转cry1Ab/cry2Aj和G10evo-epsps基因玉米中Bt蛋白的时空表达及抗性评价
2018-03-26孙红炜徐晓辉杨淑珂路兴波
孙红炜, 李 凡, 高 瑞, 徐晓辉, 杨淑珂, 路兴波
山东省农业科学院植物保护研究所/山东省植物病毒学重点实验室,山东 济南 250100
玉米是我国种植面积最大的粮食作物,2016年中国玉米种植面积3676万hm2,在我国粮食及饲料工业中占有举足轻重的作用。亚洲玉米螟Ostriniafurnacalis(Guenée)、黏虫Mythimnaseparata(Walker)和棉铃虫Helicoverpaarmigera(Hübner)是影响玉米产量的三大主要鳞翅目害虫,在个别年份,由于害虫的危害,甚至会造成玉米绝收。如2012年在吉林省、2016年在陕西渭南,均发生严重的玉米黏虫为害(聂继东和汪洪伟,2017; 王红军和郭书亚,2017; 王振营和鲁新,2010)。我国玉米害虫防治主要依赖化学防治,化学防治虽然效果较好,但存在防治滞后、无法挽回实际损失的缺点。另外,化学防治不当容易造成环境、食品污染,同时浪费大量人力、物力。
1996年转基因玉米在美国首次商品化,转基因作物在全球蓬勃发展,其中转Bt基因玉米是目前商品化生产发展最快的转基因抗虫作物之一(James,2016)。转Bt基因玉米通过持续表达外源Bt蛋白,在整个生育期都可以对靶标生物产生抑制或抗性效果,可以大大减少人力、物力的投入,且安全高效。国外应用转基因抗虫玉米已超过20年,对靶标害虫表现了良好的抗性(Burknessetal.,2001; Lynchetal.,1999; Williamsetal.,1998)。我国转基因玉米研发虽然起步较晚,但发展迅速,在Bt基因筛选、改造和合成方面取得很多专利成果,且有为数众多的转Bt基因玉米进行到中间试验或环境释放试验阶段(王延峰,2010, 岳同卿等,2010)。转基因玉米双抗12-5是浙江大学研发的含有抗虫基因cry1Ab/cry2Aj与耐除草剂基因G10evo-epsps的转基因玉米新品系,cry1Ab/cry2Aj对亚洲玉米螟有较高的杀虫活性,草甘膦靶标酶基因G10evo-epsps对草甘膦具有很高的耐受性。
转基因作物在推广之前,必须经过环境安全性评价,如抗虫性、除草剂耐受性、生存竞争能力、对农田生物多样性的影响等(姜伟丽等,2013; 马小艳等,2013)。本试验利用ELISA法测定了转基因玉米双抗12-5转育后获得的玉米材料双抗505-12-5中Bt蛋白在不同生育期不同组织中的Bt蛋白表达量,并对其室内及田间抗虫性进行了研究,为转基因玉米的基础研究、推广应用以及害虫防治方面提供依据。
1 材料与方法
1.1 供试玉米
转基因玉米双抗505-12-5和非转基因对照玉米中科玉505,由浙江大学提供;常规品种登海3号,购自当地种子市场;田间抗虫性鉴定感虫品种综31由中国农业科学院作物科学研究所提供。
1.2 试验方法
1.2.1 试验设计 每种玉米材料设3次重复,小区面积300 m2(50 m×6 m),行距60 cm,株距25 cm,小区间设2 m间隔,以防害虫在不同小区之间扩散。Bt蛋白含量测定、鳞翅目害虫室外及室内抗虫性鉴定均在相应的小区接虫或取样。
1.2.2 Bt蛋白表达量检测样品采集及前期处理 在转基因玉米双抗505-12-5的6~8叶期、大喇叭口期、吐丝期、乳熟期取根、茎、心叶,吐丝期取花丝,乳熟期取幼嫩籽粒。每小区5点取样,每点选取2株生长一致的玉米植株,每个小区采集10株玉米,按不同部位分类采集样品,同一小区同一部位样品剪碎混合,-80 ℃低温保存备用。
1.2.3 Bt蛋白的测定 采用Cry1Ab/Ac酶联免疫定量检测试剂盒(上海佑隆公司,货号:AA0341)测定样品Bt蛋白含量,按照说明书操作并绘制标准曲线,样品吸光度值由酶标仪(bio-rad iMark microplate reader)读取,记录D450 nm吸光度。
1.3 转基因玉米室内抗虫性鉴定
亚洲玉米螟、黏虫、棉铃虫均由本研究室人工繁殖,处理组为转基因玉米双抗505-12-5,对照组为非转基因玉米中科玉505。玉米螟在6~8叶期、花丝期、籽粒期采集相应幼嫩部位饲喂;黏虫在4~6叶期,棉铃虫在花丝期采集相应幼嫩部位饲喂。饲喂容器均采用6孔细胞培养板,每孔10头初孵幼虫,每板为一个重复,4次重复。所有处理每24 h更换相同来源的新组织,并记录活虫数,按以下公式计算校正死亡率。
校正死亡率/%=(处理组死亡率-对照组死亡率)/(100-对照组死亡率)×100
1.4 田间抗虫性鉴定
1.4.1 亚洲玉米螟 在玉米心叶期(6~8叶期)和吐丝期人工接种初孵幼虫,选择傍晚接虫。心叶期接种到心叶内,吐丝期接种到花丝上。每小区人工定株接虫40株,每株接虫30~40头。间隔3 d后进行第2次接虫,最后一次接虫14 d后调查。
心叶期调查:调查接虫株中上部叶片被玉米螟危害程度,包括受害叶片上的虫孔大小、数量及分布等,计算平均食叶级别,根据平均食叶级别判定玉米心叶期对玉米螟的抗性水平。危害程度分级标准及抗性评价标准参照中华人民共和国农业部(2006)。
吐丝期调查:根据雌穗被害情况、蛀孔数量、隧道长度及存活幼虫龄期、数量等,计算各小区雌穗被害级别平均值,判定玉米穗期对玉米螟的抗性水平。雌穗被害级别分级标准及抗性评价标准参照中华人民共和国农业部(2007)。
1.4.2 黏虫 4~6叶期接虫,接虫方法及接虫次数同玉米螟。最后一次接虫后14 d调查玉米叶片被害程度及黏虫幼虫存活数,记录心叶被害级别并计算被害级别平均值,判定玉米心叶期对黏虫的抗性水平。被害程度分级标准及抗性评价标准参照中华人民共和国农业部(2007)。
1.4.3 棉铃虫 吐丝期接虫,接虫方法及接虫次数同玉米螟。最后一次接虫后14 d调查雌穗被害率、幼虫存活数、雌穗被害长度,确定雌穗被害级别并计算平均值,判定玉米对棉铃虫的抗性水平。雌穗被害级别分级标准及抗性评价标准参照中华人民共和国农业部(2007)。
2 结果与分析
2.1 转基因玉米双抗505-12-5中Bt蛋白的时空表达
2.1.1 转基因玉米双抗505-12-5根、茎及叶片中Bt蛋白的时空表达 由图1看出,根、茎、叶3种器官中,6~8叶期至吐丝期,Bt蛋白含量逐渐降低,乳熟期略有回升。6~8叶期,Bt蛋白含量最高,其中根部最高(517.3 ng·g-1),其次为心叶(453.8 ng·g-1),茎中表达量最低;大喇叭口期,根、茎、心叶中Bt蛋白含量均较6~8叶期降低,心叶中Bt蛋白含量最高(353.3 ng·g-1),根中Bt蛋白降幅最大,为281.3 ng·g-1,总体表现为心叶>根>茎;吐丝期,根、茎、心叶中Bt蛋白含量继续降低,仍然表现为心叶>根>茎;乳熟期,心叶Bt蛋白含量和吐丝期相比略有下降,为185.4 ng·g-1,根、茎中Bt蛋白上升,三者Bt蛋白含量相当,在160~190 ng·g-1之间,差异不显著。6~8叶期是亚洲玉米螟的第一次发生高峰,第一代幼虫均集中在心叶内取食,因此,6~8叶期Bt蛋白表达量高低是决定转基因玉米后期能否推广的关键。从研究结果看,6~8叶期心叶Bt蛋白表达量高达453.8 ng·g-1,对于防止亚洲玉米螟幼虫侵害具有重要作用。
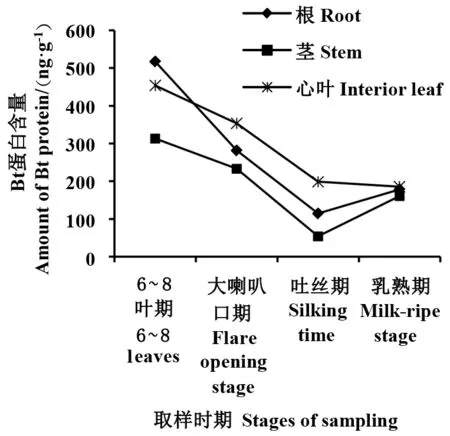
图1 转基因玉米双抗505-12-5不同生育期根、茎、叶片中Bt蛋白含量Fig.1 Bt protein content of double-resistant 505-12-5 transgenic maize in root, stem and leaf at different growth stages
2.1.2 转基因玉米双抗505-12-5中Bt蛋白在繁殖器官中的表达 由图2可以看出,转基因玉米双抗505-12-5繁殖器官中Bt蛋白含量比其他组织中含量低,不同繁殖器官Bt蛋白含量有差异,幼籽Bt蛋白含量最高(181.3 ng·g-1),其他依次是花丝、花粉、雄穗,分别为100.1、95.0、79.8 ng·g-1。从吐丝期繁殖器官Bt蛋白含量看,籽粒和花丝中Bt蛋白含量较高,有助于抵御亚洲玉米螟幼虫的侵害,可以有效减少入侵幼穗的亚洲玉米螟数量,从而保护幼穗,减少产量损失。

图2 转基因玉米双抗505-12-5繁殖器官中Bt蛋白含量Fig.2 Bt protein content of double-resistant 505-12-5 transgenic maize in reproductive organs
2.2 转基因玉米双抗505-12-5对3种鳞翅目害虫的田间抗虫性
双抗505-12-5对3种鳞翅目害虫田间抗性见
表1。感虫品种综31对3种鳞翅目害虫抗性均为感虫水平。对亚洲玉米螟来说,心叶期和雌穗期,双抗505-12-5的被害级别平均值分别为2.71、1.40,均表现为高抗,中科玉505均表现为感虫,而当地对照登海3号在心叶期表现为中抗,在雌穗期表现为感虫;对黏虫来说,心叶期,双抗505-12-5的平均食叶级值为1.97,抗性类型为高抗,中科玉505和登海3号分别为感虫和抗性级别;对棉铃虫来说,雌穗期,双抗505-12-5被害级别平均值为0.97,中科玉505和登海3号的被害级别平均值分别为5.97、5.67,均表现为感虫。
总体看,转基因玉米双抗505-12-5田间对亚洲玉米螟、黏虫、棉铃虫的抗性水平均达到高抗,抗性显著高于非转基因对照中科玉505及当地对照登海3号。

表1 转基因玉米双抗505-12-5对3种鳞翅目害虫的田间抗性Table 1 Field resistance of double-resistant 505-12-5 transgenic maize against three Lepidoptera pests
同列不同小写字母表示差异显著。S:感虫, HR:高抗, MR:中抗。
Small letters within a column represent significant differences among treatments atP<0.05. S: Susceptible; HR: High resistant; MR: Moderate resistant.
2.3 转基因玉米双抗505-12-5对3种鳞翅目害虫的室内抗虫性
6~8叶期,用双抗505-12-5幼嫩叶片饲喂亚洲玉米螟幼虫24 h后,校正死亡率为87.37%,48 h后达到100%;花丝期和乳熟期,接虫后24 h,亚洲玉米螟幼虫校正死亡率均达到100%。研究表明,转基因玉米双抗505-12-5在心叶期、花丝期和乳熟期均对亚洲玉米螟表现出明显的抗虫效果。
4~6叶期,双抗505-12-5对黏虫的抗性起效较慢,接虫后24 h,黏虫校正死亡率仅为10.17%,但接虫后48 h,校正死亡率达到98.21%,表明双抗505-12-5室内对黏虫同样具有较好的抗性。
花丝期,用双抗505-12-5花丝饲喂棉铃虫幼虫24 h后,棉铃虫幼虫校正死亡率达到80.18%,48 h后校正死亡率为92.45%,表明双抗505-12-5室内对棉铃虫也表现出较好的抗性。
3 讨论
转Bt基因玉米在美国、巴西等部分拉美国家已经得到大面积推广,转基因玉米占种植玉米总面积的80%以上,常见的转Bt基因玉米有MON810、Bt176、Bt11等。抗虫转基因玉米外源杀虫蛋白的表达量高低与玉米的杀虫效果有直接的关系。本试验中,心叶期根部Bt蛋白的含量最高,高于心叶等其他器官,但与心叶表达量相差仅为70 ng·g-1,这与其他报道心叶含量最高略有差异(姜志磊等,2008, 史晓利等,2009; 王建武等,2003)。随着植株生长,到大喇叭口期,各器官表达量尤其根部Bt蛋白的含量显著下降,心叶中的Bt蛋白表达量最高;吐丝期各器官Bt蛋白表达量持续降低,发育后期繁殖器官Bt蛋白表达量低于其他生育期各器官。总体来看,Bt蛋白表达量表现出随着生育期延长而逐渐降低的趋势,不同器官在不同生育期Bt蛋白的表达量也不同(王冬妍等,2004; 王建武等,2003),国外主要抗虫品种MON810 Bt蛋白时空表达的研究也显示了类似的结果(Székácsetal.,2010)。Bt蛋白的表达量除了与转基因调控元件有关外,与玉米品种、生长环境、播种时间也有关系。同一玉米品种播期不同,Bt蛋白表达量也有差异,如Bt11春玉米心叶比夏玉米心叶表达量低大约700 ng,而Bt-X春玉米心叶比夏玉米心叶表达量低约150 ng(王培等,2012)。另外,研究表明,Bt蛋白的表达量与温度有关,在一定范围内,温度低,蛋白表达量相对较低(路献勇等,2015)。
转基因抗虫玉米推广前,除了测定蛋白表达量,最关键的是明确抗虫效果。国外推广品种经田间及离体生测试验证实,均对靶标害虫具有显著的抗性(常雪等,2007; Burknessetal.,2001;Lynchetal.,1999; Williamsetal.,1998)。本试验中,转基因玉米双抗505-12-5的室内抗虫性鉴定结果表明,心叶期饲喂转基因玉米双抗505-12-5后48 h,亚洲玉米螟校正死亡率达100%。常雪等(2013)用转cry1Ab/cry2Aj基因玉米不同品系心叶饲喂亚洲玉米螟幼虫48 h的死亡率为60%~6.8%,96 h后大部分幼虫死亡,死亡时间较双抗505-12-5推迟;双抗505-12-5花丝期、乳熟期饲喂亚洲玉米螟幼虫校正死亡率24 h即达100%,常雪等(2013)发现亚洲玉米螟幼虫取食转cry1Ab/cry2Aj基因玉米各品系雄穗、苞叶、花丝和雌穗48 h后死亡率在90%左右;双抗505-12-5花丝期饲喂棉铃虫后48 h,校正死亡率为92.45%,而常雪等(2016)发现取食转3个转cry1Ab/cry2Aj玉米品系和1个转cry1Ab/vip3DA品系Bt玉米叶片96 h后棉铃虫死亡率为87.50%~90.00%,死亡时间较双抗505-12-5推迟48 h。双抗505-12-5心叶期饲喂黏虫,接虫后24 h校正死亡率仅为10.17%,但48 h后即达98.21%。转基因玉米双抗505-12-5室内对亚洲玉米螟、棉铃虫、黏虫杀虫效果更为快速,分析其原因可能是玉米材料不同,杀虫蛋白活性有差别,同时与试虫来源、饲养环境、饲养方法等有关。
转基因玉米双抗505-12-5田间接种3种试虫后,基本没有受害症状,在心叶期及花丝期对亚洲玉米螟的抗性、在心叶期对黏虫、花丝期对棉铃虫的抗性均达高抗水平,其平均级值与非转基因对照玉米、当地品种及感虫品种相比,差异显著。这与常雪等(2013)对国产转cry1Ab/cry2Aj基因玉米的研究一致,其抗虫效果不逊于国外常见转基因抗虫玉米(常雪等,2007; 常雪艳等,2006)。目前,国内报道的转基因玉米大多也表现出了对靶标害虫的良好抗性,如玉米螟对玉米Bt799的食叶级别为1级(武奉慈等,2014),转cry1Ah基因抗虫玉米HGK60对于亚洲玉米螟和棉铃虫有很强的杀虫活性,达到高抗级别,但对于黏虫为抗性级别(宋苗等,2016);BT-X高抗亚洲玉米螟(王培等,2012),但大部分品种只研究了对某一种害虫的抗性或仅研究了田间抗性,需进一步研究其综合抗性。
综合分析,转基因玉米双抗505-12-5对3种重要鳞翅目害虫在田间及室内均表现了显著的抗虫性,值得进一步研究和推广应用。
常雪, 常雪艳, 何康来, 王振营, 白树雄, 2007. 转cry1Ab基因玉米对黏虫的抗性评价. 植物保护学报, 34(3): 225-228.
常雪, 王伟, 沈志成, 叶恭银, 2016. 转cry1Ab/cry2Aj、cry1Ab/vip3DA玉米对棉铃虫、甜菜夜蛾和斜纹夜蛾的抗虫性评价. 植物保护学报, 43(6): 951-957.
常雪, 王伟, 沈志成, 叶恭银, 2013. 转cry1Ab/cry2Aj玉米对亚洲玉米螟的抗性评价. 植物保护学报, 40(4): 340-344.
常雪艳, 何康来, 王振营, 白树雄, 2006. 转Bt基因玉米对棉铃虫的抗性评价. 植物保护学报, 33(4): 374-378.
JAMES C, 2016. 015年全球生物技术/转基因作物商业化发展态势. 中国生物工程杂志, 36(4): 1-11.
姜伟丽, 马艳, 马小艳, 奚建平, 马亚杰, 2013. 转基因抗草甘膦抗虫棉对棉田冠层节肢动物群落的影响. 生物安全学报, 22(1): 51-56.
姜志磊, 刘德璞, 李晓辉, 孔祥梅, 李葱葱, 李飞武, 孙传波, 袁英, 2008. 转基因抗虫玉米Bt毒蛋白的时空表达分析. 吉林农业科学, 33(6): 35-37.
路献勇, 李淑英, 朱加保, 程福如, 2015. 昼夜温度变化对棉花叶片Bt蛋白表达及抗虫性的影响. 中国棉花, 42(8): 35-37, 40.
聂继东, 汪宏伟, 2017. 黏虫对玉米的危害与防治. 吉林农业 (8): 84.
马小艳, 马艳, 彭军, 姜伟丽, 马亚杰, 2013. 转基因抗草甘膦抗虫棉的生存竞争能力. 生物安全学报, 22(2): 103-108.
史晓利, 杨益众, 蔡建华, 张小丽, 施敏娟, 2009. Bt毒蛋白在转基因抗虫玉米中的表达及在亚洲玉米螟中的转移积累. 应用生态学报, 20(11): 2773-2777.
宋苗, 汪海, 张杰, 何康来, 梁革梅, 朱莉, 黄大昉, 郎志宏, 2016. 转Btcry1Ah基因抗虫玉米对亚洲玉米螟、棉铃虫和黏虫的抗性评价. 生物技术通报, 32(6): 69-75.
王冬妍, 王振营, 何康来, 丛斌, 白树雄, 文丽萍, 2004.Bt玉米杀虫蛋白含量的时空表达及对亚洲玉米螟的杀虫效果. 中国农业科学, 7(8): 1155-1159.
王红军, 郭书亚, 2017. 玉米螟对夏玉米果穗的危害及产量的影响. 江苏农业科学, 45(8): 95-97.
王建武, 冯远娇, 骆世明, 2003.Bt玉米抗虫蛋白表达的时空动态及其土壤降解研究. 中国农业科学, 36(11): 1279-1286.
王培, 何康来, 王振营, 王应伦, 2012. 转cry1Ac玉米对亚洲玉米螟的抗性评价. 植物保护学报, 39(5): 395-400.
王延锋, 2010. 转Bt基因抗虫玉米田间试验与遗传稳定性分析. 博士学位论文. 哈尔滨: 东北农业大学.
王振营, 鲁新, 2000. 我国研究亚洲玉米螟历史、现状与展望. 沈阳农业大学学报, 31(5): 402-412.
武奉慈, 刘金文, 刘娜, 曲文利, 张明, 宋新元, 2014. 转基因抗虫玉米对亚洲玉米螟的抗性评价. 玉米科学, 22(6): 148-150.
岳同卿, 郎志宏, 王延锋, 张杰, 何康来, 黄大昉, 2010. 转Btcry1Ah基因抗虫玉米的获得及其遗传稳定性分析. 农业生物技术学报, 18(4): 638-644.
中华人民共和国农业部. 2006. 玉米抗病虫性鉴定技术规范 第5部分: 玉米抗玉米螟鉴定技术规范: NY/T 1248.5-2006. 北京: 中华人民共和国农业部.
中华人民共和国农业部. 2007. 转基因玉米及其环境安全性检测 抗虫玉米 第1部分: 抗虫性: 农业部953号公告-10.1-2007. 北京: 中华人民共和国农业部.
BURKNESS E C, HUTCHISONW D, BOLIN P C, BARTELS D W, WARNOCK D F, DAVIS D W, 2001. Field efficacy of sweet corn hybrids expressing aBacillusthuringiensistoxin for management ofOstrinianubilalis(Lepidoptera: Crambidae) andHelicoverpazea(Lepidoptera: Noctuidae).JournalofEconomicEntomology, 94(1): 197-203 .
LYNCH R E, WISEM AN B R, PLAISTED D, WARNICK D,1999. Evaluation o f transgenic sweet corn hybrids expressingCryIA(b) toxin for resistance to corn ear worm and fall armyworm (Lepidoptera: Noctuidae ).JournalofEconomicEntomology, 92(1): 246-252 .
WILLIAMS W P, BUCKLEY P M, SAGERS J B, 1998. Evaluation of transgenic corn for resistance to corn earworm (Lepidoptera: Noctuidae),fall armyworm (Lepidoptera: Noctuidae), and southwestern corn borer (Lepidoptera: Crambidae) in a laboratory bioassay.JournalofAgriculturalEntomology, 15(2): 105-112.
