水淹胁迫对桢楠幼树生长及光合生理特性的影响
2018-03-29陈玉凤胡红玲胡庭兴喻秀艳杨珊珊张成浩
陈玉凤, 谭 飞,2, 胡红玲①, 胡庭兴, 喻秀艳, 周 鑫, 杨珊珊, 张成浩, 张 建
(1.四川农业大学林学院, 四川 成都 611130; 2.四川省凉山州林业调查规划设计院, 四川 西昌 615000)
桢楠(Phoebezhennan)是樟科(Lauracea)楠属(Phoebe)常绿大乔木,为我国特有的二级保护珍稀渐危种,耐阴,常生长于温暖湿润气候的山麓腐殖质酸性至中性土壤中[1],主要分布在四川盆地、贵州北部、江苏、安徽、湖北、浙江等地[2-3]。桢楠不仅是珍贵的用材树种,也是优良的观赏树种,由于其重要的经济价值和广泛的用途,四川盆地西部低山丘陵区(华西雨屏区)在近年的生态恢复及园林绿化建设中发展了较多的桢楠人工林,但是该地区夏季暴雨及连绵的秋雨常造成林地淹水或积水,对桢楠幼树生长造成严重威胁,在盆地西部低洼地带栽植的桢楠幼树每年受周期性渍水的影响,生长严重不良,甚至出现了枯死现象。
水淹是一种常见的涝害胁迫,持续降水、融雪或者潮汐等都会形成水淹环境,而过多的水分会对植物生长产生不利影响[4]。水淹胁迫使土壤长时间处于水分饱和状态,导致土壤缺氧,使得植物体能量代谢及离子吸收受到抑制[5-6],进而影响植物的形态特征[7-8]和生理代谢[9]。光合作用是植物最基本的生命活动,也是碳循环及生态水文循环的关键环节[10]。研究水淹胁迫对植物光合特性的影响,有利于揭示植物光合特性对水淹胁迫响应的方式和程度。不同的水淹胁迫程度对同一植物或不同植物的影响有所不同:秋华柳(Salixvariegatafranch)在短期水淹胁迫下净光合速率、蒸腾作用及气体交换小幅度下降,但是在长期水淹状态下,其下降到一定限度后趋于平稳[11];灰木莲(Magnoliaceaeglanca)、蒲桃(Syzygiumjambos)的叶片净光合速率及气体交换随水淹时间延长不断下降,胞间CO2浓度逐渐增加[12];枫杨(Pterocaryastenoptera)和夹竹桃(Neriumindicum)的光补偿点、表观量子效率及暗呼吸速率在水淹胁迫下均无显著变化[13]。由于桢楠生长缓慢,相关研究滞后于产业发展,现阶段对桢楠的研究主要集中在对幼树的施肥[14-15]和干旱胁迫[16]等方面,而对涝害引起的胁迫则鲜见报道。因此,该研究通过模拟不同持续时间水淹胁迫,观测桢楠幼树形态和光合生理特性的响应,探讨桢楠幼树对涝害的耐受性,为桢楠幼树的培育及人工林的科学水分管理提供理论依据。
1 材料与方法
1.1 试验材料
2015年3月将1 a生桢楠幼苗栽植于盆中,进行为期1.5 a的适应性生长,期间进行常规水分管理,采用HH2土壤水分速测仪(ML2x,GBR)监测土壤含水量,使土壤水分保持在田间持水量的70%左右。栽培基质用农耕土(沙壤),其主要化学性质为:pH值 7.85,有机质24.38 g·kg-1,全氮0.66 g·kg-1,有效磷128.63 mg·kg-1,全钾5.81 g·kg-1,速效钾15.27 mg·kg-1。栽植容器为聚乙烯塑料盆(口径29 cm、底径17 cm、盆高25 cm),每盆装土15 kg。
1.2 试验设计
于2016年7月中旬至8月底,即四川盆地月降雨量最高的季节,在四川农业大学科研园区(成都温江)进行试验,水淹处理采用套盆法,即将栽有桢楠幼树的小盆(每盆1株)放入大盆,然后向大盆内灌水,至水面没过小盆约2 cm,每7 d处理1批,每批放入5盆,定期补水,直到最早1批处理植株萎蔫失绿,在相同的时间内,共形成7×0、7×1、7×2、7×3、7×4、7×5 d这6个不同水淹胁迫处理时段(即不同程度涝害)梯度,相应记作D0(CK,未进行水淹处理)、D7、D14、D21、D28、D35。胁迫结束后采样测定各指标值。
第1批处理植株在水淹28 d时出现明显的萎蔫,但叶片还未完全失绿,并可检测出微弱的气体交换参数,为了能够准确找到桢楠幼树水淹致死时间,持续进行胁迫,到第35 d时,首批胁迫的植株全部出现严重枯萎和失绿状态,且不能检测出气体交换参数,表明这批植株已经处于死亡状态,此时撤除水淹。因此该试验能纳入分析的数据只涉及到5个处理,即D0、D7、D14、D21、D28。
1.3 测定指标及方法
1.3.1形态指标测定
植株的株高与地径分别用卷尺与游标卡尺测量。
1.3.2根系活力测定
根系活力采用TTC比色法[17]测定。
1.3.3叶绿体色素测定
叶绿素含量采用丙酮浸提法[18]测定。
1.3.4气体交换参数的测定
采用Li-6400(Li-Cor Inc.,USA)便携式光合仪,设定光照强度为1 200 μmol·m-2·s-1,CO2浓度为400 μmol·mol-1,温度为25 ℃,相对湿度为45%~65%,在各处理桢楠幼树上自顶端向下选取大致相同部位的成熟功能叶片,测定其净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr),每个处理选取5片进行测定,每片记录10个数据,测定时间在第1天09:00—11:00完成。
1.3.5光合日变化测定
采用Li-6400光合仪,设定CO2浓度为400 μmol·mol-1,测定不同时段的光强、叶片温度以及叶片Pn日变化。测定时间为07:00—18:00,第1、2次测定间隔1 h,以后每间隔2 h测1次,共测定7次,每个处理选取5片叶子进行测定,每片记录10个数据,测定在试验结束后第1天完成。
1.3.6光合-光响应曲线和光合-CO2响应曲线的测定
采用Li-6400光合仪,选取大致相同部位的成熟功能叶片测定各处理植株光合-光响应曲线和光合-CO2响应曲线,每个处理设3个重复,测定时叶室温度为25 ℃。光合-光响应曲线的测定条件:CO2浓度设定为400 μmol·mol-1,运用自动测量程序按梯度依次由低到高设定光照强度为0、25、50、75、100、200、400、600、800、1 000、1 200、1 600、1 800 μmol·m-2·s-1。光合-CO2曲线的响应测定条件:光照强度设为1 200 μmol·m-2·s-1,CO2浓度梯度设定为0、50、100、150、200、300、400、600、800、1 200、1 600、2 000 μmol·mol-1。2种曲线测定分别在试验结束后第2和第3天完成。
1.4 数据处理
气孔限制值(Ls)计算公式[19]为
Ls=1-Ci/Ca。
(1)
式(1)中,Ls为气孔限制值;Ci为胞间CO2浓度,μmol·mol-1;Ca为大气CO2浓度,μmol·mol-1。
采用非直角双曲线模型拟合光合-光响应曲线[20],并计算最大净光合速率(Pmax)、近光饱和点(Lsp)和光补偿点(Lcp);采用直角双曲线模型拟合光合-CO2响应曲线[21],并计算最大净光合速率(Pmax)、近CO2饱和点(Csp)和CO2补偿点(Ccp)。
采用SPSS 20.0统计分析软件对实验数据进行单因素方差分析(One-way ANOVA),并用最小显著差数法(LSD)进行多重比较,采用Sigmaplot 12.5软件进行图表制作。
2 结果与分析
2.1 水淹胁迫对桢楠幼树生长的影响
由表1可知,桢楠幼树株高净生长量随着水淹胁迫时间的延长而降低,除D7外各处理与CK差异均达显著水平(P<0.05),且D7、D14、D21、D28分别比CK减少43.4%、59.2%、59.2%和80.8%。地径净生长量随着胁迫时间的延长呈先升后降的趋势,除D7外,各处理间差异显著(P<0.05),胁迫21 d时地径净生长量达最大,为CK的3.68倍;而胁迫28 d时地径净生长量大幅度减小,但仍显著高于CK(P<0.05),出现这种现象的原因可能是,水淹开始后一段时间内,地径因长时间浸于水中致使皮孔增大,而随着水淹时间的增加(21 d之后),涝害胁迫加剧,使其生长受阻,植株茎秆萎缩。
表1水淹胁迫下桢楠幼树株高、地径净生长量变化
Table1VariationofnetgrowthofplantheightandbasaldiameterofPhoebezhennansaplingswiththestressoffloodinggoingon

水淹胁迫时间/d株高净生长量/cm地径净生长量/mm根系活力/(μg·g-1·h-1)07.6±2.36a1.61±0.37d3794±174.8a74.3±0.77ab1.73±0.27d3707.5±151.9a143.1±0.45bc3.62±0.97c2723.0±14.0b213.1±1.40bc7.53±1.41a1831.1±126.9c281.5±0.92c5.54±0.81b400.6±48.9d
各处理株高和地径的净生长量均为28 d时的测量值。同一列数据后英文小写字母不同表示处理间某指标差异达显著水平(P<0.05)。
2.2 水淹胁迫对桢楠幼树叶绿体色素含量的影响
如表2所示,随着水淹胁迫时间的延长,桢楠幼树叶片叶绿素a(Chl a)含量、叶绿素b(Chl b)含量和叶绿素a/b (Chl a/b) 比值均呈下降趋势。在水淹14 ~28 d时,Chl a含量与CK差异显著(P<0.05);Chl b含量总体上呈下降趋势,但各处理之间差异不显著(P>0.05)。而类胡萝卜素含量随水淹时间延长呈先升后降的趋势,其含量的下降速率低于叶绿素的下降速率,导致类胡萝卜素/叶绿素比值显著增加(P<0.05)。
表2水淹胁迫对桢楠幼树叶片光合色素含量的影响
Table2EffectoffloodingoncontentofphotoplastpigmentsinleavesofPhoebezhennansaplings

水淹胁迫时间/dChla含量/(mg·g-1)Chlb含量/(mg·g-1)Chla/b比值类胡萝卜素含量/(mg·g-1)类胡萝卜素/叶绿素比值00.86±0.06a0.24±0.06a3.68±0.21a0.34±0.06b0.31±0.02c70.71±0.04ab0.20±0.01a3.48±0.21ab0.38±0.01b0.41±0.01b140.67±0.02bc0.22±0.03a3.10±0.23b0.49±0.05a0.53±0.03a210.53±0.01bc0.18±0.01a3.01±0.23b0.29±0.02bc0.41±0.01b280.41±0.03c0.16±0.12a2.58±0.61b0.26±0.05c0.45±0.08ab
同一列数据后英文小写字母不同表示处理间某指标差异达显著水平(P<0.05)。
2.3 水淹胁迫对桢楠幼树气体交换参数的影响
由图1可知,桢楠幼树各气体交换参数在不同水淹时间下均差异显著(P<0.05),其中,Pn、Gs和Tr均随着水淹胁迫时间的增加显著下降(P<0.05),在胁迫结束时(28 d),Pn、Gs和Tr分别比CK下降89.6%、98.7%、98.2%;Ci则随着胁迫时间的增加而显著增加(P<0.05)。此外,在水淹21 ~28 d时,Pn、Gs和Tr急剧下降,其中Gs和Tr已接近0,而Ci则急剧升高。Ls随着胁迫时间的延长而显著减小(P<0.05),0、7、14、21和28 d时的分别为0.456±0.08、0.380±0.01、0.235±0.05、0.179±0.03、-0.684±0.31。Ci的增加和Ls的减小表明非气孔限制因素成为抑制植株光合作用的主要因素,从光合生理过程看,受胁迫的植株已濒临死亡。

同一幅图中直方柱上方英文小写字母不同表示处理间某指标差异达显著水平(P<0.05)。
2.4 水淹胁迫对桢楠幼树叶片光合-光响应曲线的影响
由图2可知,桢楠幼树的光合-光响应曲线变化趋势符合植物光合作用的一般变化规律(图2)。但是水淹胁迫改变了这种变化规律,在同一光照强度下,随着水淹胁迫的加重,净光合速率(Pn)呈明显下降趋势。低光照强度下(≤100 μmol·m-2·s-1),不同水淹时间的桢楠幼树叶片Pn对光照强度的响应均非常敏感,随光照强度的增加迅速增加。当光照强度达100 μmol·m-2·s-1后,光响应过程产生了较大差异:CK、D7和D14仍保持较高的增加速率,直到光照强度上升到200 μmol·m-2·s-1后,D7和D14的Pn变化才趋于平缓,且分别在光照强度达600和400 μmol·m-2·s-1后趋于稳定;D21和D28则在光照强度达100 μmol·m-2·s-1后,Pn变化趋于平缓,仅在200 μmol·m-2·s-1时有所上升。

D0、D7、D14、D21、D28分别表示各处理持续水淹0、7、14、21、28 d,对应未被水淹的天数为 28、21、14、7、0 d。
由表3可知,桢楠幼树叶片Pn,max和Lsp随着水淹胁迫时间的延长而显著降低,其变化趋势与光响应曲线一致,而Lcp上升。受到水淹胁迫后,桢楠幼树叶片的Lsp降低和Lcp上升,表明桢楠幼树对光合有效辐射的有效利用范围减小,对光的利用能力也明显下降,不利于光合干物质的积累。
2.5 水淹胁迫对桢楠幼树叶片光合-CO2响应曲线的影响
由图3可知,桢楠幼树叶片的Pn随着CO2浓度升高的变化趋势也符合植物光合作用的一般规律,即随着CO2浓度的增加而上升。
表3不同持续水淹时间下桢楠幼树叶片光响应曲线和CO2响应曲线特征参数
Table3CharacteristicparametersoflightandCO2responsecurveofphotosynthesisinleavesofthePhoebezhennansaplingsunderstressrelativetodurationofthestress

水淹胁迫时间/d光响应曲线CO2响应曲线Pn,max/(μmol·m-2·s-1)Lcp/(μmol·m-2·s-1)Lsp/(μmol·m-2·s-1)Pn,max/(μmol·m-2·s-1)Ccp/(μmol·m-2·s-1)Csp/(μmol·m-2·s-1)06.49±0.05a24.74±9.12d482.86±34.06a33.63±0.49a83.20±9.26c2195.67±206.04a75.32±0.17b36.88±8.92cd391.34±32.44b24.64±1.73b103.87±1.16b2067.18±547.10a144.77±0.40c45.49±3.46bc339.40±16.05c23.54±1.00bc105.00±6.88b2042.92±347.33a212.80±0.12d63.70±0.69b281.59±18.95b19.83±0.50bcd142.65±7.27a2512.47±160.94a282.24±0.01e89.41±2.68a289.21±0.67b14.98±0.95d129.37±13.17a2022.01±395.15a
Pn,max为最大净光合速率;Lcp为光补偿点;Lsp为光饱和点;Ccp为CO2补偿点;Csp为CO2饱和点。同一列数据后英文小写字母不同表示处理间某指标差异达显著水平(P<0.05)。
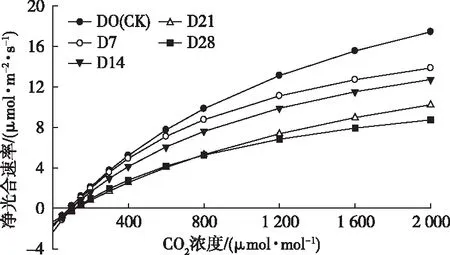
D0、D7、D14、D21、D28分别表示各处理持续水淹0、7、14、21、28 d,对应未被水淹的天数为28、21、14、7、0 d。
然而水淹胁迫显著降低了Pn随CO2浓度增加而上升的幅度。水淹胁迫时间越长,其上升的幅度越小,依次为CK> D7> D14> D21> D28。D21和D28比较而言,在CO2浓度为0~800 μmol·mol-1范围内,两者Pn值的上升幅度基本一致,当CO2浓度上升到800 μmol·mol-1后,D28的Pn值上升幅度明显减小;与CK相比,D14在CO2浓度上升到200 μmol·mol-1后Pn值上升幅度减小,而D7在CO2浓度上升到400 μmol·mol-1后Pn值上升幅度才开始减小,说明桢楠幼树水淹越严重,对CO2浓度越敏感,较低浓度的CO2即可影响植株的净光合速率。
由表3可知,桢楠幼树叶片Pn,max与CO2响应曲线变化趋势一致,D7、D14、D21、D28分别比CK显著降低26.7%、30%、41%、55.5%,说明水淹胁迫严重影响了桢楠幼树的最大光合能力。随着水淹时间的延长,Ccp显著上升,Csp却没有显著变化,表明水淹使桢楠幼树的CO2同化能力减弱。
2.6 水淹胁迫对桢楠幼树叶片净光合速率日变化的影响
不同水淹胁迫时间下桢楠幼树Pn日变化如图4所示。测定当日,光合有效辐射和叶片温度的日变化趋势相同,都呈单峰型曲线且最高峰出现在12:00;而桢楠幼树的光合日变化呈明显的双峰型曲线,且除D28外,其余各处理均在10:00和16:00出现峰值,12:00时Pn开始下降,在14:00时降到低谷,出现“午休”现象。此外,水淹胁迫对桢楠幼树Pn日变化有明显的抑制作用,且胁迫时间越长抑制作用越严重。
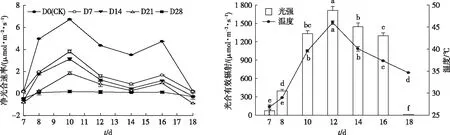
D0、D7、D14、D21、D28分别表示各处理持续水淹0、7、14、21、28 d,对应未被水淹的天数为 28、21、14、7、0 d。直方柱上方英文小写字母不同表示处理间某指标差异达显著水平(P<0.05)
3 讨论
3.1 水淹胁迫对桢楠幼树生长特性的影响
生长量是植物对逆境胁迫综合反应的直观指标,也是评价植物抗逆境能力的可靠指标[22]。在胁迫时间不超过7 d时,桢楠幼树株高和地径的增量变化不明显,这可能与根系在D7时还保持较高的活力有关。根系具有储存物质、吸收土壤水分和养分等功能,较高的根系活力有利于植物体吸收营养物质和合成干物质[23]。而在持续水淹胁迫14 d及更长的时间时,土壤中O2极度亏缺,无氧呼吸加强,CO2和O2-、H2O2、·OH 等活性氧累积,导致桢楠幼树体内各个生理代谢功能絮乱,严重威胁植物生长[24]。
3.2 水淹胁迫对桢楠幼树叶绿体色素及其光响应和CO2响应的影响
光合色素作为植物叶片光合作用的物质基础,其含量与净光合速率呈正相关,能反映植物光合作用的强弱[25]。该研究中,水淹胁迫下桢楠幼树叶片的叶绿体色素含量和比值均受到影响,即叶绿体色素总含量和Chl a/b比值均下降。虽然含量相对较高的Chl b具有更强的光能捕捉能力,但植物在逆境条件下生理代谢受阻,光能未能及时有效地用于光化学反应,反而增大叶绿体光能转化的负担,使光能富余过多而产生光抑制,加快具有破坏作用的活性氧产生[26]。类胡萝卜素含量在胁迫初期明显上升,可清除部分活性氧自由基,减小细胞膜脂过氧化,缓解水淹胁迫对植物的伤害[27],但随着胁迫时间的延长,这种防御机制遭到破坏,叶绿体色素含量下降,最后导致光合器官被破坏,影响光合作用。
光响应曲线与CO2响应曲线描述的是光照强度和CO2浓度与植物净光合速率之间的关系,能较好地解释植物对光照和CO2浓度2种影响因子变化的适应能力[21]。从桢楠幼树的光响应曲线和CO2响应曲线来看,水淹胁迫显著抑制了桢楠幼树的光合速率,根据植物对光和CO2适应理论标准[28],Lcp和Ccp上升,Lsp下降,导致桢楠幼树对环境中光和CO2的适应能力降低。曹晶等[29]对石楠的研究指出,光能利用率和同化受水淹影响较大,主要是因为受到胁迫后,细胞内活性氧大量增加,打破了植物体细胞内活性氧产生与清除的平衡机制,引起膜系统的损伤和膜透性的增加,造成叶绿素降解[30],对光合CO2的利用能力减弱从而导致光合速率下降。
3.3 水淹胁迫对桢楠幼树净光合速率及其日变化的影响
Pn是衡量叶片光合功能的重要指标。在胁迫初期(D7),桢楠幼树叶片的Pn急剧下降,Gs和Tr也显著下降,说明桢楠幼树光合作用对水淹胁迫非常敏感,在胁迫初期气孔关闭,阻碍外界CO2进入叶肉细胞,影响光合作用。但Ci在水淹胁迫下不断增加,这是因为细胞呼吸产生的CO2增加,且能够用于光合作用,所以与干旱胁迫下影响桢楠叶片Pn下降的因素不同[16],水淹胁迫时Pn下降并非由缺少CO2引起。由气孔因素的判断标准[31],即Ls与Ci的变化方向也可知桢楠幼树Pn的降低主要是由光合色素减少或叶肉细胞光合活性下降等非气孔因素所致。这与吴静等[32]研究鸡爪槭(Acerpalmatum)、张晔等[33]研究湿地松(Pinuselliottii)的结果相似。植物光合速率日变化受环境条件及其自身生理活性影响。该研究中,桢楠幼树净光合速率的日变化在水淹胁迫下受到明显抑制,这与桢楠幼树在逆境条件下对光能的利用效率低有关,也与温度有一定关系,邵玺文等[34]对黄芩(Scutellariabaicalensis)的研究中指出温度过高会影响叶肉细胞内光合关键酶的活力。桢楠幼树的净光合日变化速率在初期随着温度上升而增加,当温度上升到最高点时净光合速率开始下降,表明温度对净光合速率也有一定的影响,过高的温度会导致净光合速率下降,有机物质的合成减少,进而影响幼树生长。
综上所述,水淹胁迫使桢楠幼树叶片气孔关闭,光合色素含量降低,光合器官在一定程度上遭到破坏,导致其对光与CO2的利用能力降低,光合速率下降,营养物质积累减少,株高净生长量降低。结束胁迫后,水淹14 d以下的植株都能恢复生长,而水淹21 d的植株有部分死亡(即使恢复生长的植株,也只能从树干基部重新发芽),持续水淹28 d以上的植株则大量死亡。因此,在季节性洪涝灾害频繁发生的地区,尤其在华西雨屏区沟谷低洼地带,切忌大规模发展桢楠人工林。如果出于绿化目的,确需在局部地段栽植桢楠,则必须深垄整地,做好严格的排涝措施。
[1] 奉向阳,肖爱群,陈晓辉,等.楠木育苗与造林技术之浅谈[J].湖北林业科技,2013,42(1):87-88.[FENG Xiang-wei,XIAO Ai-quan,CHEN Xiao-qu,etal.Talking About the Seedling and Afforestation Techniques ofPhoebezhennan[J].Hubei Forestry Science and Technology,2013,42(1):87-88.]
[2] 丁鑫,肖建华,黄建峰,等.珍贵木材树种楠木的野生资源调查[J].植物分类与资源学报,2015,37(5):629-639.[DING Xin,XIAO Jian-hua,HUANG Jian-feng,etal.Investigation on Wild Resources of Valuable TimberPhoebezhenan(Lauraceae)[J].Plant Diversity,2015,37(5):629-639]
[3] 陈云霞,谢春平.华东地区楠木属植物的地理分布及资源保护对策[J].林业科技开发,2014,28(3):13-17.[CHEN Yun-xia,XIE Chun-ping.Geographical Distribution and Resource Protection Countermeasures ofPhoebeFrom East China[J].China Forestry Science and Technology 2014,28 (3):13-17.]
[4] 刘祖祺.植物抗性生理学[M].北京:中国农业出版社,1994:160-161.[LIU Zu-qi.Plant Resistance Physiology[J].Beijing:China Agriculture Press,1994:160-161.]
[5] JACKSON M B,ARMSTRONG W.Formation of Aerenchyma and the Processes of Plant Ventilation in Relation to Soil Flooding and Submergence[J].Plant Biology,1999,1(3):274-287.
[6] 李航.植物淹水胁迫对各生理特性的影响概述[J].安徽农业科学,2014,42(13):3802-3804.[LI Hang.Review on Effects of Waterlogging Stress on Plant Physiological Properties[J].Journal of Anhui Agricultural Sciences,2014,42 (13):3802-3804.]
[7] CHEN L,WANG W Q,PENG L.Photosynthetic and Physiological Responses ofKandeliacandelL.Druce Seedlings to Duration of Tidal Immersion in Artificial Seawater[J].Environmental and Experimental Botany,2005,54(3):256-266.
[8] THOMAS A L,GUERREIRO S M,SODEK L.Aerenchyma Formation and Recovery From Hypoxia of the Flooded Root System of Nodulated Soybean[J].Annals of Botany,2005,96(7):1191-1198.
[9] PAROLIN P.Morphological and Physiological Adjustments to Waterlogging and Drought in Seedlings of Amazonian Floodplain Trees[J].Oecologia,2001,128(3):326-335.
[10] TUITTILA E S,VASANDER H,LAINE J.Sensitivity of C Sequestration in ReintroducedSphagnumto Water-Level Variation in a Cutaway Peatland[J].Restoration Ecology,2004,12(4):483-493.
[11] 陈芳清,郭成圆,王传华,等.水淹对秋华柳幼苗生理生态特征的影响[J].应用生态学报,2008,19(6):1229-1233.[CHEN Fang-qing,GUO Cheng-yuan,WANG Chuan-hua,etal.Effects of Waterlogging on Ecophysiological Characteristics of Salix Variegate Seedlings[J].Chinese Journal of Applied Ecology,2008,19(6):1229-1233.]
[12] 谢腾芳,薛立,郭淑红,等.华南地区4种园林绿化树种的光合对水淹胁迫响应[J].安徽农业大学学报,2014,41(3):468-473.[XIE Teng-fang,XUE Li,GUO Shu-hong,etal.Photosynthetic Response of Four Landscaping Tree Species to Waterlogging Stress in South China[J].Journal of Anhui Agricultural University,2014,41 (3):468-473.]
[13] 王晓荣,胡兴宜,唐万鹏,等.模拟长江滩地水淹胁迫对3种树种幼苗生理生态特征的影响[J].东北林业大学学报,2015,43(1):45-49.[WANG Xiao-rong,HU Xing-yi,TANG Wan-peng,etal.Effects of Waterlogging Stress on Ecophysiological Characteristics of Three Species in Flooding Land of Yangtze River[J].Journal of Northeast Forestry University,2015,43(1):45-49.]
[14] 谭飞,周光良,胡红玲,等.不同基质配比对桢楠幼苗生长及其光合特性的影响[J].应用与环境生物学报,2016,22(5):823-830.[TAN Fei,ZHOU Guang-liang,HU Hong-ling,etal.Effects of the Substrate Ratio on the Growth and Photosynthesis Properties ofPhoebezhennanContainerized Seedlings[J].Chinese Journal of Applied and Environmental Biology,2016,22(5):823-830.]
[15] 贺维,胡庭兴,王锐,等.施肥对桢楠幼苗光合生理及生长特性的影响[J].西北植物学报,2014,34(6):1187-1197.[HE Wei,HU Ting-xing,WANG Rui,etal.Effect of Fertilization on Photosynthetic Physiology and Growth Characteristics ofPhoebezhennanSeedlings[J].Acta Botanica Boreali-Occidentalia Sinica,2014,34(6):1187-1197.]
[16] 王彬.干旱胁迫及复水对桢楠幼树生长和生理特性的影响[D].成都:四川农业大学,2015.[WANG Bin.Effects of Drought Stress and Re-Watering on Growth and Physilolgy ofPhoebezhennanSeedlings[D].Chengdu:Sichuan Agricultural University,2015.]
[17] 白宝璋,金锦子,白崧,等.玉米根系活力TTC测定法的改良[J].玉米科学,1994,2(4):44-47.[BAI Bao-zhang,JIN Jin-zhi,BAI Song,etal.Improvement of TTC Method Determining Root Activity in Corn[J].Journal of Maize Sciences,1994,2(4):44-47.]
[18] 李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2006:130-137.[LI He-sheng.Principles and Techniques of Plant Physiological and Biochemical Experiments[M].Beijing:Higher Education Press,2006:130-137.]
[19] 李永秀,申双和,李丽,等.土壤水分对冬小麦生育后期叶片气体交换及叶绿素荧光参数的影响[J].生态学杂志,2012,31(1):74-80.[LI Yong-xiu,SHEN Shuang-he,LI Li,etal.Effects of Soil Moisture on Leaf Gas Exchange and Chlorophyll Fluorescence Parameters of Winter Wheat at Late Growth Stage[J].Chinese Journal of Ecology,2012,31(1):74-80.]
[20] 叶子飘,于强.一个光合作用光响应新模型与传统模型的比较[J].沈阳农业大学学报,2007,38(6):771-775.[YE Zi-piao,YU Qiang.Comparison of a New Model of Photosynthetic Light Response With Traditional Models[J].Journal of Shenyang Agricultural University,2007,38(6):771-775.]
[21] 周光良,罗杰,胡红玲,等.干旱胁迫对巨桉幼树生长及光合特性的影响[J].生态与农村环境学报,2015,31(6):888-894.[ZHOU Gang-liang,LUO Jie,HU Hong-ling,etal.Effects of Drought Stress on Growth and Photosynthetic Characteristics ofEucalyptusGrandis Saplings[J].Journal of Ecology and Rural Enviroment,2015,31(6):888-894.]
[22] 罗杰,周光良,胡庭兴,等.干旱胁迫对润楠幼苗生长和生理生化指标的影响[J].应用与环境生物学报,2015,21(3):563-570.[LUO Jie,ZHOU Guang-liang,HU Ting-xing,etal.Effects of Drought Stress on Growth and Physiological and Biochemical Indexes ofMachiluspingiiSeedlings[J].Chinese Jouranl of Applied and Environmental Biology,2015,21(3):563-570.]
[23] 魏道智,宁书菊,林文雄.小麦根系活力变化与叶片衰老的研究[J].应用生态学报,2004,15(9):1565-1569.[WEI Dao-zhi,NING Shu-ju,LING Wen-xiong.Relationship Between Wheat Root Activity and Leaf Senescence[J].Chinese Journal of Applied Ecology,2004,15(9):1565-1569.]
[24] 陈张焕,黄青,于进英,等.高温及水淹胁迫对薰衣草生理生化特性的影响[J].湖南农业科学,2012(9):46-50.[CHEN Zhang-huan,HUANG Qing,YU Jin-ying,etal.Effects of High Temperature and Waterlogging Stress on Physiological and Giochemical Characteristics ofLavandulaiatifoliaMedik[J].Hunan Agricultural Sciences,2012(9):46-50.]
[25] 孙小玲,许岳飞,马鲁沂,等.植株叶片的光合色素构成对遮阴的响应[J].植物生态学报,2010,34(8):989-999.[SUN Xiao-ling,XU Yue-fei,MA Lu-yi,etal.A Review of Acclimation of Photosynthetic Pigment Composition in Plant Leaves to Shade Environment[J].Chinese Journal of Plant Ecology,2010,34(8):989-999.]
[26] 王宗星,吴统贵,虞木奎.植物光合特性对水淹胁迫的响应机制[J].世界林业研究,2010,23(5):20-24.[WANG Zong-xing,WU Tong-gui,YU Mu-kui.Response Mechanism of Plant Photosynthesis Characteristic to Flooding Stress[J].World Forestry Research,2010,23(5):20-24.]
[27] 费小红,安保光,赵宝华,等.类胡萝卜素参与水稻抗氧化胁迫的研究进展[J].现代农业科学,2009(1):22-23.[FEI Xiao-hong,AN Bao-guang,ZHAO Bao-hua,etal.Carotenoid Antioxidant Coercion to Participate in Rice Research[J].Modern Agricultura Sciences,2009(1):22-23.]
[28] 韩刚,赵忠.不同土壤水分下4种沙生灌木的光合光响应特性[J].生态学报,2010,30(15):4019-4026.[HAN Gang,ZHAO Zhong.Light Response Characteristics of Photosynthesis of Four Xerophilous Shrubs Under Different Soil Moistures[J].Acta Ecologica Sinica,2010,30(15):4019-4026.]
[29] 曹晶,姜卫兵,翁忙玲,等.夏秋季旱涝胁迫对红叶石楠光合特性的影响[J].园艺学报,2007,34(1):163-172.[CAO Jing,JIANG Wei-bing,WENG Mang-ling,etal.Effects of Drought and Flooding Stress on Photosynthetic Characteristics ofPhotiniafraseryin Summer and Autumn[J].Acta Horticulturae Sinica,2007,34(1):163-172.]
[30] 李德明,张秀娟,陈娟.涝渍对植物光合作用的影响及其生理危害[J].北方园艺,2010(5):210-212.[LI De-ming,ZHANG Xin-juan,CHEN Juan.Photosynthesis and Related Physiological Characteristica Affected by Waterlogging in Plant[J].Northern Horticulture,2010(5):210-212.]
[31] 许大全.光合作用气孔限制分析中的一些问题[J].植物生理学通讯,1997,33(4):241-244.[XU Da-quan.Some Problems in Stomatal Limitation Analysis of Photosynthesis[J].Plant Physiology Communications,1997,33(4):241-244.]
[32] 吴静,王小德,叶建峰,等.涝渍胁迫对鸡爪槭幼苗光合生理特性的影响[J].西北林学院学报,2014,29(5):26-30.[WU Jing,WANG Xiao-de,YE Jian-feng,etal.Effects of Waterlogging Stress on Photosynthetic Physiological Characteristics ofAcertruncatumSeedlings[J].Journal of Northwest Forestry University,2014,29 (5):26-30.]
[33] 张晔,李昌晓.水淹与干旱交替胁迫对湿地松幼苗光合与生长的影响[J].林业科学,2011,47(12):158-164.[ZHANG Ye,LI Chang-xiao.Effects of Submergence and Drought Alternation on Photosynthesis and Growth ofPinuselliottiiSeedlings[J].Scientia Silvae Sinicae,2011,47(12):158-164.]
[34] 邵玺文,韩梅,韩忠明,等.不同生境条件下黄芩光合日变化与环境因子的关系[J].生态学报,2009,29(3):1470-1477.[SHAO Xi-wen,HAN Mei,HAN Zhong-ming,etal.Relationship Between Diurnal Changes of Photosynthesis ofScutellariabaicalensisand Environmental Factors in Different Habitats[J].Acta Ecologica Sinica,2009,29(3):1470-1477.]
