黄土高原两种针叶树种不同器官水碳氮磷分配格局及其生态化学计量特征
2018-03-22陈美玲崔君滕王凯博
陈美玲,崔君滕,邓 蕾,王凯博
1. 青岛林业投资开发有限公司,青岛266061 2. 中国科学院地球环境研究所 黄土与第四纪地质国家重点实验室,西安 710061 3. 中国科学院水利部水土保持研究所 黄土高原土壤侵蚀与旱地农业国家重点实验室,杨凌 712100
生态化学计量学是一门结合了生物学、化学和物理学等基本原理的新兴交叉学科,是研究生物系统能量平衡和多重化学元素(碳、氮、磷等)平衡的科学(Sterner and Elser,2000;曾德慧和陈广生,2005)。当前关于生态化学计量学的研究涉及到生态学研究的各个领域,其在生物体营养动态(Agren,2008)、消费者驱动的养分循环(Frost et al,2005;Wagner et al,2013)、 限 制性养分判别(Elser et al,2000;Gusewel,2004)以及全球及区域碳、氮、磷等元素生物地球化学循环(Reich and Oleksyn,2004;He et al,2006)方面具有重要应用,是当前生态学研究的前沿领域之一(贺金生和韩兴国,2010)。植物体不同器官具有不同的结构物质和功能属性,同一植物不同器官的水、碳、氮、磷等含量不尽相同(刘超等,2012)。对同种植物不同器官水、碳、氮、磷等元素含量及其生态化学计量特征的研究对揭示物种的生态策略和环境适应性具有重要的生态学和植物生理学意义(贺合亮等,2017)。
油松(Pinus tabulaeformis)是黄土高原地区分布最广的常绿针叶树种,有良好的保持水土、涵养水源及改良土壤的作用,也是暖温带湿润半湿润气候区的地带性植被(焦醒和刘广全,2009)。柴松(Pinus tabulaef i rmis f. shekanensis)为形似油松的一种松属植物,除具有良好的抗逆性和适应性等特点外,与油松相比,柴松还具有生长快、林分蓄积生长量高、树干通直和天然整枝良好等优点,是黄土高原上的珍贵乡土树种基因库,其在改善当地生态环境中具有举足轻重的作用(王凯博和上官周平,2006)。当前,针对黄土高原油松和柴松林种群动态(王凯博和上官周平,2006;焦醒和刘广全,2009;韩文娟等,2012)、群落结构(王梅和张文辉,2009;郝江勃等,2010)、根系分布(安慧等,2007;韦兰英和上官周平,2007)、碳库特征(杨晓梅等,2010;申家朋等,2013;孙美美等,2017)等方面已开展大量研究工作,然而,针对黄土高原油松和柴松不同器官之间水、碳、氮和磷含量的分配格局及其化学计量特征的研究鲜见报道。因此,本文以黄土高原两种典型乡土针叶树种油松和柴松为研究对象,研究两种树种水、碳、氮、磷在不同器官的分配格局,明确两种树种不同器官水、碳、氮、磷的生态化学计量特征,揭示其生态适应策略和环境适应性差异,为黄土高原两种重要乡土树种的高效经营及其生态功能维护提供科学依据。
1 研究方法
1.1 研究区概况
研究区位于黄土高原子午岭林区北部(35°03′ — 36°37′ N,108°10′ — 109°08′ E), 属黄土高原丘陵沟壑区,海拔1200 — 1600 m。该区为半湿润季风气候,多年平均气温7.4℃,平均降雨量在550 — 600 mm,≥10℃积温2671℃,干燥度0.97。研究区土壤为原生(山坡)或次生(沟谷)黄土,厚度一般为50 — 100 m,其下为厚约80 — 100 m的红土,以石灰性褐土为主,其次为粗骨褐色土(邹厚远等,2002)。
1.2 采样方法
在植物生长旺盛季节,选取研究区典型油松和柴松林设立20 m × 20 m的标准样地,在标准样地内进行每木检尺,选择与标准木相近的植株5株,分器官采样。根系测定采用根钻法,在油松和柴松样地内随机选择9个样点,用土钻(φ = 9 cm)采集0 — 20 cm土样,从土样中捡出所有根系装袋。叶片采集成熟健康叶,枝和根按不同发育程度采样,保证粗枝和细枝,粗根和细根比例均衡,皮和干在树木胸径处(距地面1.3 m)采样,分别标记混匀后装入自封袋,并迅速带回实验基地。油松和柴松样地基本信息见表1。
1.3 测定方法
植物水分测定采用烘干法。取部分新鲜植物样品放入纸袋称重记录,然后将样品放入烘箱105℃杀青10 min,70℃烘干至恒重后称量,计算各器官含水量。另取部分样品,杀青、烘干至恒重,粉碎后过100目筛制成供试样品。植物碳(C)含量的测定采用重铬酸钾外加热法。植物氮(N)含量测定采用凯氏定氮法(Kjeldahl)(KJELTEC2300全自动定氮仪,Sweden)。植物磷(P)含量测定采用钼黄比色法(U-2800分光光度计,中国上海)。每一样品重复3次测定,测定结果以单位质量的养分含量(g ∙ kg-1)表示(鲁如坤,2000)。

表1 试验样地基本概况Tab.1 Description of sampling sites
1.4 数据分析
采用SPSS统计分析软件包(SPSS 16.0 for Windows,USA)对数据进行One-Way ANOVA方差分析,Pearson相关分析和回归分析,并采用LSD法进行多重比较。
2 研究结果
2.1 水、碳、氮和磷含量特征
油松和柴松不同器官含水量总体差异显著(P < 0.05,图1a),变化范围为207‰ — 625‰,变异系数为31.4%。两种针叶树种各器官含水量大小顺序均表现为:根>叶>干>枝>皮;其中根和叶含水量差异不明显,在600‰左右;枝和干含水量差异不明显,在500‰左右;树皮含水量最低,大约200‰。总体看来,油松叶、枝和根的含水量略高于柴松,而皮和干的含水量略低于柴松,但不同器官含水量在两树种间差异均不显著(图1a)。
两种针叶树种不同器官有机碳含量差异相对较小,变化范围在 434.0 — 596.8 g ∙ kg-1,变异系数为11.2%。油松叶和根有机碳含量最高,显著高于枝、皮和干等器官;柴松各器官有机碳含量差异不显著。总体看来,除叶和根的有机碳差异不显著外,柴松枝、皮和干的有机碳含量均显著高于油松(图1b)。
油松和柴松不同器官全氮含量差异显著,变化范围为 1.1 — 13.0 g ∙ kg-1,变异系数为 97.1%。其中,叶片全氮含量最高,分别为13.0 g ∙ kg-1和11.6 g ∙ kg-1,显著高于其他器官;树干氮含量最低,分 别 为 1.3 g ∙ kg-1和 1.1 g ∙ kg-1; 根、 枝和 皮含 量居中,为2.0 — 3.1 g ∙ kg-1。总体看来,油松叶和皮的氮含量显著高于柴松,其他器官的氮含量差异不显著(图1c)。
油松和柴松不同器官全磷含量变化范围为1.3 — 2.4 g ∙ kg-1,变异系数为 22.9%。其中,叶片全磷含量最高,分别为 2.4 g ∙ kg-1和 2.3 g ∙ kg-1,显著高于其他器官;根系全磷含量最低,均为1.3 g ∙ kg-1;根、枝和皮含量居中,为 1.4 — 1.8 g ∙ kg-1。总体看来,不同器官全磷含量在两树种间差异均不显著(图1d)。
2.2 水、碳、氮和磷生态化学计量特征
除树皮外,油松和柴松不同器官水碳比差异相对较小,变化范围在0.77 — 1.14,变异系数为13.1%。其中,油松叶、枝、干和根的水碳比差异均不显著;但显著高于树皮。柴松根系水碳比最高,显著高于其他器官,叶、枝和干的水碳比居中,树皮的水碳比最低。总体看来,油松叶、枝和干的水碳比显著高于柴松,而树皮和根系的水碳比在两树种间无显著差异(图2a)。
两种针叶树种不同器官水氮比差异显著,变化范围在44.8 — 499.1,变异系数为78.4%。其中,油松和柴松均表现为树干的水氮比最大,显著高于其他器官;枝和根的水氮比居中,显著高于叶和皮,叶片的水氮比最低。两树种各器官水氮比大小顺序均为:干>根>枝>皮>叶。总体看来,柴松树干和皮的水氮比显著高于油松,其他器官间差异不显著(图2b)。
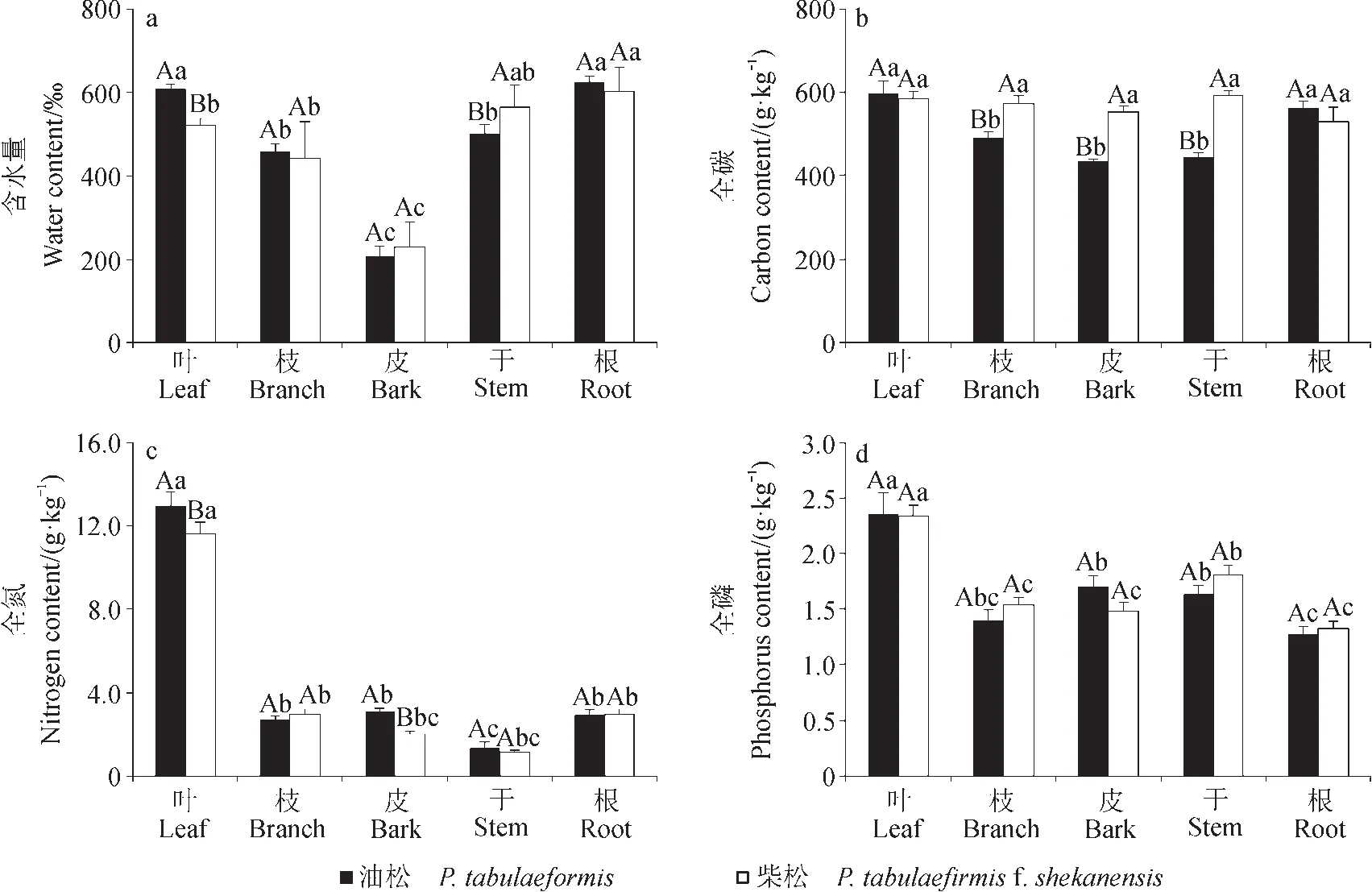
图1 油松和柴松不同器官水、碳、氮和磷含量特征Fig.1 Water, carbon, nitrogen and phosphorus content of P. tabulaeformis and P. tabulaef i rmis f. shekanensis in different organs
油松和柴松不同器官水磷比变化范围在121.6 — 491.7,变异系数为39.6%。其中,两种针叶树种均表现为根系的水磷比最大,显著高于其他器官;其次为干、枝和叶,皮的水磷比最低。两树种各器官的水磷比大小顺序均为:根>枝>干>叶>皮。总体看来,油松叶、枝和根的水磷比略高于柴松,皮和干的水磷比略低于柴松,但其差异均不显著(图2c)。
两种针叶树种不同器官碳氮比变化范围在45.9 — 524.9,变异系数为78.4%。其中,油松树干的碳氮比最大,显著高于其他器官;其次为根和枝,再次为皮,叶片碳氮比最小。柴松树干的碳氮比最高,显著高于其他器官;其次为皮,枝和根的碳氮比相近,略低于皮,叶片碳氮比最低。总体看来,油松干、皮碳氮比均显著低于柴松,而叶、枝和根的碳氮比与柴松差异不显著(图2d)。
两种针叶树种不同器官碳磷比差异变化范围在250.5 — 443.4,变异系数为20.9%。其中,油松根系的碳磷比最大,显著高于其他器官;枝的碳磷比居中,显著高于叶、皮和干;叶、皮和干碳磷比差异不显著。柴松根系的碳磷比最大,但与枝、干差异不显著,叶片的碳磷比最小。总体看来,油松皮和干的碳磷比显著低于柴松,其他器官的碳磷比差异在两树种间不显著(图2e)。
油松和柴松不同器官的氮磷比差异显著,变化范围在0.6 — 5.5,变异系数为69.3%。其中,油松和柴松均表现为叶片氮磷比最大,显著高于其他器官。油松根、枝和皮之间氮磷比差异不显著,但均显著高于干;柴松根和枝的氮磷比差异不显著,但均显著高于皮和干,且柴松皮的氮磷比也显著高于干。总体看来,油松皮的氮磷比显著高于柴松,其他器官氮磷比差异在两树种间不显著(图2f)。
2.3 水、碳、氮、磷及其生态化学计量比相关关系
油松和柴松各器官含水量与水碳比和水磷比、全氮与全磷及氮磷比、全磷与氮磷比、水碳比与水磷比、水氮比与碳氮比以及水磷比和碳磷比均呈显著正相关性(P < 0.05);全氮与碳氮比、全磷与碳磷比、水氮比与氮磷比以及碳氮比与氮磷比均呈显著负相关性(P < 0.05);其他变量间相关性不显著(表2)。
两种树种器官含水量与水碳比、水氮比与碳氮比、水磷比与碳磷比均能用线性方程较好拟合(P < 0.05);含水量与水磷比、水碳比与水磷比、全氮与碳氮比和氮磷比、全磷与碳磷比、水氮比与氮磷比、碳氮比和氮磷比均能用幂函数方程进行很好的拟合(P < 0.05)。进一步分析发现,由于叶片全氮含量显著高于其他器官,导致全氮与全磷以及全磷与氮磷比线性拟合方程也呈显著正相关性,若去除与叶片变量相关的两个点,则全氮与全磷呈不显著负相关关系,全磷与氮磷比之间呈显著负相关性(图3)。

图2 油松和柴松不同器官水、碳、氮和磷生态化学计量特征Fig.2 The ratio of water, carbon, nitrogen and phosphorus of P. tabulaeformis and P. tabulaef i rmis f. shekanensis in different organs
3 讨论
水是植物细胞的重要组成部分,也是大多数生命活动的原料和介质。通常代谢旺盛的器官或组织含水量都很高,植物器官的含水量随木质化程度增加而减少(王宝山,2010)。本研究油松和柴松叶片和根系较其他器官含水量高,表明叶片和根系是两种树种生命活动最为旺盛的器官,其次为树干和树枝,树皮的代谢活动最弱(图1a)。此外,油松叶片含水量显著高于柴松,而树干含水量低于柴松,表明油松较柴松可能具有更积极的水分利用策略,在水分条件较好情况下油松生长较柴松快,但柴松能够更好适应相对干旱的环境。与柴松相比,油松叶片和根系的碳含量相当,但枝、干和皮中碳含量较低(图1b);相反,油松叶片氮含量显著高于柴松,而其他器官差异不明显(图1c)。两种树种对碳、氮元素的分配策略反映了油松将更多的氮分配给植物生产性器官,而柴松将更多的碳分配给植物防御性器官,这也表明柴松较油松具有更加保守的生长策略,能够更好适应逆境条件。油松和柴松全磷含量在不同器官差异不显著(图1d),一方面表明两种树种对磷的利用策略相似,另一方面也说明与水分和氮素相比,研究区的磷素供应相对充足,磷素不是该区植物生长的限制性元素。
与上述结果一致,油松叶、枝和干的水碳比显著高于柴松(图2a),而皮和干的水氮比显著低于柴松(图2b),其他器官水碳比和水氮比差异均不显著,并且两种树种不同器官水磷比差异均不显著(图2c),这也反映了油松在生产单位有机碳较柴松投入更多的水分,而柴松在树干等支撑储存器官中投入更多的氮素以抵御潜在的环境胁迫。柴松这种保守型的生长策略也反映在树干较油松具有显著高的碳氮比以及枝、皮具有较高的碳磷比上(图2d,2e)。一般认为,当氮磷比小于14时,植物生长主要受氮限制,而氮磷比大于16时,植物生长受磷限制(郑淑霞和上官周平,2006)。本研究油松和柴松各器官氮磷比均显著低于14,表明研究区两树种生长主要受氮素限制,这与我们前面推论一致。此外,油松除树皮氮磷比显著高于柴松外,两树种其他器官氮磷比差异不显著,主要是因为两树种均具有较高的磷含量和低的氮含量(图2f)。
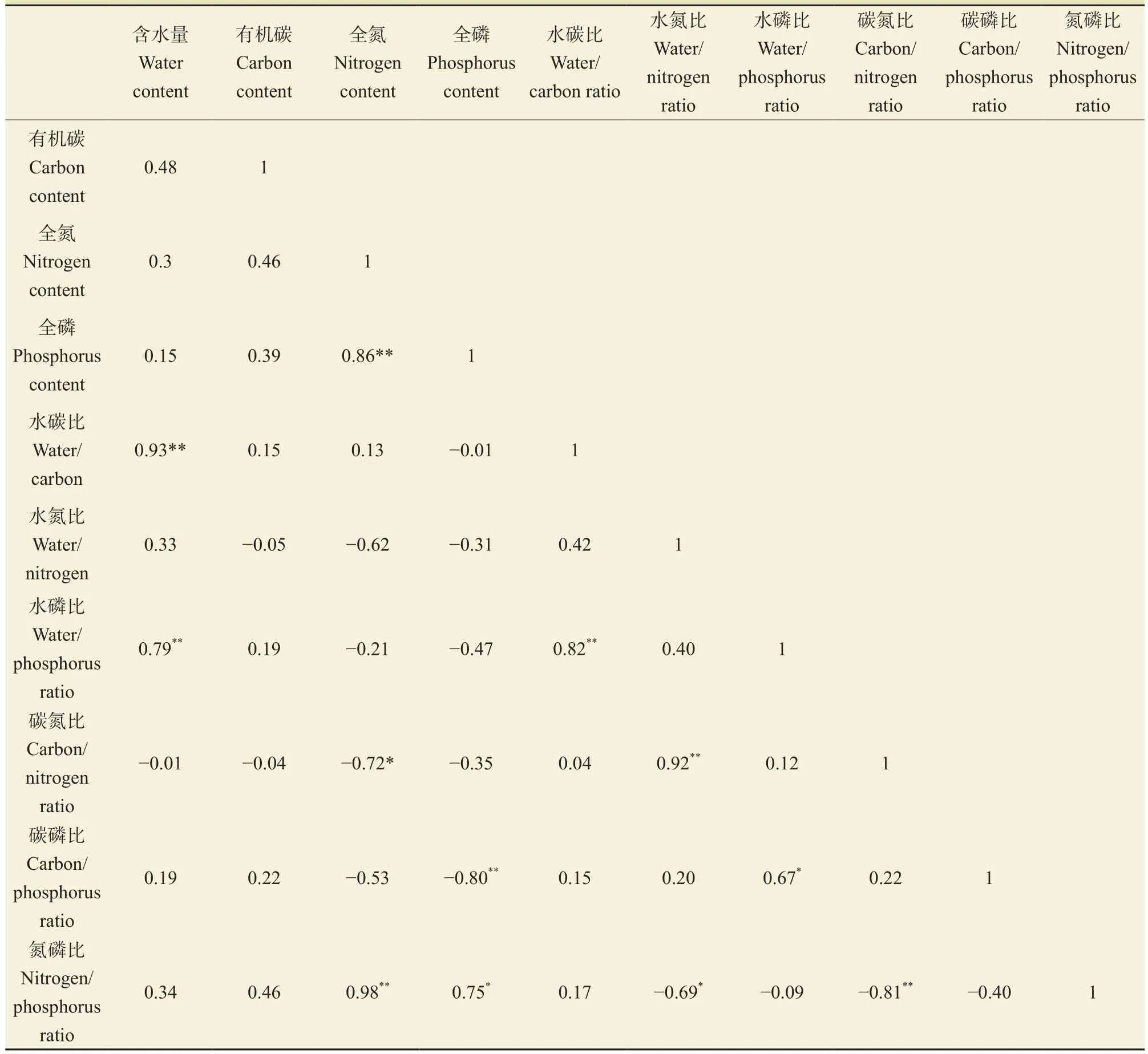
表2 油松和柴松不同器官水、碳、氮、磷及其生态化学计量比的相关关系Tab.2 Correlation coeff i cients among water, carbon, nitrogen and phosphorus content and their ratios of P. tabulaeformis and P. tabulaef i rmis f. shekanensis
本研究两树种不同器官含水量与水碳比和水磷比呈显著正相关性,并且水碳比和水磷比与含水量的关系能通过线性和幂函数高度拟合(图3a,3f),表明器官含水量是决定两种树种水碳比和水磷比的主导因素。两树种全氮与碳氮比,全磷与碳磷比均呈显著负相关性,并且二者均能通过幂函数很好的拟合(图3h,3j)。这反映了氮、磷在决定碳氮比和碳磷比中的重要性,同时也说明碳作为植物基本结构建成组分在不同器官之间的分配差异相对较小,而氮、磷作为植物生长的限制性要素在植物生产和支撑器官间分配差异较大。另外,尽管氮与磷以及磷与氮磷比也表现出较好的正相关性(表2),但从图3d和图3e可以看出,这种正相关性主要是由于两种树种叶片氮、磷含量,尤其是氮含量显著高于其他器官,从而导致该结果出现。此外,相关性及回归拟合分析还发现,两树种器官水碳比与水磷比、水氮比与碳氮比、水磷比与碳磷比均呈显著正相关性(图3g,3b,3c),而水氮比与氮磷比、碳氮比与氮磷比均呈显著负相关性(图3k,3l),表明尽管植物不同器官某两种元素之间可能没有表现出显著相关性,但植物不同器官元素间通过相互耦合后可以表现出较好的相关性,这也反映了植物不同器官元素之间具有高度复杂的协同关系,这种协同关系的体现形式可能与植物类型及其对外界环境变化的生长适应策略密切相关。

图3 油松和柴松不同器官水、碳、氮、磷及其生态化学计量比的拟合方程Fig.3 The fi tted equations of water, carbon, nitrogen and phosphorus content and their ratios of P. tabulaeformis and P. tabulaef i rmis f. shekanensis
4 主要结论
(1)油松和柴松对水、碳、氮和磷在不同器官具有不同的分配策略,其对水分和碳素的分配策略较为接近,而对氮、磷元素的分配策略较为相似。对水分和碳素的分配格局总体表现为:根和叶最大,其次为枝和干,树皮最小;对氮、磷的分配格局则表现为叶片显著高于其他器官。
(2)油松较柴松具有更高的叶片含水量和氮含量以及较低的树干含水量与枝、干和皮含碳量,表明油松将更多的水和氮分配给生产性器官,而柴松将更多的水和碳分配给防御性器官,反映了柴松较油松具有更加保守的生长策略,能够更好适应逆境条件。此外,油松和柴松磷含量在不同器官差异不显著,表明磷素可能不是该区植物生长的限制性元素。
(3)尽管植物器官不同元素间可能没有表现出显著相关性,但不同元素间通过相互耦合后可以表现出较好的相关性,反映了植物元素之间具有高度复杂的协同关系,这种协同关系的体现形式可能与植物类型及其对外界环境变化的生长适应策略密切相关。
安 慧, 韦兰英, 刘 勇, 等. 2007. 黄土丘陵区油松人工林和白桦天然林细根垂直分布及其与土壤养分的关系 [J]. 植物营养与肥料学报, 13(4): 611 – 619. [An H,Wei L Y, Liu Y, et al. 2007. Distribution characters of fi ne root of artificial Pinus tabulaeformis and natural Betula platyphylla forests and their relation to soil nutrients in Hilly Loess Regions [J]. Plant Nutrition and Fertilizer Science, 13(4): 611 – 619.]
韩文娟, 张文辉, 何景峰, 等. 2012. 不同坡向油松人工林建群种种群结构及群落特征分析[J]. 西北农林科技大学学报(自然科学版), 40(3): 47 – 55. [Han W J, Zhang W H, He J F, et al. 2012. Analysis of population structure and plantation community characteristics of Pinus tabulaeformis plantations in Loess Plateau in different slope [J]. Journal of Northwest A & F University (Natural Science Editon), 40(3): 47 – 55.]
郝江勃, 王孝安, 郭 华, 等. 2010. 黄土高原柴松群落空间结构 [J]. 生态学杂志 , 29(12): 2379 – 2383. [Hao J B, Wang X A, Guo H, et al. 2010. Spatial structure of Pinus tabulaeformis f. shekannesis community on Loess Plateau [J]. Chinese Journal of Ecology, 29(12): 2379 – 2383.]
贺合亮, 阳小成, 李丹丹, 等. 2017. 青藏高原东部窄叶鲜卑花碳、氮、磷化学计量特征 [J]. 植物生态学报,41(1): 126 – 135. [He H L, Yang X C, Li D D, et al. 2017.Stoichiometric characteristics of carbon, nitrogen and phosphorus of Sibiraea angustata shrub on the eastern Qinghai-Xizang Plateau [J]. Chinese Journal of Plant Ecology, 41(1): 126 – 135.]
贺金生,韩兴国. 2010. 生态化学计量学:探索从个体到生态系统的统一化理论 [J]. 植物生态学报, 34(1):2 – 6. [He J S, Han X G. 2010. Ecological stoichiometry:Searching for unifying principles from individuals to ecosystems [J]. Chinese Journal of Plant Ecology, 34(1):2 – 6.]
焦 醒, 刘广全. 2009. 陕西黄土高原油松生长状况及其影响因子分析 [J]. 西北植物学报, 29(5): 1026 – 1032.[Jiao X, Liu G Q. 2009. Growth and its inf l uencing factors of Pinus tabulaeformis in the Loess Plateau of Shaanxi Province [J]. Acta Botany Boreal-Occident Sinica, 29(5):1026 – 1032.]
刘 超, 王 洋, 王 楠, 等. 2012. 陆地生态系统植被氮磷化学计量研究进展 [J]. 植物生态学报, 36(11):1205 – 1216. [Liu C, Wang Y, Wang N, et al. 2012.Advances research in plant nitrogen, phosphorus and their stoichiometry in terrestrial ecosystems: a review [J].Chinese Journal of Plant Ecology, 36(11): 1205 – 1216.]
鲁如坤. 2000. 土壤农业化学分析方法 [M]. 北京: 中国农业科技出版社. [Lu R K. 2000. Soil agricultural chemistry analysis methods [M]. Beijing: Chinese Agricultural Science and Technology Press.]
申家朋, 张文辉, 李彦华, 等. 2013. 黄土高原丘陵区退耕还林地油松人工林碳储量及分配特征研究 [J]. 西北植物学报 , 33(11): 2309 – 2316. [Shen J P, Zhang W H,Li Y H, et al. 2013. Characteristics of carbon storage and sequestration of Pinus tabulaeformis forest land converted by farmland in Loess hilly area [J]. Acta Botany Boreal-Occident Sinica, 33(11): 2309 – 2316.]
孙美美, 关晋宏, 吴春荣, 等. 2017. 黄土高原西部3个降水量梯度近成熟油松人工林碳库特征 [J]. 生态学报,37(8): 1 – 8. [Sun M M, Guan J H, Wu C R, et al. 2017.Carbon storage features of Pinus tabulaeformis plantations that are approaching maturity at three sites across a precipitation gradient in western Loess Plateau [J]. Acta Ecological Sinica, 37(8): 1 – 8.]
王宝山. 2010. 逆境植物生物学 [M]. 北京: 高等教育出版社:103-106. [Wang B S. 2010. Plant biology under stress [M].Beijing: High Education Press: 103-106.]
王凯博, 上官周平. 2006. 黄土高原子午岭天然柴松林种群结构与动态研究 [J]. 西北植物学报, 26(12): 2553 – 2559.[Wang K B, Shangguan Z P. 2006. Structure and dynamics of natural Pinus tabulaeformis f. shekannesis populations in Ziwuling forest region of the Loess Plateau [J]. Acta Botany Boreal-Occident Sinica, 26(12): 2553 – 2559.]
王 梅, 张文辉. 2009. 不同密度油松人工林生长更新状况及群落结构 [J]. 西北农林科技大学学报(自然科 学 版), 37(7): 75 – 80. [Wang M, Zhang W H. 2009.Growth, regeneration and community structure of Pinus tabulaeformis artif i cial forest under different densities [J].Journal of Northwest A & F University (Natural Science Editon), 37(7): 75 – 80.]
韦兰英, 上官周平. 2007. 黄土高原子午岭天然柴松林细根垂直分布特征 [J]. 西北农林科技大学学报(自然 科 学 版), 35(7): 69 – 74. [Wei L Y, Shangguan Z P.2007. Vertical distribution of the fine root of the natural Pinus tabulaeformis f. shekannesis forest in Ziwuling Loess Plateau [J]. Journal of Northwest A & F University(Natural Science Editon), 35(7): 69 – 74.]
杨晓梅, 程积民, 孟 蕾. 2010. 黄土高原天然柴松林碳储量与碳密度特征 [J]. 中国水土保持科学, 8(2):41 – 45. [Yang X M, Cheng J M, Meng L. 2010. Carbon storage and density features of natural forest of Pinus tabulaeformis f. shekannesisin Loess Plateau [J]. Science of Soil and Water Conservation, 8(2): 41 – 45.]
曾德慧, 陈广生. 2005. 生态化学计量学: 复杂生命系统奥秘的探索 [J]. 植物生态学报, 29(6): 1007 – 1019. [Zeng D H, Chen G S. 2005. Ecological stoichiometry: a science to explore the complexity of living systems [J]. Acta Phytoecologica Sinica, 29(6): 1007 – 1019.]
郑淑霞, 上官周平. 2006. 黄土高原地区植物叶片养分组成的空间分布格局 [J]. 自然科学进展, 16(8): 965 – 973.[Zheng S X, Shangguan Z P. 2006. The distribution pattern of leaf nutrient composition in the Loess Plateau [J].Progress in Natural Science, 16(8): 965 – 973.]
邹厚远, 刘国彬, 王晗生. 2002. 子午岭林区北部近50年植被的变化发展 [J]. 西北植物学报, 22(1): 1 – 8. [Zou H Y, Liu G B, Wang H S. 2002. The vegetation development in North Ziwulin forest region in last fi fty years [J]. Acta Botany Boreal-Occident Sinica, 22(1): 1 – 8.]
Agren G I. 2008. Stoichiometry and nutrition of plant growth in natural communities [J]. Annual Review of Ecology Evolution and Systematics, 39: 153 – 170.
Elser J J, Fagan W F, Denno R F, et al. 2000. Nutritional constraints in terrestrial and freshwater food webs [J].Nature, 408: 578 – 580.
Frost P C, Evans-White M A, Finkel Z V, et al. 2005. Are you what you eat? Physiological constraints on organismal stoichiometry in an elementally imbalanced world [J].Oikos, 109(1): 18 – 28.
Gusewell S. 2004. N: P ratios in terrestrial plants: variation and functional significance [J]. New Phytologist, 164:243 – 266.
He J S, Fang J Y, Wang Z H, et al. 2006. Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grassland biomes of China [J]. Oecologia, 149(1):115 – 122.
Reich P B, Oleksyn J. 2004. Global patterns of plant leaf N and P in relation to temperature and latitude [J]. Proceedings of the National Academy of Sciences of the United States of America, 101(30): 11001 – 11006.
Sterner R W, Elser J J. 2002. Ecological stoichiometry: the biology of elements from molecules to the biosphere [M].New York: Princeton University Press.
Wagner N D, Hillebrand H, Wacker A, et al. 2013. Nutritional indicators and their uses in ecology [J]. Ecology Letters,16(4): 535 – 544.
