) is based on protection from apoptotic mechanisms mediated by Rxfp3 on a cellular level (Bergeron et al., 2015).
Finally, relaxin-2 has furthermore already been once (to our knowledge) tested on human subjects who had suffered a stroke. Milia et al. (2013) describe a trial performed in a rehabilitation unit in Italy in 2013, consisting of 18 patients who received relaxin-2 and rehabilitation as compared to 18 patients who were treated with rehabilitation alone. All participants had suffered from an ischemic stroke in the previous three weeks. The Modified Rankin Scale (mRs),the Trail Making Test (TMT) and Functional Independence Measure (FIM) were employed to compare patients and to assess potential progress. The TMT and mRs exhibited better scores for relaxin-treated patients at both temporal evaluation points (after 20, respectively 40 days), while the FIM revealed no signi ficant differences at day 20 but a clear bene fit on day 40 (FIM 96 as compared to non-relaxin-treated patient group with 75). The authors note that no obvious adverse effects were noticed during relaxin treatment (Milia et al., 2013). As stated in the paper, relaxin-2 was administered in a dose of 40 μg per day (per patient), supposedly orally.Compared to other studies, e.g., RELAX-AHF (Ghosh et al.,2017), this dose seems to be extremely low (roughly 50–100 times lower). Furthermore, data on plasma concentrations obtained by this dosage were not presented, which raises the question as to whether the used amount of relaxin-2 couldactivate the relaxin-receptors at all (Table 1).
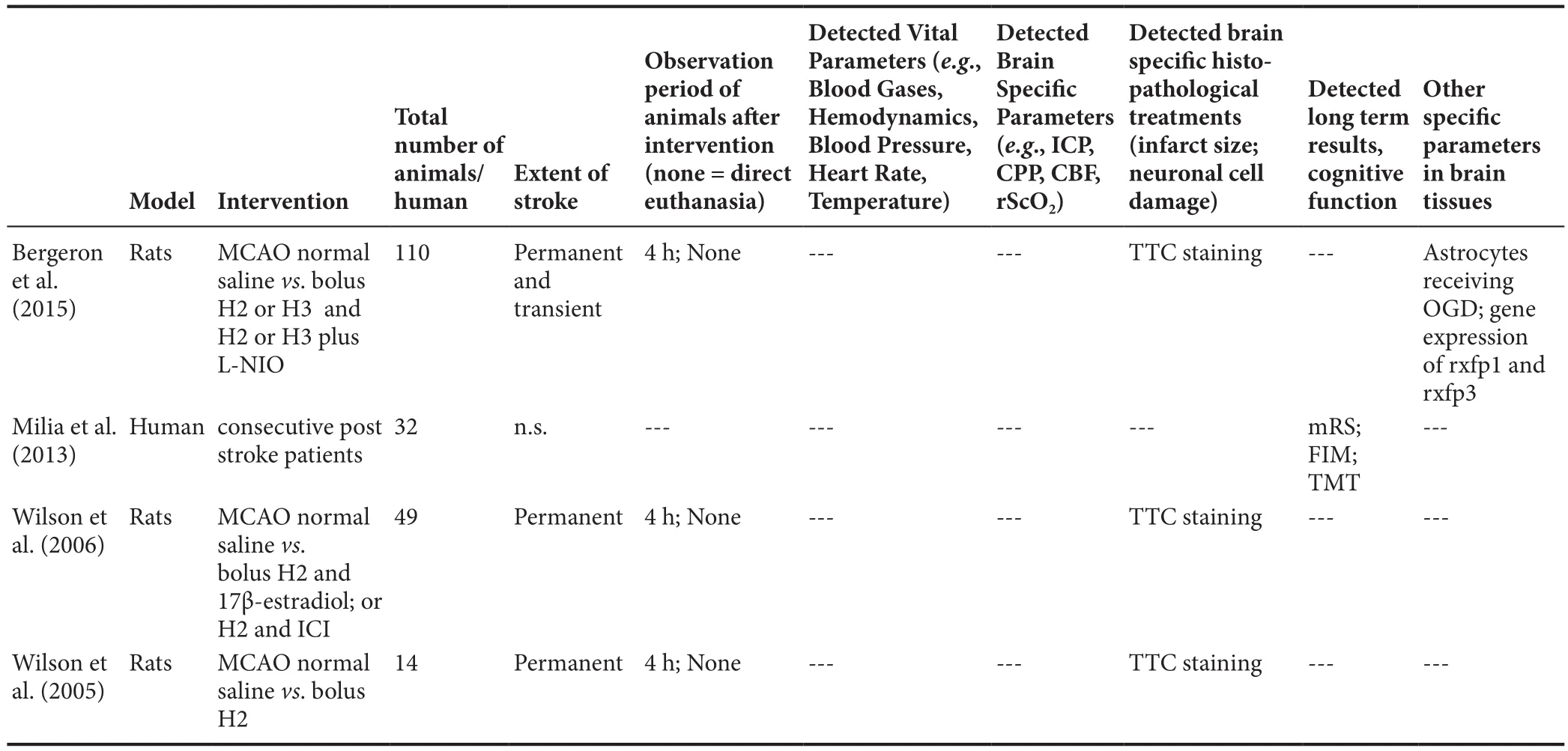
Table 1 General overview of included studies
Future Neuroprotective Application of Relaxin?
While the results of the studies discussed above justify an optimistic outlook on the potential of relaxin peptide hormones as neuroprotective agents, it has to be clearly stated that our current knowledge on the exact mechanisms through which they exert their effects is incomplete, and that further in-depth research is required. While all three of the rat experimental models reported a reduced infarct size in relaxin-treated rats, their results were only based on staining procedures performed after brain removal. To elucidate both the safety and the effectiveness of relaxin treatment,more studies are needed. A wider array of brain speci fic parameters and longer observation periods of the experimental animals should be included into future more comprehensive efforts.Noteworthy is that none of the studies documented vital parameters (Wilson et al., 2005, 2006; Bergeron et al.,2015) — this even includes Milia et al.’s study on humans,that, while stating that no adverse effects were observed,similarly did not report basic vital parameters (Milia et al.,2013). An apparent difference regarding the baseline values of the TMT and mRS scores of relaxin-treated patients and the control group is likely due to the randomization procedure of the small patient cohort that was included (Milia et al., 2013).
Bischoff et al. (2016) have performed a study on the microcirculatory effects of serelaxin (human recombinant relaxin-2) in a sheep model. After injection of 30μg/kg serelaxin, an increased cortical cerebral blood flow (CBF) was observed through Laser Doppler flowmetry and sidestream dark- field imaging, but not a concomitant increase in subcortical CBF. Expression levels of Rxfp1 and Rxfp2 were additionally investigated and interestingly did not differ between cortex and subcortex, which might indicate that the greater cortical CBF response is due to area-speci fic differences in signal transduction pathways (Bischoff et al., 2016).This study’s results therefore correspond to the vasodilation and improved circulation described by Wilson et al. (2005,2006) and Bergeron et al. (2015). These effects could conceivably be utilized in situations of cerebral hypoperfusion.Severe hemorrhagic shock is known to transcend the limits of the cerebral autoregulation and to lead to concomitant cerebral damages through hypoperfusion (Rickards, 2015).Cortical regions are more prone to damages during these situations than the subcortex (Heckbert et al., 1998), the reasons for these disparate occurrences of cerebral damages are currently not known. A systematic review by Nistor et al. demonstrated that, while some promising approaches are currently under research, to date no speci fic neuroprotective strategies are available that would offer specific protection from cerebral damages due to hemorrhagic shock (Nistor et al., 2017). The context of cerebral damages due to states of hemorrhagic shock might therefore represent a further field of the potential application of relaxin-2 as a neuroprotective agent — especially since the hormone’s NO-mediated vasodilation might mitigate cellular damages. The relatively new discovered relaxin-3 might represent an interesting new field of research as well, both in the context of an elementary understanding of neurophysiology and its potential thera-peutic application. Callander and Bathgate note that already,there is evidence that relaxin-3 is at least a contributing factor in many behavioral functions, memory, sleep and the stress system (Callander and Bathgate, 2010). Bergeron’s investigations in mice con firm that relaxin-3 has a therapeutic value as well — since Rxfp3 seems to mitigate apoptotic cell death (Bergeron et al., 2015), relaxin-3 could also be applied for neuroprotection during hemorrhagic shock, since the mechanism of this type of hypoperfusion is essentially an ischemia-reperfusion injury as well.
Conclusion
In conclusion, the relaxin peptide hormone family seems to provide many opportunities for future research and might be utilized for therapeutic purposes. Early studies have yielded positive results that strengthen the assumption that relaxin-2 possess neuroprotective properties. Relaxin-2 might be used for post-stroke treatment; its natural occurrence in humans and the current state of research indicate that severe adverse effects cannot be expected. Relaxin-3 might mitigate the damage of ischemia-reperfusion injuries of varying causes through currently not well researched mechanisms.Therefore, the promising results of the available studies warrant future well-designed studies on neuroprotection by relaxin peptides.
Author contributions:MN and RS designed and performed the study,analysed data. MN, MS and RS discussed data, interpreted results, draft-ed, edited and finally approved the manuscript.
Con flicts of interest:None declared.
Plagiarism check:Checked twice by iThenticate.
Peer review:Externally peer reviewed.
Open access statement:This is an open access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 3.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as the author is credited and the new creations are licensed under identical terms.
Open peer review report:
Reviewer:Andrew L. Gundlach, The University of Melbourne, Australia.
Comments to authors:A ‘strength’ of the article is the enticing concept that relaxin systems might be neuroprotective.
Bathgate RA, Halls ML, van der Westhuizen ET, Callander GE, Kocan M, Summers RJ (2013) Relaxin family peptides and their receptors.Physiol Rev 93:405-480.
Bergeron LH, Willcox JM, Alibhai FJ, Connell BJ, Saleh TM, Wilson BC, Summerlee AJ (2015) Relaxin peptide hormones are protective during the early stages of ischemic stroke in male rats. Endocrinology 156:638-646.
Bischoff SJ, Schmidt M, Lehmann T, Irintchev A, Schubert H, Jung C, Schwab M, Huber O, Matziolis G, Schiffner R (2016) Increase of cortical cerebral blood flow and further cerebral microcirculatory effects of Serelaxin in a sheep model. Am J Physiol Heart Circ Physiol 311:H613-620.
Callander GE, Bathgate RA (2010) Relaxin family peptide systems and the central nervous system. Cell Mol Life Sci 67:2327-2341.
Chan SL, Cipolla MJ (2011) Relaxin causes selective outward remodeling of brain parenchymal arterioles via activation of peroxisome proliferator-activated receptor-γ. FASEB J 25:3229-3239.
Chan SL, Sweet JG, Cipolla MJ (2013) Treatment for cerebral small vessel disease: effect of relaxin on the function and structure of cerebral parenchymal arterioles during hypertension. FASEB J 27:3917-3927.
Conrad K1, Shroff SG (2011) Effects of relaxin on arterial dilation, remodeling, and mechanical properties. Curr Hypertens Rep 13:409-420.
Debrah DO, Novak J, Matthews JE, Ramirez RJ, Shroff SG, Conrad KP(2006) Relaxin is essential for systemic vasodilation and increased global arterial compliance during early pregnancy in conscious rats.Endocrinology 147:5126-5131.
Ghosh RK, Banerjee K, Tummala R, Ball S, Ravakhah K, Gupta A (2017)Serelaxin in acute heart failure: Most recent update on clinical and preclinical evidence. Cardiovasc Ther 35:55-63.
Halls ML, Bathgate RA, Sutton SW, Dschietzig TB, Summers RJ (2015)International Union of Basic and Clinical Pharmacology. XCV.Recent advances in the understanding of the pharmacology and biological roles of relaxin family peptide receptors 1-4, the receptors for relaxin family peptides. Pharmacol Rev 67:389-440.
Heckbert SR, Vedder NB, Hoffman W, Winn RK, Hudson LD, Jurkovich GJ, Copass MK, Harlan JM, Rice CL, Maier RV (1998) Outcome after hemorrhagic shock in trauma patients. J Trauma 45:545-549.
Ma L, Gul R, Habibi J, Yang M, Pulakat L, Whaley-Connell A, Ferrario CM, Sowers JR (2012) Nebivolol improves diastolic dysfunction and myocardial remodeling through reductions in oxidative stress in the transgenic (mRen2) rat. Am J Physiol Heart Circ Physiol 302:H2341-2351.
Ma S1, Shen PJ, Burazin TC, Tregear GW, Gundlach AL (2006)Comparative localization of leucine-rich repeat-containing G-protein-coupled receptor-7 (RXFP1) mRNA and [33P]-relaxin binding sites in rat brain: restricted somatic co-expression a clue to relaxin action? Neuroscience 141:329-344.
Milia P, Caserio M, Bani D, Rastelli TF, Sonaglia F, Bigazzi B, Bigazzi M (2013) Efficacy of relaxin on functional recovery of post stroke patients. Ital J Anat Embryol 118:S92-97.
Nistor M, Behringer W, Schmidt M, Schiffner R (2017) A systematic review of neuroprotective strategies during hypovolemia and hemorrhagic shock. Int J Mol Sci 18:E2247.
No authors listed (2017) Top heart-failure contender serelaxin flops.Nat Biotechnol 35:297.
Rickards CA (2015) Cerebral blood-flow regulation during hemorrhage. Compr Physiol 5:1585-1621.
Rosengren KJ, Lin F, Bathgate RA, Tregear GW, Daly NL, Wade JD,Craik DJ (2006) Solution structure and novel insights into the determinants of the receptor speci ficity of human relaxin-3. J Biol Chem 281:5845-5851.
Sarwar M, Du XJ, Dschietzig TB, Summers RJ (2017) The actions of relaxin on the human cardiovascular system. Br J Pharmacol 174:933-949.
Wilkinson TN, Speed TP, Tregear GW, Bathgate RA (2005) Evolution of the relaxin-like peptide family. BMC Evol Biol 5:14.
Wilson BC, Milne P, Saleh TM (2005) Relaxin pretreatment decreases infarct size in male rats after middle cerebral artery occlusion. Ann N Y Acad Sci 1041:223-228.
Wilson BC, Connell B, Saleh TM (2006) Relaxin-induced reduction of infarct size in male rats receiving MCAO is dependent on nitric oxide synthesis and not estrogenic mechanisms. Neurosci Lett 393:160-164.
杂志排行

中国神经再生研究(英文版)的其它文章
- The biological clock: future of neurological disorders therapy
- Cerebral ischemia and neuroregeneration
- SNARE complex in axonal guidance and neuroregeneration
- Heterozygous carriers of galactocerebrosidase mutations that cause Krabbe disease have impaired microglial function and defective repair of myelin damage
- Roles of neural regeneration in memory pharmacology
- Towards frequency adaptation for delayed feedback deep brain stimulations