硒对油菜幼苗铜胁迫的缓解效应
2018-03-27崔雪梅李冰晖李春生郭海如
崔雪梅, 李冰晖, 李春生, 郭海如
(湖北工程学院生命科学技术学院,湖北孝感 432000)
硒(Se)是植物生长发育过程中的有益元素[1],也是生物所必需的营养元素,对人和动物的抗氧化系统起着关键作用。Se对植物生长发育的影响具有二重性,低浓度的Se能促进一些植物的萌发及幼苗生长,提高作物产量与品质,促进植物新陈代谢和提高植物对环境胁迫的抗性[2];而过量的Se对植物的生长发育有毒害作用[3]。环境中的Se不足或过量都会通过食物链而影响人类的健康,所以,有关Se对植物的影响受到了普遍关注,但Se对不同植物表现出的生理学差异还有待深入研究。
铜(Cu)是植物生长发育所必需的一种微量元素,但植物正常生长所需要的Cu量很少,当过量Cu被植物吸收后,会对植物产生毒害作用,抑制植物的生长发育甚至使植物死亡。目前,农业生产活动中使用大量高铜杀菌剂、污水灌溉等多种原因导致土壤中Cu污染日益加剧[4]。
油菜是我国主要的油料作物,种植面积和产量均居世界首位[5]。油菜种植不仅能收获菜籽生产食用油,还可以采摘菜薹作为蔬菜,而且其饼粕富含蛋白质,可作为动物饲料,是一种倍受青睐的作物[6]。要使油菜籽和菜薹的产量高、质量好,势必要注意油菜的生长环境等诸多因素。目前,虽有探讨硒、铜对油菜生长的影响方面的报道,但多集中在单独一种元素对油菜生长的影响上。王波等在恒温箱培养条件下研究了铜胁迫对油菜种子萌发期生理生化指标的影响,结果表明,低浓度的铜对油菜有促进作用,而高浓度的铜则产生了抑制作用[7];张驰等应用土培与水培相结合的盆栽方式,研究了硒对油菜苗期生长和生理生化指标的影响,结果表明,硒能明显影响油菜的生长发育[8-9],适量的补硒处理具有明显的增产效果[10]。而关于油菜对Se、Cu交互作用的生理响应机制还缺乏系统研究。本研究拟采用不同浓度的Se、Cu溶液处理盆栽油菜幼苗,处理结束后测定幼苗叶绿素、丙二醛(MDA)、硝态氮、可溶性糖、蛋白质等指标含量及过氧化物酶活性、淀粉酶活性,以揭示Se、Cu交互胁迫下油菜的生理响应规律,从而找出最佳的Se、Cu供应比例,为油菜品质的改善和产量的提高提供参考。
1 材料与方法
1.1 试验方法
本试验于2015年10月至2016年1月在湖北工程学院生命科学技术学院生科楼进行。先用1%次氯酸钠溶液对油菜种子进行15 min的消毒处理,然后用无菌水冲洗5次,再用蒸馏水浸泡种子24 h,让其充分吸水后挑选沉于水下方的籽粒饱满的种子若干,将选好的种子均匀撒在铺有2层滤纸的白色托盘内,用适量蒸馏水湿润滤纸(水量以保持滤纸湿润,但不宜流动即可),将托盘放入MJX-250B-Z型恒温箱进行催芽,温度设置为25 ℃,其间每天向托盘中添加适量、等量蒸馏水,3 d后种子露白,再移入花盆中进行土培试验。
取养分贫瘠的沙土,用筛子筛出沙土中较大的石砾等杂质。将土均匀地分到45个盆中,浇适量、等量蒸馏水保持土壤湿润。
将发芽后的油菜种子移至花盆中,每盆10株,均匀分布。将花盆放在向阳通风处,每天浇2次等量Hoagland’s营养液[11]。待油菜幼苗长至4~5张叶片之后,将45盆油菜随机分成3组,每组3个重复,进行Se、Cu胁迫处理。
1.2 试验设计
Cu原液主要成分为CuSO4,浓度设置为0、25、75、100、200 mg/L,Se原液主要成分为Na2SeO3,浓度设置为0、12、22 mg/L。试验具体设计见表1。
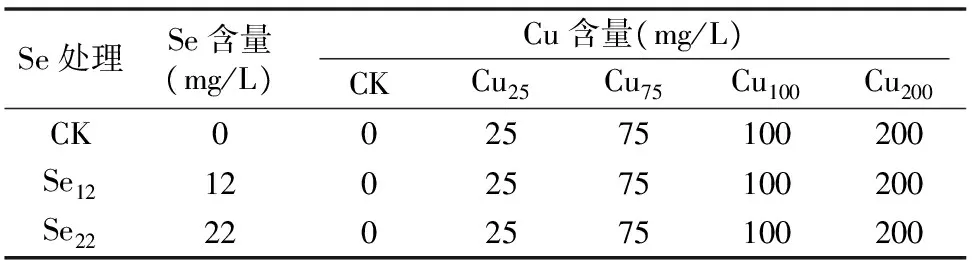
表1 试验设计
1.3 指标测定方法
胁迫处理25 d后,取油菜样品测定各项生理指标。主要指标测定方法[12-16]如表2所示。

表2 各指标测定方法
1.4 数据处理方法
采用Excel进行数据计算与处理,对数据进行无重复双因素方差分析。
2 结果与分析
2.1 供硒对铜胁迫油菜中叶绿素含量的影响
叶绿素是植物进行光合作用的主要色素,是反映植物叶片光合作用能力的重要指标[17]。胁迫处理结束后,测定油菜叶片总叶绿素含量。由图1可看出:(1)单独喷施Cu溶液时,油菜幼苗总叶绿素含量随着Cu浓度的增加先增加后降低。当Cu浓度达到75 mg/L时,总叶绿素含量达最高值,而Cu浓度过高(200 mg/L)时,则产生一定的抑制作用;同样,单独喷施Se时,当Se浓度为12 mg/L时,总叶绿素含量最高,而当Se浓度为22 mg/L时,总叶绿素含量低于对照。由此可知,低浓度Se、Cu对油菜合成叶绿素有促进作用,而高浓度Se、Cu则会产生抑制作用。(2)Se、Cu同时供应时,低浓度Cu(≤75 mg/L)、低浓度Se(12 mg/L)配施,油菜叶绿素含量均高于对照和高硒组。当Cu浓度超过100 mg/L时,高Se浓度(22 mg/L)组总叶绿素含量均高于对照和低Se组。说明适量的Se、Cu对叶绿素含量有促进作用,当环境中的Cu浓度高于100 mg/L时,喷施高浓度的Se对Cu胁迫有较好的缓解作用。
由表3可知:(1)以Se浓度为差异源时,F值>F临界值,说明Se对油菜中总叶绿素含量的影响达到显著水平;(2)以Cu浓度为差异源时,F值>F临界值,说明Cu对油菜总叶绿素含量的影响也达到显著水平;(3)差异源是Se、Cu的交互作用时,F值>F临界值,说明Se和Cu的交互作用达到显著水平,即Se和Cu在共同影响油菜总叶绿素含量方面有明显的交互作用。
2.2 供硒对铜胁迫油菜中丙二醛浓度的影响
丙二醛是常用的膜脂过氧化指标,在植物衰老生理和抗性生理研究中,MDA浓度是一个常用指标,可以通过MDA了解膜脂过氧化的程度,以此判断植物的膜系统受损程度及其抗逆性。胁迫处理结束后,测定油菜叶片中MDA浓度。由图2可看出:(1)无论喷Se浓度为多少,MDA浓度均随Cu浓度的增加而呈现先减少后增加的趋势,都在Cu浓度为 100 mg/L 时达到最小值,说明低浓度的Cu可以减少MDA的产生,增强了植物的抗逆性。另外,当单独供Se时,植物叶片中MDA浓度在Se浓度为12 mg/L时最低,说明低浓度的Se能增强植物的抗逆性。(2)当Cu浓度≤100 mg/L时,Se浓度为22 mg/L时叶片中MDA浓度低于对照组、低Se组,说明当环境中Cu浓度较低时,高浓度Se对油菜的Cu胁迫缓解效果较好。(3)而当Cu浓度为200 mg/L时,则表现为12 mg/L 浓度Se的缓解效果更佳,说明低浓度Se对高浓度的Cu胁迫缓解效果更佳。
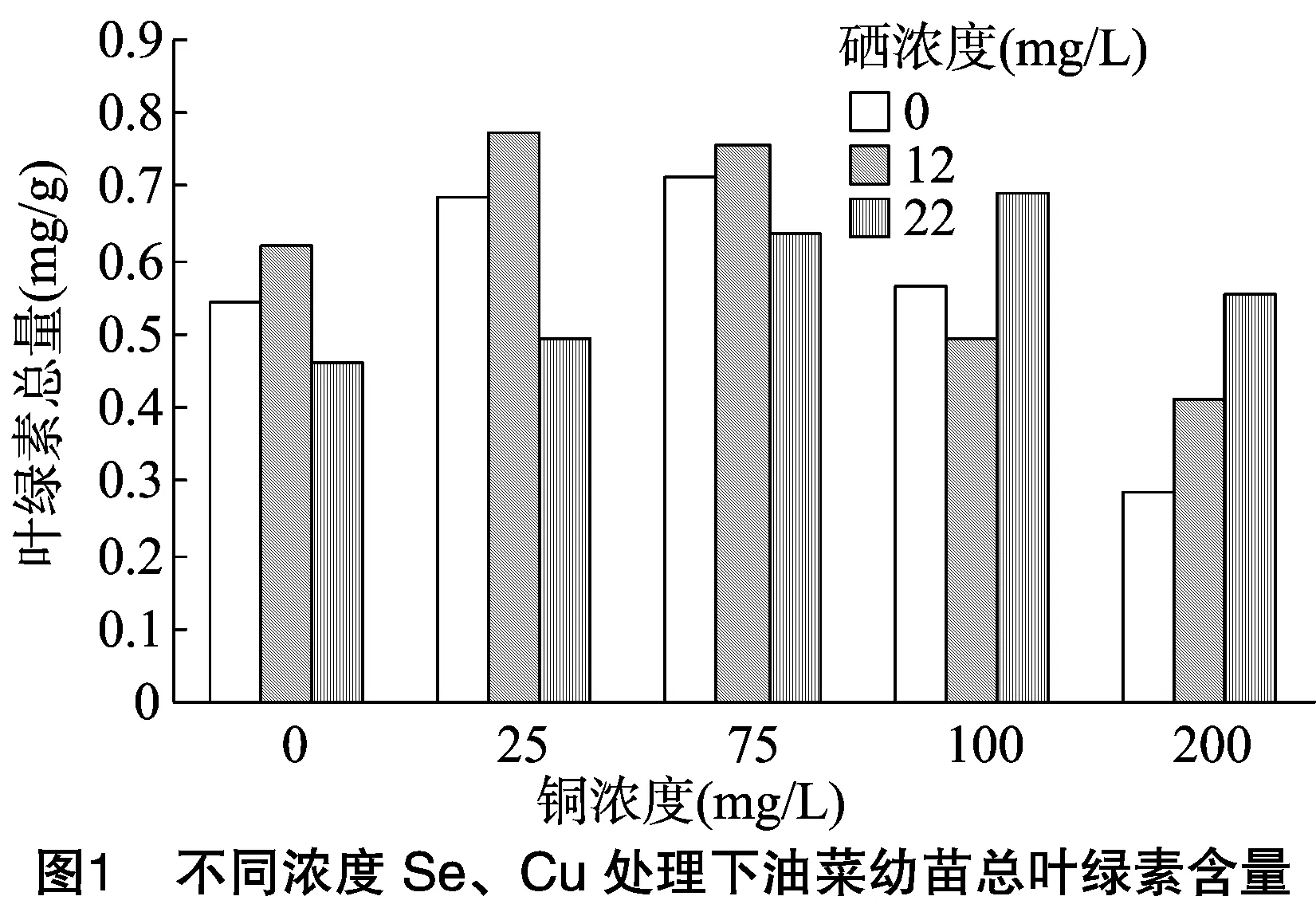
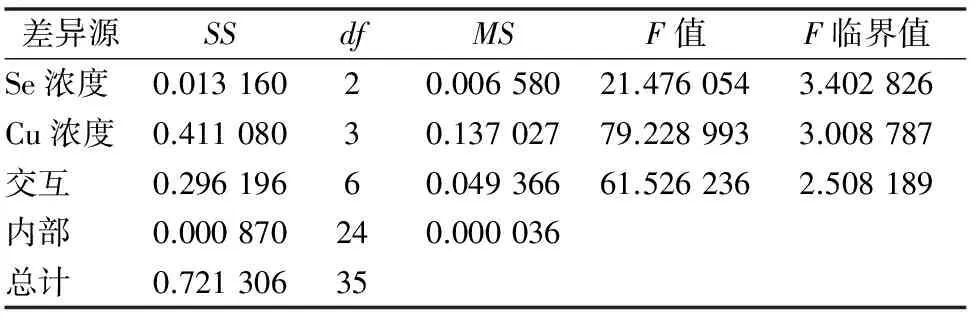
表3 总叶绿素含量变化方差分析结果
注:α=0.05;SS为方差,df为自由度,MS为均方。下表同。
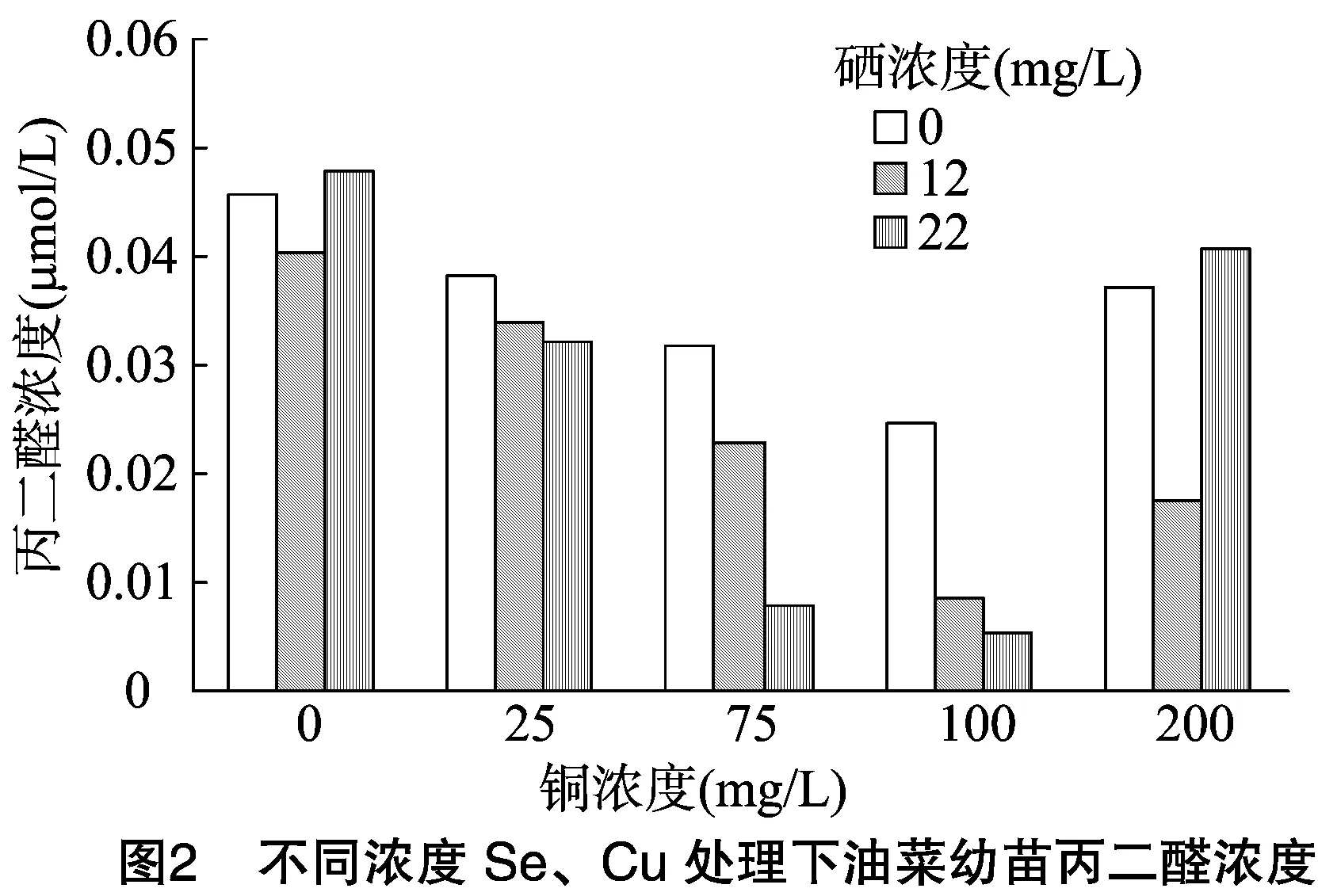
由表4可知:(1)以Se浓度为差异源时,F值>F临界值,说明Se对油菜中MDA浓度的影响达到显著水平;(2)以Cu浓度为差异源时,F值>F临界值,说明喷Cu对油菜MDA浓度的影响也达到显著水平;(3)当差异源是Se、Cu交互作用时,F值>F临界值,说明Se和Cu的交互作用达到显著水平,即Se、Cu在对油菜MDA浓度影响方面有明显的交互作用。
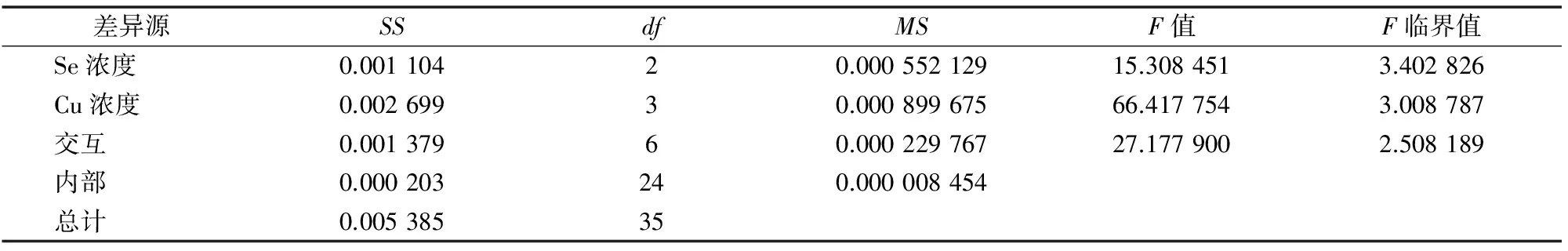
表4 丙二醛浓度变化方差分析结果
2.3 供硒对铜胁迫油菜中蛋白质含量的影响
胁迫处理结束后,测定油菜叶片中蛋白质的含量。由图3可知:(1)当单独供Cu时,油菜中蛋白质含量随Cu浓度的升高而呈现先增加后减少的趋势,当Cu浓度为75 mg/L时达到最大值,说明低浓度的Cu对油菜合成蛋白质有一定的促进作用,当Cu浓度为200 mg/L时,蛋白质含量明显低于对照组,表明高浓度的Cu产生抑制作用。(2)无论是单独供Se还是Se、Cu同时供应,都表现为低浓度Se(12 mg/L)条件下蛋白质含量最高,说明低浓度Se(12 mg/L)对Cu胁迫油菜有一定的促进作用,这种促进作用在Cu浓度为25 mg/L时达到最佳效果。当铜浓度为200 mg/L时,供Se都在一定程度上缓解了高浓度铜的胁迫。
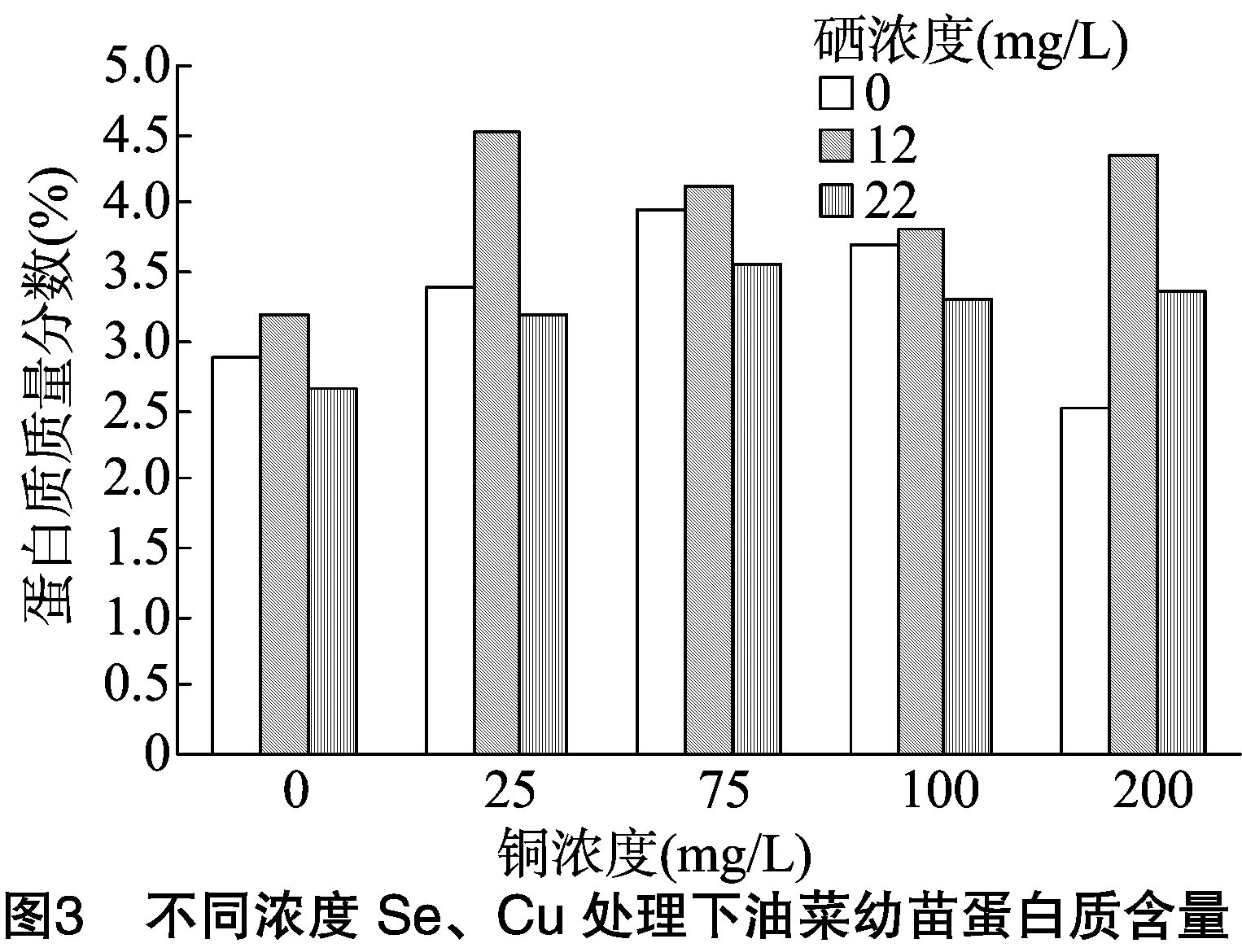
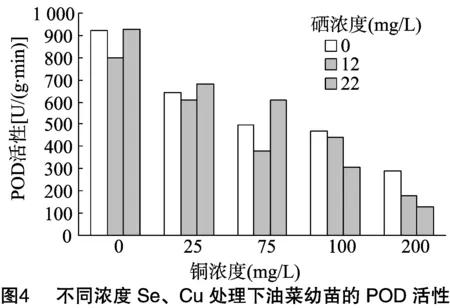
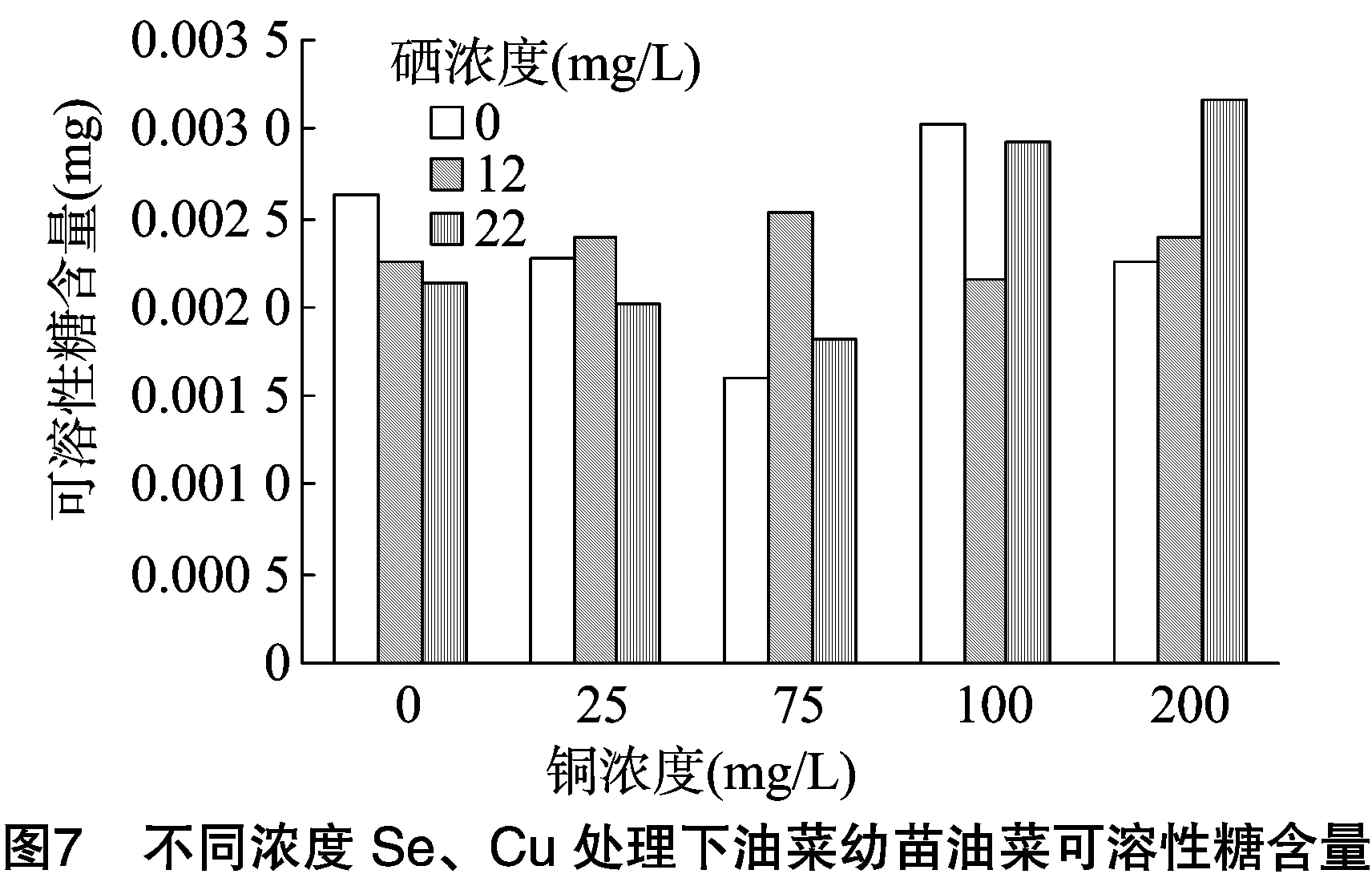
对蛋白质含量进行双因素方差分析,由表5可知:(1)以Se浓度为差异源时,F值 表5 蛋白质含量方差分析结果 植物体中含有大量过氧化物酶,这是活性较高的一种酶,它与呼吸作用、光合作用及生长素的氧化等都有关系。胁迫处理结束后,测定油菜叶片中的POD活性。由图4可看出:(1)在Se浓度为0和高Se浓度(22 mg/L)处理下,随着Cu浓度的增加,油菜中POD活性逐渐降低;(2)在低Se浓度(12 mg/L)处理下,油菜中POD活性随Cu浓度增加呈现先降低后升高再降低的趋势。结果说明,适量Se对Cu胁迫下油菜的POD活性抑制有一定的缓解作用。 对蛋白质含量进行双因素方差分析,由表6可知:(1)以Se浓度为差异源时,F值 表6 油菜POD活性方差分析结果 测定植物体内的硝态氮含量,能反映植物的氮素营养状况,还可为鉴定蔬菜及其加工品的品质提供依据。胁迫处理结束后,测定油菜叶片中硝态氮的含量。由图5可看出:(1)不供Se时,油菜中硝态氮含量随Cu浓度的增加而逐渐升高,当Cu浓度≤75 mg/L时,硝态氮含量变化不明显,当Cu浓度>100 mg/L 时,硝态氮含量开始急剧增加。说明低浓度Cu对油菜硝态氮含量影响较小,高浓度Cu会促进油菜硝态氮的累积。(2)在Se、Cu同时供应的条件下,硝态氮含量均在一定范围内先增加后降低,在Se浓度为12 mg/L、Cu浓度为100 mg/L组合下,硝态氮含量最低。当Cu浓度为 200 mg/L 时,低浓度Se组和高浓度Se组油菜的硝态氮含量都明显低于对照,说明Se能明显缓解高浓度Cu胁迫下的硝态氮含量的升高。 对硝态氮含量进行双因素方差分析,由表7可知:(1)以Se浓度为差异源时,F值>F临界值, 说明Se对油菜中硝态氮含量的影响没有达到显著水平;(2)以Cu浓度为差异源时,F值>F临界值,说明Cu对油菜硝态氮含量的影响达到了显著水平;(3)当差异源是Se、Cu交互作用时,F值>F临界值,说明Se和Cu的交互作用达到显著水平,即Se和Cu在共同影响油菜硝态氮含量方面的交互作用非常明显。 植物的能量来源主要是淀粉, 所以需要淀粉酶来分解淀粉获得能量,以提供组织生长需要以及完成各种生理活动。淀粉酶活性的高低常作为植物抗逆性的生化指标。胁迫处理结束后,测定油菜幼苗中淀粉酶的活性。由图6可看出:(1)当单独供Se时,油菜淀粉酶活性随着Se浓度的增加而增加, 表7 硝态氮含量方差分析结果 说明Se对油菜淀粉酶活性有促进作用;(2)单独供Cu时,随着Cu浓度的增加,淀粉酶活性先增加后降低,在Cu浓度为75 mg/L时达到最高值,此后虽然有降低,但仍高于对照组,说明在此浓度范围内,Cu对油菜淀粉酶活性均有促进作用,且低浓度Cu的促进效果更明显;(3)当Se、Cu同时供应时,表现为高浓度Se(22 mg/L)与低浓度Cu(75 mg/L)组合下的淀粉酶活性最高。由结果可知,低浓度Se对高浓度Cu胁迫下的淀粉酶活性有一定的缓解作用。 对淀粉酶活性进行双因素方差分析,由表8可知:(1)以Se浓度为差异源时,F值 表8 淀粉酶活性方差分析结果 糖类作为呼吸基质,为作物的各种合成过程和各种生命活动提供了所需的能量。测定蔬菜作物中可溶性糖含量,可以了解和鉴定作物品质的高低。胁迫处理结束后,测定油菜叶片中可溶性糖含量,由图7可以看出:(1)单独供Cu时,油菜可溶性糖含量随Cu浓度增加呈现先降后升再降的变化趋势。在Cu浓度为100 mg/L时达到最大值,Cu浓度为 200 mg/L 时,可溶性糖含量明显低于对照,说明低浓度的Cu对油菜有促进作用,高浓度则产生抑制。(2)单独供Se时,油菜可溶性糖含量呈下降趋势。Se、Cu同时供应时,低浓度Se(12 mg/L)与低浓度Cu(75 mg/L)会提高可溶性糖含量,随着Cu浓度的增加,则表现为高浓度Se(22 mg/L)与高浓度Cu组合会提高可溶性糖的含量。结果表明,低浓度Se与低浓度Cu对油菜可溶性糖含量有促进作用,而高浓度Se能有效缓解高浓度的Cu胁迫。 对可溶性糖含量进行双因素方差分析,由表9可知:(1)以Se浓度为差异源时,F值 表9 可溶性糖含量方差分析 本研究对盆栽油菜进行不同浓度Se、Cu单施和配施试验,分析幼苗叶绿素、丙二醛、硝态氮、可溶性糖、蛋白质等指标的含量及过氧化物酶活性、淀粉酶活性的变化规律。结果表明:(1)单独喷施Cu溶液时,油菜叶绿素、蛋白质含量及淀粉酶活性均在Cu浓度为75 mg/L时达最高值,此后逐渐降低;而可溶性糖含量在铜浓度为100 mg/L时达最高值,当Cu浓度为200 mg/L时,大部分指标表现为受抑制。另外,Se对POD活性影响较小,而Cu对POD活性的影响较明显。王波等研究铜溶液对几种油菜种子萌发期的影响时也得出,低浓度铜对萌发期可溶性糖含量及淀粉酶活性有促进作用,高浓度铜溶液抑制种子的萌发[9]。(2)单独喷施Se溶液时,油菜叶绿素、MDA、蛋白质含量均在硒浓度为12 mg/L时效果最佳,其余部分指标表现为Se浓度为22 mg/L时效果最佳,与对照相比,供Se组各项指标都较好。刘新伟等在水培环境下研究硒对油菜生长的影响时也得出,少量供硒对油菜的生长有促进作用[18-19]。(3)Se、Cu同时供应时,低浓度Se(12 mg/L)与低浓度Cu(75 mg/L)的组合,高浓度Se(22 mg/L)与高浓度Cu(>100 mg/L)的组合对油菜生理指标的综合影响较好。说明低浓度的Se、Cu配施,对油菜各项生理指标均有促进作用,而当环境中的Cu浓度超过 100 mg/L 时,表现为较高浓度的Se能有效缓解Cu对油菜的胁迫作用。 综上所述,对各项指标进行双因素方差分析表明,单独喷Se溶液,对油菜中总叶绿素和MDA含量的影响达到显著水平,而对其他5项指标的影响均未达显著水平;单独喷Cu溶液时,Cu对油菜总叶绿素含量、MDA浓度、POD活性、硝态氮含量、淀粉酶活性影响均达显著水平,仅对可溶性糖、蛋白质含量的影响为不显著;同时喷施Se、Cu溶液时,对大部分指标影响均达显著水平。说明硒Se对Cu胁迫有较好的缓解作用。 目前,还没有关于硒、铜交互胁迫下油菜的生理响应研究。但已有文献报道了低浓度的硒可以缓解西葫芦铅胁迫的毒害,高浓度的硒与铅胁迫存在协同互作而使西葫芦毒害程度加重[1]。本试验在盆栽环境下研究了硒、铜交互作用下油菜的蛋白质含量等生理指标、酶活性的变化规律,确定了最佳的硒、铜配施比例,这不仅对提高粮油产量和品质具有重要意义,也为农业生产实践提供了重要的科学依据。 [1]王彩霞,吕有军. 硒对铅胁迫下西葫芦幼苗膜系统和抗氧化酶的影响[J]. 种子,2015,34(8):14-17. [2]彭 玲,贾 芬,田小平,等. 硒对油菜根尖镉胁迫的缓解作用[J]. 环境科学学报,2015,35(8):2597-2604. [3]张美德,艾伦强,何银生,等. 硒对镉胁迫下白术抗氧化能力的影响[J]. 南方农业学报,2014,45(7):1211-1214. [4]朱 健,张志红,范菲菲,等. 铜胁迫对海菜花幼苗生理特征的影响[J]. 江苏农业学报,2015,31(4):883-886. [5]崔雪梅,郭海如,方 嫱,等. 基于主成分分析的油菜盐害生理反应规律[J]. 江苏农业科学,2016,44(2):105-108. [6]刘凤兰,杨经泽,李光明,等. 双低甘蓝型油菜菜薹、菜子兼用高效技术研究[J]. 湖北农业科学,2005(2):35-36,43. [7]王 波,陈 辰,张 磊. 铜胁迫对油菜种子萌发期生理生化指标的影响[J]. 中国农学通报,2009,25(4):137-139. [8]张 驰,刘信平,周大寨,等. 油菜苗期对硒的生物富集作用动态研究[J]. 江苏农业科学,2007(4):21-25. [9]张 驰,吴永尧,彭振坤,等. 油菜苗期对硒的生物富集分布[J]. 河南农业科学,2005(9):29-32. [10]张 驰,周大寨,吴永尧,等. 硒对油菜苗期生长和生理生化指标的影响[J]. 湖北农业科学,2007,46(3):363-365. [11]高晓旭,张志刚,段 颖,等. 高浓度营养液对黄瓜和番茄下胚轴徒长的抑制作用[J]. 植物营养与肥料学报,2014,20(5):1234-1242. [12]李合生. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2000:89-90. [13]赵世杰,许长成,邹 琦,等. 植物组织中丙二醛测定方法的改进[J]. 植物生理学通讯,1994,30(3):207-210. [14]邹 琦. 植物生理学实验指导[M]. 北京:中国农业出版社,2000:223-224. [15]张志良. 植物生理学实验指导[M]. 北京:高等教育出版社,2012:121-123. [16]武 晶,杜新民. 硒对西葫芦幼苗根系生长及保护酶活性的影响[J]. 黑龙江农业科学,2014(11):88-90. [17]史典义,刘忠香,金危危. 植物叶绿素合成、分解代谢及信号调控[J]. 遗传,2009,31(7):698-704. [18]刘新伟,王巧兰,段碧辉,等. 亚硒酸盐对油菜幼苗硒吸收、根系形态及生理指标的影响[J]. 应用生态学报,2015,26(7):2050-2056. [19]杨文秀,王 萍,杨晓宇,等. 叶面施硒对油菜产量及品质的影响[J]. 内蒙古农业大学学报(自然科学版),2010,31(3):88-90.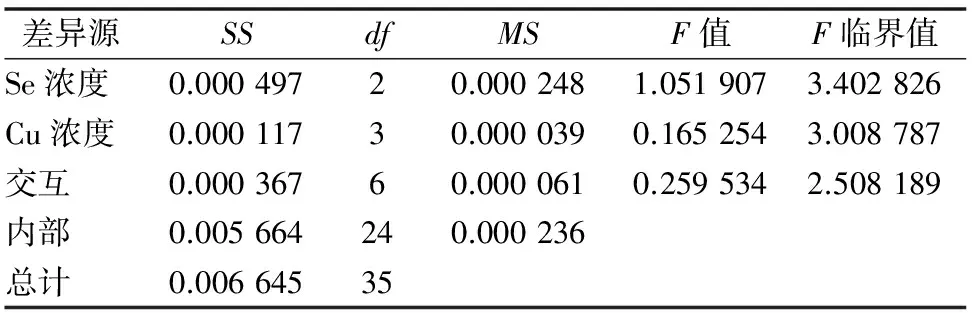
2.4 供硒对铜胁迫油菜中POD活性的影响
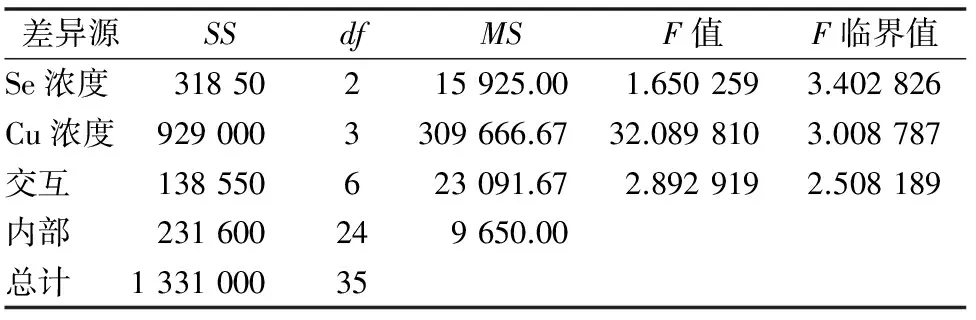
2.5 供硒对铜胁迫油菜中硝态氮含量的影响
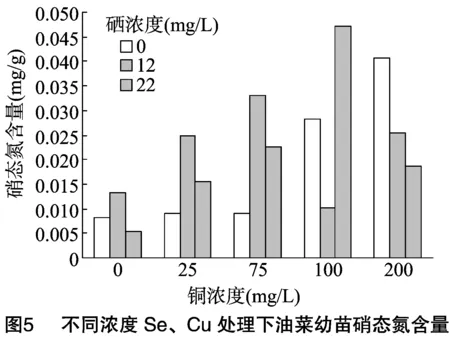
2.6 供硒对铜胁迫油菜中淀粉酶活性的影响

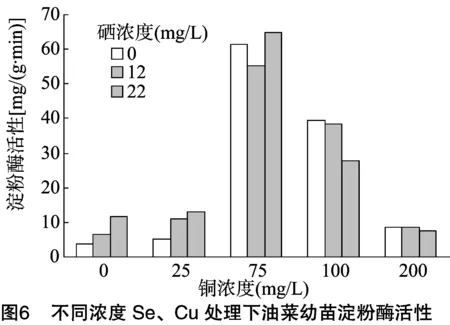
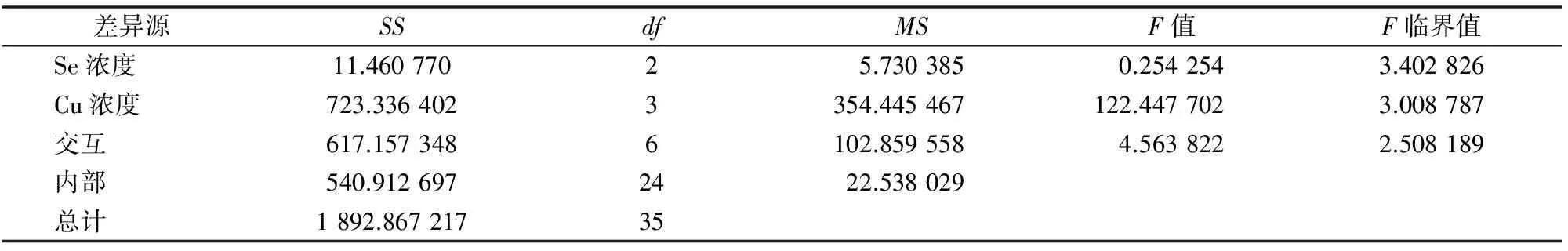
2.7 供硒对铜胁迫油菜中可溶性糖含量的影响

3 结论与讨论
