壳聚糖纳米颗粒包裹猪白细胞介素2和融合白细胞介素4/6基因对猪圆环病毒2型疫苗免疫的强化
2018-03-26陈祎宋婷玉李金海肖永乐曾光志万小平杨璐一方鹏飞王泽洲高荣
陈祎, 宋婷玉, 李金海, 肖永乐, 曾光志, 万小平,杨璐一, 方鹏飞*, 王泽洲, 高荣*
1. 四川大学生命科学学院,生物资源与生态环境教育部重点实验室,动物疾病防控和食品安全四川省重点实验室,成都610065;2. 四川省华派生物制药有限公司,成都610026; 3. 四川省动物疫病预防控制中心,成都610035)
细胞因子因其安全性和高效性而成为一种理想的免疫佐剂,例如γ-干扰素(IFN-γ)(Wangetal.,2013)、白细胞介素2(IL-2)(Yangetal.,2010)、白细胞介素4(IL-4)(Zhangetal.,2007)和白细胞介素6(IL-6)(Lietal.,2011)。IL-2、IL-4和IL-6在细胞和体液免疫反应中起重要作用。IL-2具有多种生物学功能,包括促进T细胞增殖和增强NK细胞的细胞毒性,刺激活化的B淋巴细胞增殖,并诱导免疫球蛋白分泌等(Collins & Oldham,1993)。IL-4可影响体液免疫和细胞免疫应答,如免疫球蛋白的产生、类别转换和分泌(Pasquinietal.,1997)。IL-6具有促进白细胞介素基因表达、B细胞分化、T细胞活化的作用(Kishimoto,2010)。
猪IL-2或IL-6与CpG免疫刺激序列的混合基因能有效提高动物对病原菌的抵抗力(Yangetal.,2010;Huangetal.,2013);此外,猪IL-4/6融合基因与单独的IL-4和IL-6相比,能诱导机体产生更强的免疫应答;猪IL-4/6融合基因能加强大肠杆菌疫苗、猪肺炎支原体疫苗、猪繁殖与呼吸综合征(PRRS)疫苗对机体的体液和细胞免疫,提高接种动物的免疫保护性(Zhangetal.,2007;Zhangetal.,2012;Yangetal.,2013);猪IL-2、IL-4、IL-6的融合基因(IL-4/6-2)相比于IL-4/6和IL-2在小鼠Musmusculus上可产生更强的免疫协同调节作用(杨璐一等,2014)。
因此,本实验在前期研究的基础上,为研制新型高性价比的PCV-2疫苗免疫调节剂,评估共表达IL-2和IL-4/6融合基因纳米颗粒对仔猪生长和PCV-2免疫应答的作用。
1 材料与方法
1.1 材料
质粒:真核表达载体VR1020-猪融合白细胞介素4/6-2(VRIL-4/6-2),由本实验室构建并保存;壳聚糖(CS):15 kD,脱乙酰度95%以上,购自Sigma Aldrich;多聚磷酸钠(TPP):购自Sigma(USA);鲎试剂:购自湛江A & C公司;疫苗:PCV-2灭活疫苗(ZJ/C株)(圆环康)和实验动物:健康21日龄长白、约克夏和杜洛克杂交仔猪均由四川省华派生物制药有限公司提供。
1.2 方法
1.2.1VRIL-4/6-2真核表达质粒的大量制备及内毒素检测参照《分子克隆指南 第三版》(Josephetal.,2002)制备质粒,溶解于TE缓冲液,质粒浓度和纯度使用紫外分光光度计检测;质粒内毒素含量用鲎试剂检测。
1.2.2VRIL-4/6-2质粒壳聚糖纳米颗粒的制备离子交联法(Bodmeieretal.,1989)制备VRIL-4/6-2质粒壳聚糖纳米颗粒。将壳聚糖溶解于1%的冰醋酸溶液(pH5.5),配制为2.4 mg·mL-1的溶液;ddH2O配制10 mg·mL-1的磷酸三苯酯(TPP)溶液;以上溶液均用0.22 μm微孔滤膜过滤除菌。将VRIL-4/6-2质粒与适量TPP溶液混匀,55 ℃孵育20 min;壳聚糖与质粒质量比为30∶1,在50~55 ℃水浴磁力搅拌下将质粒与TPP的预混液缓慢滴加至壳聚糖溶液中,混合均匀,恒温孵育10 min备用,记作VRIL-4/6-2-CS。用Zetasizer3000HS/IHPL粒度仪检测纳米颗粒粒径和电位。
1.2.3动物实验10头21日龄长白、约克夏和杜洛克杂交仔猪,经ELISA和荧光定量PCR(qPCR)检测PCV-2、PRRS病毒、猪瘟病毒和支原体均为阴性。随机分成实验组和对照组,每组5头。实验组颈部肌肉注射2.5 mL VRIL-4/6-2-CS(0.5 mg·mL-1),对照组注射相同剂量生理盐水;2组均颈部肌肉注射2.5 mL PCV-2疫苗。猪免疫接种前记为第0天,接种后第7、14、28天定时采集前腔静脉血用于检测免疫反应。接种前及实验结束第28天时分别测量2组仔猪的体质量,用于评价实验质粒对仔猪生长的影响。
观察组48例中,痊愈15例,显效20例,有效10例,无效3例,总有效率为93.75%;对照组48例中,痊愈11例,显效15例,有效12例,无效10例,总有效率为79.16%,观察组明显优于对照组,两组效果比较,差异有统计学意义(P<0.05)。
1.2.4PCV-2特异性抗体的测定取500 μL抗凝血低速离心后收集血清,参照ELISA试剂盒说明书操作,在酶标仪上测定OD630值,检测血清中PCV-Ab的含量,试剂盒购自武汉科前生物公司。
1.2.5IgG1和IgG2a的测定取500 μL抗凝血低速离心后收集血清,参照ELISA 试剂盒说明书操作,在酶标仪上测定OD450值,检测血清中IgG1和IgG2a的含量,试剂盒购自成都敏鑫科生物科技公司。
1.2.6CD4+和CD8+T淋巴细胞的测定取100 μL新鲜抗凝血摇匀,加入2 μL anti-porcine CD3-SPRD、2 μL anti-porcine CD4-FITC、2 μL anti-porcine CD8-PE,震荡混匀避光孵育30 min;加入600 μL红细胞裂解液,避光裂解10 min,500 r·min-1离心5 min,去上清;加入1 mL 磷酸缓冲盐溶液(PBS),轻轻悬浮细胞,500 r·min-1离心5 min,去上清;重复洗涤1次,细胞沉淀加入300 μL PBS混匀,悬浮细胞,上机检测。
1.2.7荧光定量检测基因表达水平100 μL抗凝血加入1 mL的RNAiso pius,充分裂解后提取细胞总RNA,用TransScript All-in-One First-Strand cDNA Synthesis SuperMix for Qpcr (One-Step gDNA Removal)反转录试剂盒合成cDNA。根据GenBank中的猪PPIA、TLR-2、TLR-7、IL-2、IL-4、IL-6、TNF-α、STAT-1、STAT-2、STAT-3基因的cDNA序列,分别设计合成其特异性扩增引物(表1)。
以仔猪不同时期血液cDNA为模板,表1中设计的引物进行扩增。用Bio-Rad IQ5荧光定量PCR仪检测不同基因相对表达的情况,15 μL体系,用SsoAdvanceTMUniversal SYBR Green Supermix进行荧光定量分析。扩增参数为:95 ℃ 预变性30 s;95 ℃变性 5 s,最佳退火温度退火30 s,40个循环。熔解曲线参数:65~95 ℃,每6 s上升0.5 ℃。PPIA为内参基因,采用2-ΔΔCT法分析实时荧光PCR数据,比较同一目的基因不同时期的相对表达水平差异。
1.2.8数据分析以上各数据用GraphPad Prism 6中的双因素方差分析和单因素方差分析进行差异比较,P<0.05表示差异有统计学意义。

表1 荧光定量PCR特异性引物Table 1 The primers for qPCR
2 结果
2.1 仔猪体质量变化
实验开始及结束时分别称量每只仔猪的体质量,分别计算每组仔猪体质量的平均值和方差,仔猪体质量变化见表2。结果显示,实验组仔猪体质量增加显著高于对照组(P<0.05)。
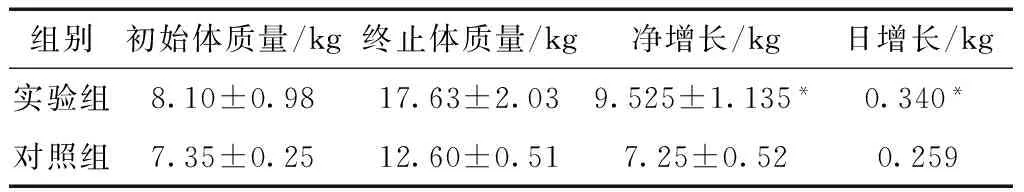
表2 实验28天仔猪体质量变化Table 2 Body mass change of piglets during 28 days
注 Note:*P<0.05
2.2 仔猪血清中特异性抗体含量
在接种第7天后,实验组和对照组中均可以检测到PCV-2抗体,但是2组含量之间的差异无统计学意义(P>0.05)(图1)。
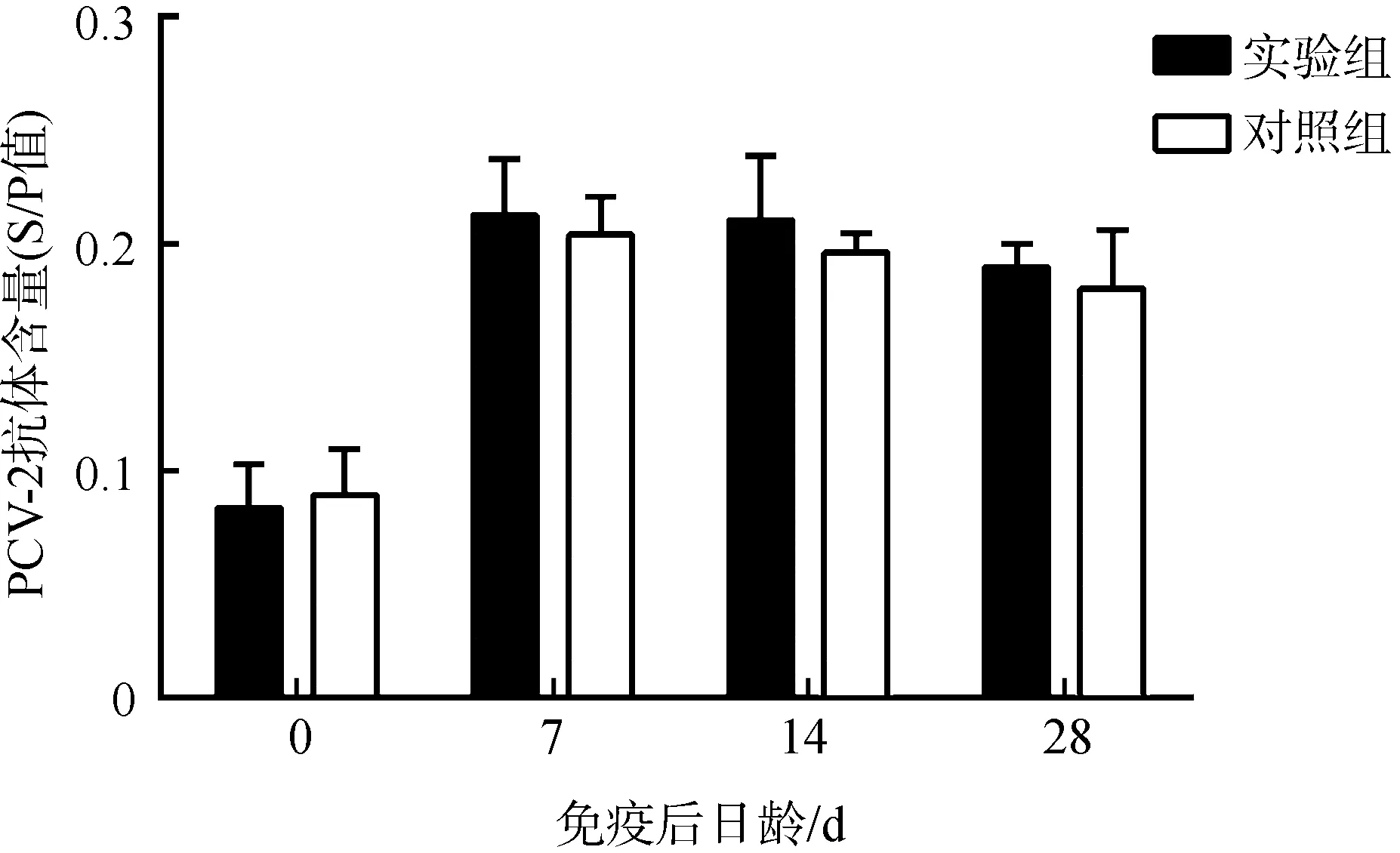
图1 仔猪血清中PCV-2抗体含量的变化Fig. 1 PCV-2 antibody titers in the serum of the experimental piglets
S/P=(S-N)/(P-N)=(样品OD值-阴性对照OD值)/(阳性对照OD值-阴性对照OD值), 当S/P≥0.16时, 认为PCV-2抗体检测为阳性
S/P=(S-N)/(P-N)=(ODsample-ODnegative control)/(ODpositive control-ODnegative control), titer of PCV-2 antibody was considered positive whenS/P≥0.16
2.3 仔猪血清中IgG1 和IgG2a 含量
实验组仔猪血清中IgG1的含量显著低于对照组(P<0.05),而IgG2a的含量显著高于对照组(P<0.05)(图2)。
2.4 CD4+和CD8+T淋巴细胞亚群
流式细胞术检测仔猪血液中CD4+和CD8+T淋巴细胞亚群的数量变化,实验组仔猪血液中的CD4+T和CD8+T淋巴细胞的数量与对照组相比均显著增加(P<0.05)(图3)。
2.5 免疫相关基因表达量分析
2.5.1Toll-like受体基因表达定量分析实验组TLR-2基因的表达水平只在免疫后第14天显著高于对照组(P<0.05),而实验组TLR-7基因的表达水平在免疫后第28天显著高于对照组(P<0.05)(图4)。
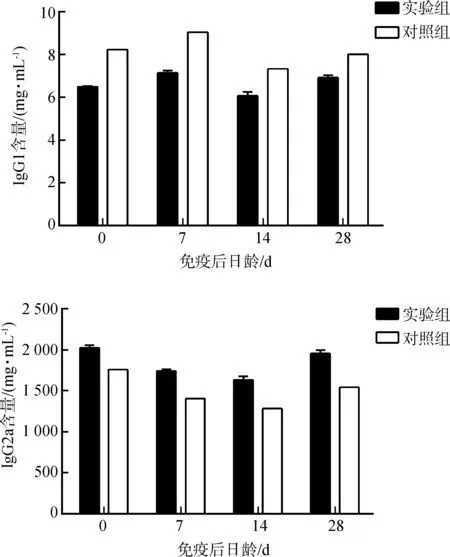
图2 仔猪血清中IgG1和IgG2a含量的变化Fig. 2 The change of the levels of IgG1 and IgG2a in the serum of piglets
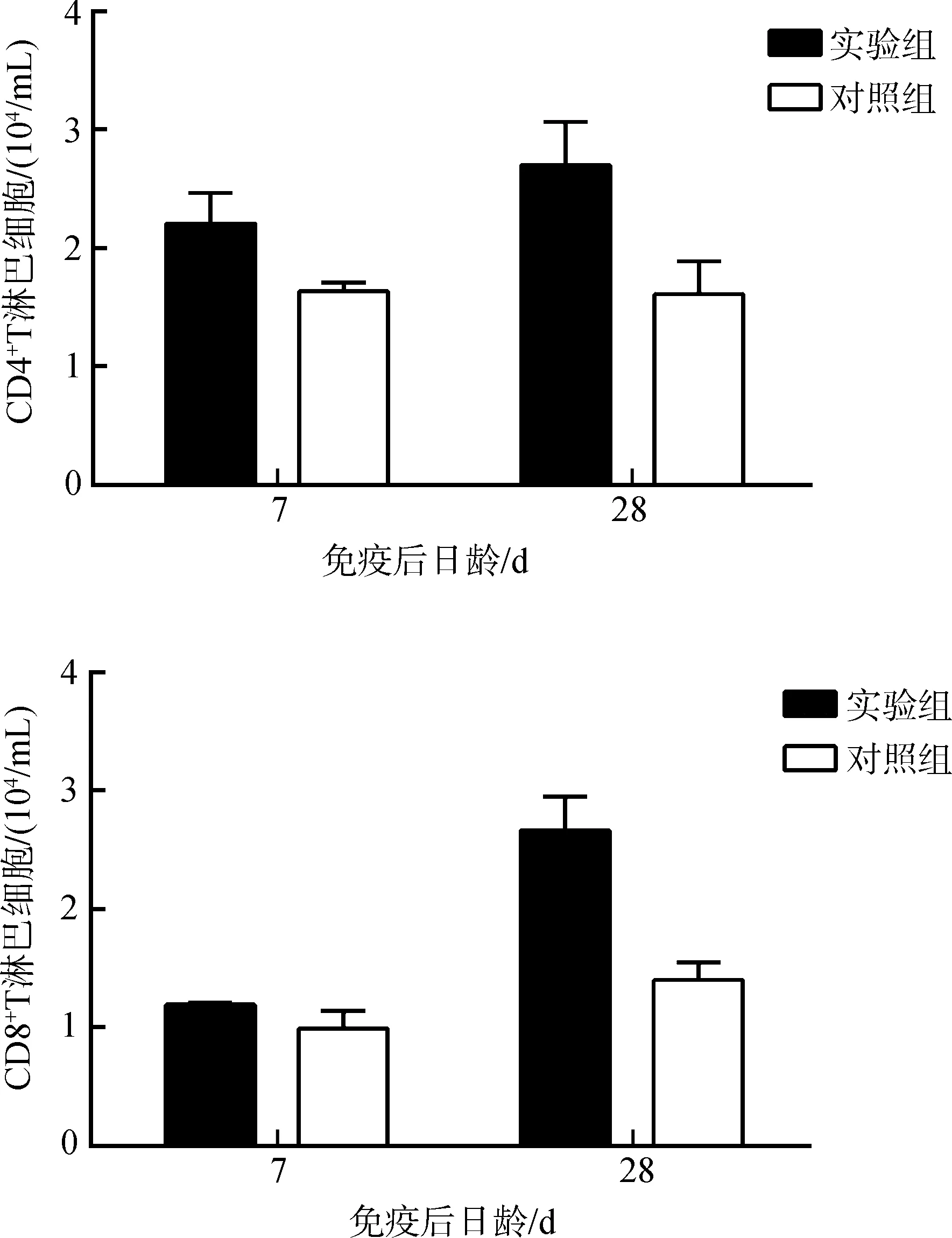
图3 仔猪血液中 CD4+和CD8+T淋巴细胞数量的变化Fig. 3 CD4+ and CD8+ T cell numbers in the blood of experimental piglets
2.5.2细胞因子基因表达定量分析实验组仔猪血液中IL-2、IL-4、IL-6和TNF-α基因的表达水平在免疫后第28天均显著高于对照组(P<0.05),其中,IL-4和IL-6基因表达水平分别在第7天和第14天显著高于对照组(P<0.05)(图5)。
2.5.3免疫信号传导分子基因表达定量分析STAT-1基因表达水平在免疫后第7~14天均显著高于对照组(P<0.05),STAT-2基因表达水平在免疫后第14~28天显著高于对照组(P<0.05),STAT-3基因表达水平在免疫后第7、28天显著高于对照组(P<0.05)(图6)。
3 讨论
细胞因子是调节动物免疫功能的关键分子。许多细胞因子被证实作为佐剂对体液或细胞免疫有增强作用(Pasquinietal.,1997;Kayamuroetal.,2010)。IL-2不仅支持T细胞和NK细胞的活化和增殖,还可以刺激和活化B淋巴细胞增殖和诱导免疫球蛋白分泌(Smith,1988;Collins & Oldham,1993)。IL-4可促进体液免疫,增加特异性和非特异性杀伤功能(Erbetal.,1997;Paul,2015)。IL-6可调节B细胞活化、抗体产生和Th1型免疫反应(Paul & Seder,1994;Kishimoto,2006)。有研究表明,猪IL-4/6、IL-2和IL-4/6-2融合基因均可安全增强动物系统和全面的免疫(Yangetal.,2010,2013;杨璐一等,2014)。基于以前的研究,本研究第一次将猪IL-4/6-2融合基因应用到猪上,来加强对PCV-2疫苗的免疫效果。
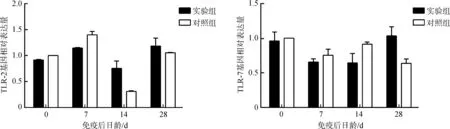
图4 仔猪TLR基因表达水平的变化Fig. 4 The change of expression levels of TLR gene in the blood of experimental piglets
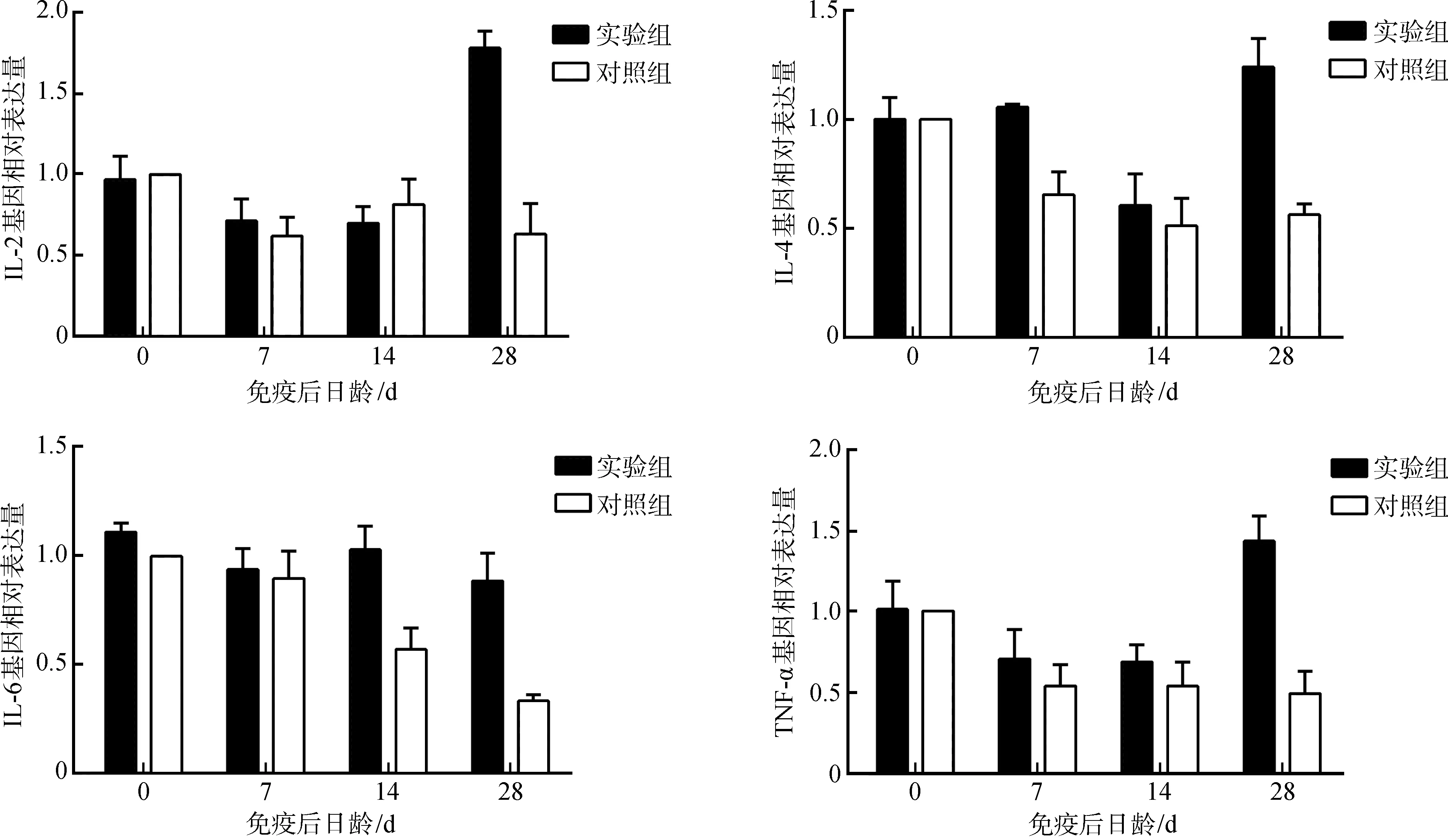
图5 仔猪细胞因子基因表达水平的变化Fig. 5 The change of cytokine gene expression in the blood of experimental piglets
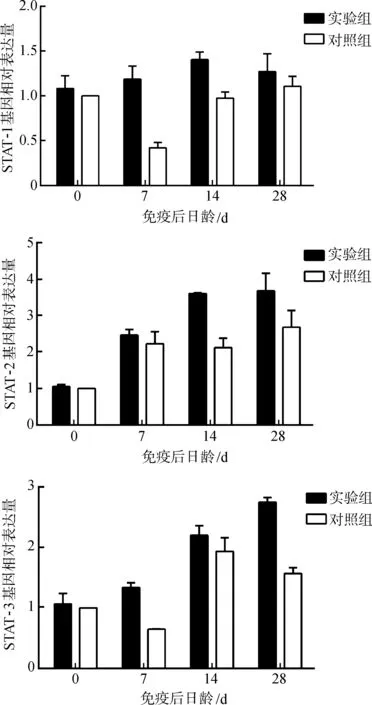
图6 仔猪免疫信号传导分子基因表达水平的变化Fig. 6 The change of the immune signal transduction molecules related genes in the blood of experimental piglets
本实验用壳聚糖将重组质粒VRIL-4/6-2包裹,制备壳聚糖纳米颗粒,与PCV-2疫苗同时对实验仔猪进行肌肉注射免疫接种。实验所用重组质粒通过2A短肽连接3种白细胞介素基因,构建共表达质粒。2A短肽具有的自剪切机制使连接前后的基因表达量相同(de Felipe & Ryan,2004;Szymczak & Vignali,2005)。裸露的基因不稳定且半衰期短,转染效率低;而壳聚糖包裹质粒可形成小的带正电荷的纳米颗粒,与裸露的质粒相比,它可以更有效地携带目的基因进入细胞,还可以抑制其在生物体内降解,被证实是一种有潜力的缓释材料,并且还具有生物相容性(Chewetal.,2003;Cui & Mumper,2003),因此,壳聚糖作为基因载体的研究发展迅速(Buschmannetal.,2013;Fernándezetal.,2016)。Huang等(2005)通过荧光标记研究壳聚糖-DNA复合粒子进入细胞的过程,证实了壳聚糖-DNA复合粒子可以被细胞内吞,继而进行后续基因转染。之前的研究也表明,壳聚糖纳米粒子在动物身上用于基因传递几乎没有毒性和影响(Yangetal.,2010;Zhangetal.,2012),并且空白载体质粒组与空白组没有差异(Yangetal.,2010)。本次实验由于阴性动物难于筛选获得,数量有限,并未进行空白载体质粒组的实验。
在实验期间,实验组和对照组仔猪注射局部均未出现病变、损伤或其他系统性症状。实验开始和结束时分别记录了每只仔猪的体质量,结果显示,实验组仔猪的生长速率显著高于对照组。有研究显示,IL-2、IL-4、IL-6、IL-10、TNF-α等细胞因子可在猪小肠中持续表达,并且细胞因子除了直接的免疫调节作用,还可影响上皮细胞的生长、内环境稳态和免疫细胞的运输(Oswald,2006;Devriendtetal.,2010)。推测VRIL-4/6-2质粒对调节仔猪内分泌、促进物质代谢有积极作用。
PCV-2特异性抗体和PCV-2中和抗体是体液免疫反应的重要检测指标(Meertsetal.,2006;Fortetal.,2007)。研究结果表明,实验组和对照组仔猪血清中PCV-2抗体的含量差异无统计学意义,且实验组仔猪血清中IgG1的含量显著低于对照组,这表明IL-4/6-2可能没有明显增强仔猪对PCV-2疫苗的体液免疫。此外,有研究显示,诱导细胞免疫对于PCV-2的防控很有必要(Fortetal.,2009)。本研究结果表明,实验组的CD4+和CD8+T淋巴细胞数量显著高于对照组,并且实验组的IL-2、TNF-α基因表达水平在第28天显著高于对照组。IL-2、IFN-γ和TNF-α均由Th1细胞产生,Th1细胞可诱导巨噬细胞活化、迟发性超敏反应和IgG2a的产生(Mosmann & Coffman,1989;Abbasetal.,1996)。同时,本次实验发现,接种质粒的实验组比对照组产生了更多的IgG2a抗体,表明实验组仔猪产生了更强的Th1型免疫应答,相比对照组,VRIL-4/6-2-CS提高了仔猪对PCV-2疫苗的细胞免疫应答。
另外,实验组仔猪血液中IL-4、IL-6、TLR-2、TLR-7、STAT-1、STAT-2、STAT-3基因的表达水平在不同时间段内也显著高于对照组。TLRs是先天免疫系统的重要组成部分,Toll-like受体在先天免疫中识别不同病原菌的保守分子模式,并参与对病原体的特异性体液和细胞免疫反应的激活(Kawai & Akira,2010)。STAT-1、STAT-2参与IFNs应答反应,并在干扰素抗病毒应答调节中起重要作用(Mitchell & John,2005;Steen & Gamero,2013);STAT-3可由多种细胞因子激活,例如IL-6、IL-12具有有效的抗炎作用,可调节细胞生长、凋亡、炎性基因的转录等重要细胞过程(Egwuagu,2009);JAK激酶和STAT蛋白参与了许多细胞因子的信号转导,它在细胞因子介导的免疫反应和调控中具有重要作用(Shuai & Liu,2003)。综上,这些基因表达水平的提高表明VRIL-4/6-2-CS诱导了更加全面的免疫应答,增强了仔猪的先天性和适应性免疫应答。
总之,本研究结果首次证实VRIL-4/6-2-CS可有效协同增强仔猪对PCV-2疫苗的免疫应答。VRIL-4/6-2-CS是一种新型高性价比佐剂,可提高猪对抗PCV-2感染的免疫预防能力,具有重要应用前景。
Joseph S, David W, 黄培堂, 等. 2002. 分子克隆实验指南(第三版)[M]. 北京: 科学出版社: 32-36.
杨璐一, 肖永乐, 万小平, 等. 2014. 猪白细胞介素2与融合白细胞介素4/6基因共表达的免疫效应研究[J]. 四川动物, 33(2): 167-173.
Abbas AK, Murphy KM, Sher A. 1996. Functional diversity of helper T lymphocytes[J]. Nature, 383(6603): 787.
Allan G, McNeilly F, Ellis J,etal. 2004. PMWS: experimental model and co-infections[J]. Veterinary Microbiology, 98(2): 165-168.
Beach NM, Meng XJ. 2012. Efficacy and future prospects of commercially available and experimental vaccines against porcine circovirus type 2 (PCV2)[J]. Virus Research, 164(1): 33-42.
Bodmeier R, Chen H, Paeratakul O. 1989. A novel approach to the oral delivery of micro- or nanoparticles[J]. Pharmaceutical Research, 6(5): 413-417.
Buschmann MD, Merzouki A, Lavertu M,etal. 2013. Chitosans for delivery of nucleic acids[J]. Advanced Drug Delivery Reviews, 65(9): 1234-1270.
Chae C. 2005. A review of porcine circovirus 2-associated syndromes and diseases[J]. The Veterinary Journal, 169(3): 326-336.
Chew JL, Wolfowicz CB, Mao HQ,etal. 2003. Chitosan nanoparticles containing plasmid DNA encoding house dust mite allergen, Der p 1 for oral vaccination in mice[J]. Vaccine, 21(21): 2720-2729.
Collins R, Oldham G. 1993. Recombinant human interleukin 2 induces proliferation and immunoglobulin secretion by bovine B-cells: tissue differences and preferential enhancement of immunoglobulin A[J]. Veterinary Immunology and Immunopathology, 36(1): 31-43.
Cui Z, Mumper RJ. 2003. Microparticles and nanoparticles as delivery systems for DNA vaccines[J]. Critical Reviews in Therapeutic Drug Carrier Systems, 20(2-3): 103-137.
de Felipe P, Ryan MD. 2004. Targeting of proteins derived from self-processing polyproteins containing multiple signal sequences[J]. Traffic, 5(8): 616-626.
Devriendt B, Stuyven E, Verdonck F,etal. 2010. EnterotoxigenicEscherichiacoli(K88) induce proinflammatory responses in porcine intestinal epithelial cells[J]. Developmental & Comparative Immunology, 34(11): 1175-1182.
Egwuagu CE. 2009. STAT3 in CD4+T helper cell differentiation and inflammatory diseases[J]. Cytokine, 47(3): 149-156.
Erb KJ, Rüger B, Von Brevern M,etal. 1997. Constitutive expression of interleukin (IL)-4invivocauses autoimmune-type disorders in mice[J]. Journal of Experimental Medicine, 185(2): 329-340.
Fernández EF, Santos-Carballal B, Weber WM,etal. 2016. Chitosan as a non-viral co-transfection system in a cystic fibrosis cell line[J]. International Journal of Pharmaceutics, 502(1): 1-9.
Fort M, Fernandes LT, Nofrarias M,etal. 2009. Development of cell-mediated immunity to porcine circovirus type 2 (PCV2) in caesarean-derived, colostrum-deprived piglets[J]. Veterinary Immunology and Immunopathology, 129(1): 101-107.
Fort M, Olvera A, Sibila M,etal. 2007. Detection of neutralizing antibodies in postweaning multisystemic wasting syndrome (PMWS)-affected and non-PMWS-affected pigs[J]. Veterinary Microbiology, 125(3): 244-255.
Ge X, Wang F, Guo X,etal. 2012. Porcine circovirus type 2 and its associated diseases in China[J]. Virus Research, 164(1): 100-106.
Gillespie J, Opriessnig T, Meng X,etal. 2009. Porcine circovirus type 2 and porcine circovirus-associated disease[J]. Journal of Veterinary Internal Medicine, 23(6): 1151-1163.
Huang J, Chen JL, Wang Y,etal. 2013. Enhancement of the immunity of piglets to pseudorabies vaccine with plasmids containing interleukin-6 gene and CpG motifs encapsulated in chitosan nanoparticles[J]. Procedia in Vaccinology, 7: 8-14.
Huang M, Fong CW, Khor E,etal. 2005. Transfection efficiency of chitosan vectors: effect of polymer molecular weight and degree of deacetylation[J]. Journal of Controlled Release, 106(3): 391-406.
Kawai T, Akira S. 2010. The role of pattern-recognition receptors in innate immunity: update on toll-like receptors[J]. Nature Immunology, 11(5): 373-384.
Kayamuro H, Yoshioka Y, Abe Y,etal. 2010. Interleukin-1 family cytokines as mucosal vaccine adjuvants for induction of protective immunity against influenza virus[J]. Journal of Virology, 84(24): 12703-12712.
Kishimoto T. 2006. Interleukin-6: discovery of a pleiotropic cytokine[J]. Arthritis Research & Therapy, 8(2): S2.
Kishimoto T. 2010. IL-6: from its discovery to clinical applications[J]. International Immunology, 22(5): 347-352.
Li D, Chen JL, Zhang H,etal. 2011. Improvement of the immunity of pig to hog cholera vaccine by recombinant plasmid with porcine interleukin-6 gene and CpG motifs[J]. Vaccine, 29(22): 3888-3894.
Meerts P, Misinzo G, Lefebvre D,etal. 2006. Correlation between the presence of neutralizing antibodies against porcine circovirus 2 (PCV2) and protection against replication of the virus and development of PCV2-associated disease[J]. BMC Veterinary Research, 2(1): 6.
Mitchell TJ, John S. 2005. Signal transducer and activator of transcription (STAT) signalling and T-cell lymphomas[J]. Immunology, 114(3): 301-312.
Mosmann TR, Coffman RL. 1989. Heterogeneity of cytokine secretion patterns and functions of helper T cells[J]. Advances in Immunology, 46: 111-147.
Oswald IP. 2006. Role of intestinal epithelial cells in the innate immune defence of the pig intestine[J]. Veterinary Research, 37(3): 359-368.
Pasquini S, Xiang Z, Wang Y,etal. 1997. Cytokines and costimulatory molecules as genetic adjuvants[J]. Immunology & Cell Biology, 75(4): 397-401.
Paul WE, Seder RA. 1994. Lymphocyte responses and cytokines[J]. Cell, 76(2): 241-251.
Paul WE. 2015. History of interleukin-4[J]. Cytokine, 75(1): 3-7.
Segalés J, Allan GM, Domingo M. 2005. Porcine circovirus diseases[J]. Animal Health Research Reviews, 6(2): 119-142.
Shuai K, Liu B. 2003. Regulation of JAK-STAT signalling in the immune system[J]. Nature Reviews Immunology, 3(11): 900-911.
Smith KA. 1988. Interleukin-2: inception, impact, and implications[J]. Science, 240(4856): 1169.
Steen HC, Gamero AM. 2013. STAT2 phosphorylation and signaling[J]. Jak-Stat, 2(4): e25790. DOI: 10.4161/jkst.25790.
Szymczak AL, Vignali DA. 2005. Development of 2A peptide-based strategies in the design of multicistronic vectors[J]. Expert Opinion on Biological Therapy, 5(5): 627-638.
Wang YP, Liu D, Guo LJ,etal. 2013. Enhanced protective immune response to PCV2 subunit vaccine by co-administration of recombinant porcine IFN-γ in mice[J]. Vaccine, 31(5): 833-838.
Yang X, Sun WK, Chen WL,etal. 2010. Promotion of the immunity of piglets to hog cholera vaccine induced by shuffled pig interleukin-2 gene and CpG immunostimulatory sequences encapsulated in chitosan nanoparticles[J]. Procedia in Vaccinology, 2(1): 51-59.
Yang X, Xiao YL, Chen JL,etal. 2013. Potentiation of immunity of piglets to mycoplasma hyopneumoniae vaccine with fused gene for pig IL-4 and IL-6 embodied in chitosan nanoparticles[J]. Procedia in Vaccinology, 7: 15-22.
Zhang H, Cheng C, Zheng M,etal. 2007. Enhancement of immunity to anEscherichiacolivaccine in mice orally inoculated with a fusion gene encoding porcine interleukin 4 and 6[J]. Vaccine, 25(41): 7094-7101.
Zhang HB, Wan XP, Bai GM,etal. 2012. Improvement of the immunity of piglets to PRRS vaccine by a porcine IL-4 and IL-6 fusion gene encapsulated in chitosan nanoparticles[J]. Procedia in Vaccinology, 6: 113-124.
