鹤山不同植被土壤有机碳分布特征①
2018-03-22汤松波任文丹旷远文
汤松波,习 丹,任文丹,旷远文
鹤山不同植被土壤有机碳分布特征①
汤松波1,2,3,习 丹4,任文丹1,2,3,旷远文1,3*
(1中国科学院华南植物园退化生态系统植被恢复与管理重点实验室,广州 510650;2中国科学院大学,北京 100049;3广东省应用植物学重点实验室,广州 510650;4福建农林大学林学院,福州 350002)
不同植被类型下土壤有机碳(SOC) 储量和动态变化是全球变化研究的热点之一。对南亚热带鹤山6种不同植被类型(灌草、马尾松林、桉树林、乡土树林、马占相思林、季风常绿阔叶林) SOC干湿季、空间 (0 ~ 10,10 ~ 20,20 ~ 40 cm) 变化特征、土壤惰性指数及其与土壤有效氮(TAN) 的关系研究表明:①6种植被中,干季SOC含量显著高于湿季,SOC含量随土层深度降低,马占相思林SOC含量最高,马尾松林和灌草最低;②6种植被SOC储量在0 ~ 10 cm土层所占比例最高,占0 ~ 40 cm土层SOC含量40% 以上;③土壤惰性指数随土壤深度增加而下降,常绿阔叶林、乡土树林和马占相思林烷基碳和ROC惰性指数高于桉树林和马尾松林,揭示这3种植被SOC具有更高稳定性;④SOC与土壤TAN含量呈显著正相关。结果揭示,在植被恢复过程中,选择豆科植物,辅以乡土树种营造常绿阔叶林,有利于提高森林潜在碳汇功能。
土壤有机碳;总有效氮;植被类型;季节变化;惰性指数
土壤碳库是陆地生态系统三大主要碳库(土壤、大气和植被) 中最大碳库[1],据估计,全球森林土壤碳库约为861Pg C[2]。土壤有机碳(soil organic carbon,SOC) 作为森林土壤碳库的重要组成部分,对调控全球气候变化具有重要作用[3]。森林SOC主要来自凋落物输入和根系周转,并在土壤生物作用下以CO2输出,实现大气-植被-土壤碳库间的循环[4]。受人类活动影响,全球森林面积锐减,森林土壤固碳能力被严重削弱[5];同时,全球变化背景下,森林SOC储量和动态已发生显著变化,森林土壤正逐渐由碳汇转变为碳源[6],对全球碳平衡造成极大影响。
森林SOC受植被类型[7]、土壤氮含量[8]、土壤水分[9]和土壤微生物群落结构[8]等多种因素影响,不同森林生态系统SOC存在较大的时空差异。在时间上,过去认为SOC随森林演替呈现先增加后稳定趋势。但最新研究表明,成熟森林也具有一定的固碳能力[10];在空间上,SOC随着土壤深度增加而下降,且主要集中在0 ~ 100 cm土层[11-12];另外,植被类型对SOC影响很大,通常以常绿针叶林最大(约6.81 Pg C),其次是常绿阔叶林、落叶针叶林、落叶阔叶林和混交林[13-14]。
惰性有机碳 (recalcitrant organic carbon,ROC) 是土壤碳库中理化性质相对稳定的部分。根据质谱分析结果可将ROC分为烷基碳(Alkyl-C)、烷氧碳(O-alkyl-C)、芳香碳(Aromatic-C) 和羧基碳(Carboxyl-C)[15]4种组分。随着凋落物分解进行,ROC在土壤碳库中所占比例逐渐增加,但当土壤中ROC/TOC (土壤总有机碳) 值达到60% ~ 65% 时将会保持稳定或稍有减少[16];植被类型对土壤ROC含量影响显著[17],如阔叶林或物种多样性较高的森林其ROC含量要大于针叶林或其他纯林[17-18],固氮植物类型土壤ROC含量大于非固氮植被类型[19]等;此外,环境变化对ROC也影响较大,如大气CO2浓度升高有利于土壤ROC积累[20]。
本研究以南亚热带地区鹤山不同植被类型(灌草、马尾松林、桉树林、乡土树林、马占相思林、常绿阔叶林) 不同深度(0 ~ 10,10 ~ 20,20 ~ 40 cm) 土壤为对象,分析SOC季节、空间变化规律及其与土壤总有效性氮(total available nitrogen,TAN,定义为NO– 3-N与NH4+-N之和) 的关系,并基于不同植被下ROC (烷基碳,烷氧碳,芳香碳,羧基碳) 比例及惰性指数空间变化规律,以期揭示不同植被下SOC稳定特征,为退化生态系统植被恢复过程中构建高效固碳人工林树种的选择和搭配提供理论依据。
1 材料与方法
1.1 样点概述
研究点为中国科学院鹤山森林生态系统国家野外科学观测研究站,属典型南亚热带季风气候,具明显干湿季节交替特征。年平均气温为21.7 ℃,年均降水量为1 700 mm[21]。森林土壤为赤红壤,呈酸性。早期受人为干扰严重,部分植被退化为荒坡地。自1984 年起,鹤山站先后通过营造多种人工林开展植被恢复研究,并构建林、果、草、渔复合农业生态系统示范模式。本研究的6 种植被类型为:①灌草(shrub-grass forest, SF):主要优势种为芒萁()及一些小灌木;②马尾松林(masson pine forest, PF):林龄30 a以上,胸径20 ~ 30 cm,盖度25% ~ 35%,林下有豹皮樟()、银柴()、阴香()、九节()等;③桉树林(eucalyptus forest, EF):树高15 m 以上,盖度25% ~ 35%,林下有红车 ())、浓子茉莉 ()、紫玉盘()等;④乡土树林(native-species forest, NF):林龄25 a以上,盖度75% ~ 85%,主要以荷木()为主,伴生九节()、银柴()、潺槁木姜子()等;⑤马占相思林(acacia forest, AF):林龄25 a以上,乔木层盖度40% ~ 60%;胸径20 ~ 25 cm,林下有竹节树 ()、白车 ()、黑咀蒲桃 ()等;⑥季风常绿阔叶林(monsoon evergreen broadleaved forest, MF):盖度达95% 以上,主要有小果山龙眼()、黄桐(Benth.)和橄榄[(Lour.) Raeusch.]等高大乔木,林下灌木与草本植物种类较少,地表凋落物较多。本研究中6种植被类型立地条件见表1。

表1 鹤山6种植被类型立地条件
注: 表中“–”表示数据未记录。
1.2 土样采集与化学分析
分别从上述6种不同植被样地上、中、下3 个坡位,用内径2 cm、长60 cm 土钻分0 ~ 10、10 ~ 20、20 ~ 40 cm 土层采集土样。每个样地内采集10 管土合成1个混合样,每个样地保证3个混合样。土样带回实验室自然风干,挑去根系和石砾,粉碎,过2 mm 筛干燥保存,备分析。土样采集时间为2011年8 月(代表湿季)和2012年2 月(代表干季)。
土壤SOC 测定采用K2Cr2O7-H2SO4油浴外加热法。土壤NO– 3-N和NH4+-N采用凯氏定氮法测定,二者之和即为土壤总有效性氮(TAN)。烷基碳、烷氧碳、芳香碳、羧基碳测定方法见Mathers等[15]和张晋京等[22]:称量5 g风干土于100 ml 塑料离心管中,加50 ml HF 溶液(10%,/),盖上盖子,震荡 1 h,离心 10 min (3 000 r/min),上清液去掉,残余物继续用 HF 溶液处理,重复8次,8次处理过程中样品在摇床上振荡的时间为:第 1 ~ 4 次均1 h,第5 ~ 7 次均12 h,第8次为24 h。土壤样品经 HF 处理完以后,用双蒸水洗残余物4次,去除土壤样品中残留的 HF。经 HF 处理的残余物,在 40 ℃的烘箱中烘干,磨细过60目。将经 HF 预处理过的土壤样品进行固态魔角旋转-核磁共振测定(AVANCE II 300 MH)。试验采用 7 mm CPMAS 探头,观测频率为 100.5 MHz,MAS 旋转频率为 5 000 Hz,接触时间为 2 ms,循环延迟时间为 2.5 s。化学位移的外标物为六甲基苯(Hex methylbenzene,HMB,甲基17.33)。所测图谱可分为4 个共振区域: 0 ~ 50 (烷基碳)、50 ~ 110 (烷氧碳)、110 ~ 160 (芳香碳)、160 ~ 220 (羧基碳)。对谱峰曲线进行区域积分,获得4 种碳化学组分的相对含量,根据公式 (烷基碳+ 芳香碳/(烷氧碳 + 羧基碳)[23]计算出惰性指数。土壤容重参考文献[24-25],土壤碳密度和碳储量分别按照如下公式计算:
SOCD= ∑(1-θ) × BD×C×T/100 (1)
SOCS = Area× SOCD(2)
式中:SOCD:层土壤碳密度(kg/m2);θ:第层砾石含量(%,/);BD:第层土壤容重(g/cm3);C:第层土壤有机碳含量(C,g/kg);T:第层土层厚度(cm)。SOCS:层碳储量(kg);Area:林型面积(m2)。
1.3 数据分析
土壤SOC含量的差异显著性采用多因素方差分析(Multi-way ANOVA)检验和新复极差法(SSR) 比较不同林型和不同土层之间的差异。SOC与土壤TAN相关性采用线性相关进行分析。
2 结果与分析
2.1 SOC随季节、土层深度及植被类型变化规律
三因素方差分析结果(表2) 表明,6种植被SOC含量受季节、土层深度、植被类型及其交互作用影响显著(< 0.05)。SOC含量干季显著大于湿季,在0 ~ 10 cm土层含量最高,且随土层深度显著降低(< 0.05);在6种植被中,SOC含量表现为马占相思林> 常绿阔叶林> 桉树林> 乡土树林= 马尾松林=灌草,马占相思林SOC显著高于其他5种植被(< 0.05)。

表2 季节、土壤深度和植被类型对SOC影响三因素方差分析结果
2.2 SOC储量及分布特征
6种植被土层SOC 季节变化特征见图1,除桉树林和乡土树林20 ~ 40 cm土壤层外,其余植被类型干季SOC含量均高于湿季,灌草0 ~ 10 cm、松林10 ~ 20 cm、桉树林10 ~ 20 cm、马占相思林10 ~ 20 cm、常绿阔叶林10 ~ 20 cm和20 ~ 40 cm土壤层干湿季差异均达到显著水平(< 0.05),其他土层SOC在干湿季无显著差异。

(SF:灌草,PF:马尾松林,EF:桉树林,NF:乡土树林,AF:马占相思林,MF:季风常绿阔叶林,下图同;误差柱表示一倍的标准误(SE),*表示干湿季差异显著(P < 0.05))
6种不同植被SOC含量在湿季和干季垂直变化特征相似(图2),除湿季松林外,0 ~ 10 cm土层SOC含量均显著高于(< 0.05) 10 ~ 20 cm和20 ~ 40 cm 土层,但10 ~ 20 cm与20 ~ 40 cm土层SOC含量无显著性差异(> 0.05)。方差分析显示,在0 ~ 10 cm土层,SOC含量表现为马占相思林显著(< 0.05) 高于其他几种植被,而马尾松林显著(< 0.05) 低于桉树林和常绿阔叶林,其余植被间无显著性差异(> 0.05);在10 ~ 20 cm土层,马占相思林和常绿阔叶林SOC含量相似且显著(< 0.05) 高于其余植被类型SOC;20 ~ 40 cm土层SOC含量在不同植被间无显著差异。

(图中大写字母不同表示同一土层不同植被类型间差异达到P < 0.05显著水平,小写字母不同表示同一植被类型不同土层间差异达到P < 0.05显著水平,下同)
就储量看,无论干季还是湿季,0 ~ 40 cm土层中总SOC有43% 以上储存在0 ~ 10 cm土层中 (表3);对0 ~ 10 cm土层 SOC储量所占比例方差分析表明,6种植被类型0 ~ 10 cm 土层SOC储量差异不显著(> 0.05),采样季节、植被类型对0 ~ 10 cm土层SOC储量在0 ~ 40 cm土层总SOC储量比例影响不显著。但马占相思林0 ~ 10 cm土层SOC占0 ~ 40 cm土层SOC百分数最高,表明相对其他植被,豆科植物马占相思能有效在森林土壤表层累积SOC。
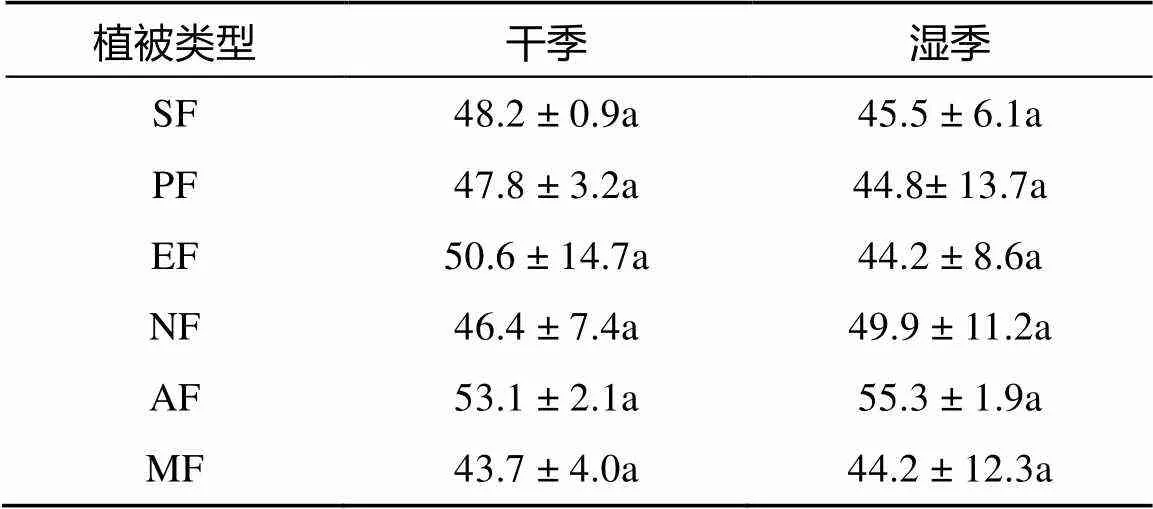
表3 干湿季0 ~ 10 cm土层SOC储量占0 ~ 40 cm土层SOC比例(%)
注: 表中小写字母相同表示不同植被间差异没有达到显著水平(>0.05)。
2.3 惰性碳组分及惰性指数随植被类型的变化
6种植被土壤在各土层中4种ROC组分比例从大到小顺序依次为:烷氧碳 > 烷基碳> 羧基碳> 芳香碳。烷基碳比例除季风阔叶林和马占相思林外,均随土壤深度增加而下降(图3A),烷氧碳和羧基碳比例随土壤深度增加而增大(图3B、3D)。对6种植被各土层惰性指数分析结果表明,惰性指数均随土层深度降低(表4),变化趋势与烷基碳一致。常绿阔叶林惰性指数各层均最大,马尾松林、乡土树林和桉树林土壤惰性碳分别在0 ~ 10、10 ~ 20和20 ~ 40 cm土层中最小。
对不同土层中4种ROC组分与ROC含量作相关分析发现,在0 ~ 10 cm土层除芳香碳外(= 0.034),其余组分与ROC含量均无显著相关(相关系数及值分别是:1 = –0.738,= 0.094;2 = –0.417,= 0.41:3 = 0.846,= 0.034;4 = 0.480,= 0.336);10 ~ 20 cm和20 ~ 40 cm土壤层各组分与ROC含量均无显著相关性,且土壤ROC含量随芳香碳增加而增加,表明芳香碳对森林表层土壤中ROC的稳定性具有一定影响。
2.4 SOC与TAN相关性
6种植被下土壤TAN含量空间分布规律与SOC相似,随土层深度降低。除马尾松林和湿季常绿阔叶林外,0 ~ 10 cm土层TAN含量均显著(< 0.05) 高于10 ~ 20 cm和20 ~ 40 cm土层。干季,以马占相思林高,季风常绿阔叶林最低 (图4);湿季,0 ~ 10 cm土层TAN含量则以灌草和乡土树林含量最高,常绿阔叶林最低(图4)。
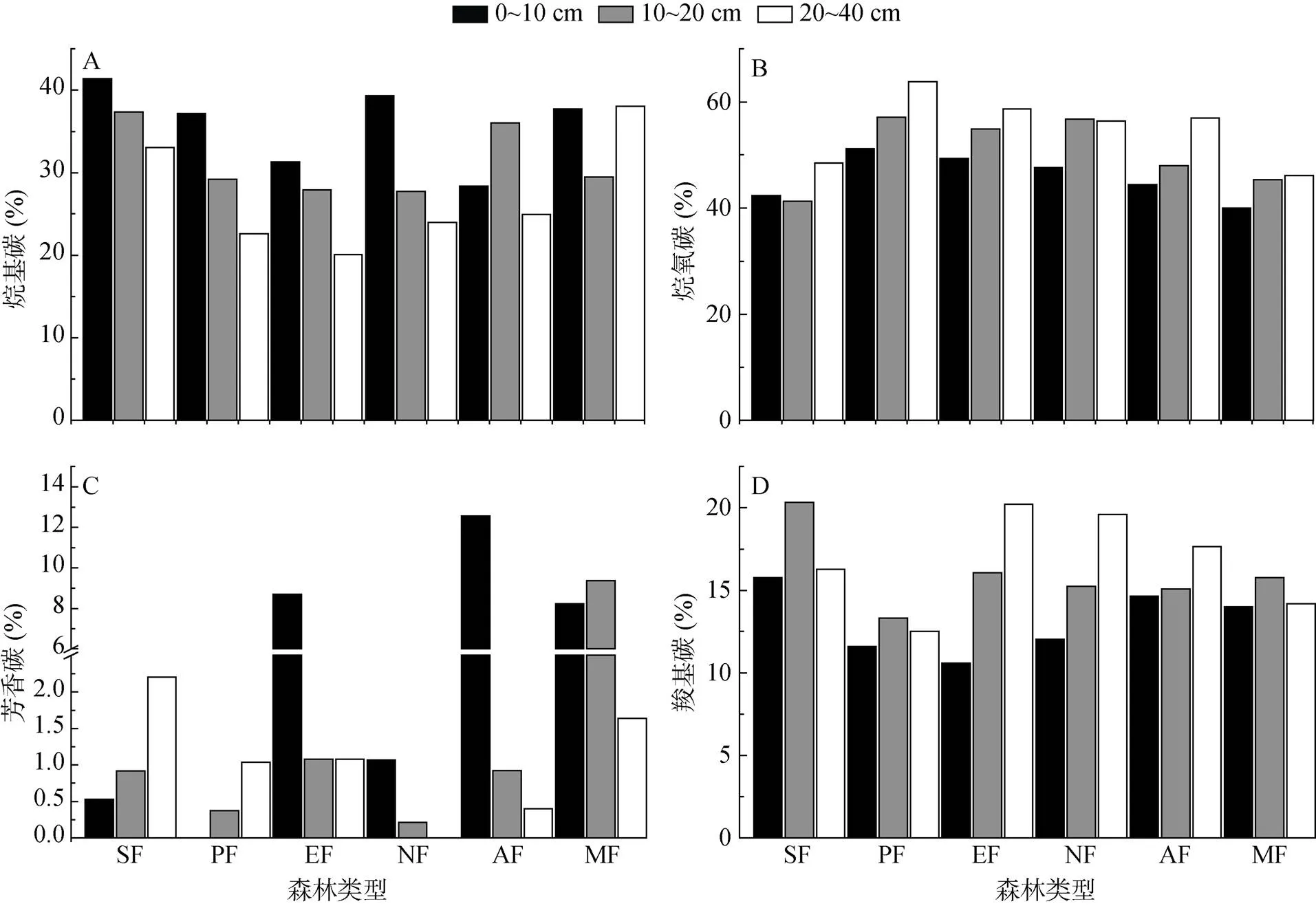
(A. 烷基碳;B. 烷氧碳;C. 芳香碳;D. 羧基碳)

表4 不同植被类型不同土层惰性指数
不同植被SOC含量与TAN相关性分析发现,干季除马尾松林外,其余5种植被SOC含量与TAN呈显著正相关;湿季,除常绿阔叶林外,其他5种植被SOC含量与TAN也具显著正相关性(图5)。
3 讨论
3.1 SOC季节与空间变化解析
森林SOC含量主要受凋落物输入、死亡根分解、土壤微生物活动、土壤理化性质等因子影响。研究表明,湿季土壤呼吸显著大于干季,且与土壤微生物生物量、细根生物量及土壤碳存在显著正相关[26],凋落物分解速率湿季显著高于干季[27];降水量也与森林SOC周转速率呈正相关[28]。本研究中,除乡土树林外,其余植被类型SOC在湿季均小于干季或具有下降趋势(图1)。其主要原因可能在于:湿季土壤含水量及土壤温度高于干季,土壤微生物活动增强,加速SOC分解[29];同时,湿季植物生长比干季快,植物凋落物、细根输入量大,受激发效应影响SOC分解加快[30]。

图4 不同植被类型土壤TAN含量

(实 心点、实线代表湿季;空心点、虚线代表干季)
森林SOC大部分储存在土壤表层,研究发现0 ~ 20 cm土层的SOC占土壤1 m深SOC 50% 以上,且随土层深度迅速下降[11-12]。植被类型对SOC在土壤中的垂直分布影响显著[12],不同植被下0 ~ 20 cm土壤中SOC占1 m土壤总SOC比例以森林最大,草地次之,灌丛最少[7]。深层土壤SOC主要由稳定性有机质输入量决定[31],还与土壤自身结构、质地紧密相关[32]。本研究发现0 ~ 10 cm土层SOC含量超过0 ~ 40 cm土层40% 以上(表3),无论干湿季,马占相思0 ~ 10 cm土层SOC最高,灌草、马尾松林最低,可能与植物生长和群落生产力有关。马占相思林作为豆科植物,具有较高生产力,且土壤微生物对氮素获取并不完全依赖于SOC分解;灌草和马尾松林可能由于生产力较低,尤其是灌草地表凋落物积累相对较少,表层SOC含量较低,植被根系主要分布浅层土壤,深层土壤SOC的输入也相对较少。因此,在植被恢复前期,选择豆科树种能有效提高表层土壤SOC含量。
3.2 土壤速效氮对SOC的影响
碳和氮参与生态系统诸多过程,如物质循环、能量流动,且互相耦合[33]。氮主要通过促进植物生长、提高生态系统生产力来增加凋落物输入[34-36];另一方面,生态系统氮饱和容易导致土壤酸化,抑制土壤微生物活动,减少SOC分解[8]。鹤山6种植被土壤TAN含量存在明显干湿季变异特征(图4),但6种植被SOC含量均与TAN呈显著正相关 (图5),可能原因在于:鹤山植被均属于非氮限制,尤其是马占相思林本身具固氮功能,植物具有较高生产力;其次,在氮沉降增加背景下[37],鹤山森林土壤酸化对土壤微生物具有抑制作用,SOC分解减少[38]。尽管灌草和乡土树林湿季TAN含量高于另外4种植被,但其SOC含量仍较低,可能由于灌草郁闭度较低,减少了林冠对大气氮沉降截留效应[39-41],加之凋落物输入少,使SOC处于较低水平。随植被恢复时间的延长,SOC逐渐积累,但土壤中氮逐年减少[42]。因此,在植被恢复过程中,生态系统固碳能力受土壤氮限制。
3.3 ROC组分和惰性指数特征及其在植被恢复中的指示意义
SOC是植被退化生态系统恢复效果的重要指标之一。ROC对SOC稳定性贡献较其他组分大,ROC惰性指数被广泛关注,惰性指数越高,SOC稳定性越大[23, 36]。如Jien 等[43]和陈小梅等[44]发现,松林凋落物中烷氧碳含量多于季风林和混交林,而烷基碳含量后者高于前者,并推测SOC稳定性与土壤惰性指数正相关。本研究中,季风常绿阔叶林惰性指数最高(表4),可能与其较高烷基碳含量有关(图3);6种不同植被类型惰性指数均随土壤深度下降,其原因可能是土壤中小分子碳水化合物及蛋白质可以形成土壤胶质体或被黏土矿物质通过物理过程等方式长期保存在深层土壤中[45];同时,生态系统凋落物输入增加可以产生激发效应,使深层土壤宿存、惰性有机碳稳定性下降,分解加速[30, 46],导致深层土壤惰性指数降低。惰性指数随土壤深度变化规律提示我们在研究生态系统固碳过程时不能简单认为土壤越深SOC越稳定或惰性碳含量越高。本研究中虽然常绿阔叶林和灌草惰性指数高于其他植被类型,但由于灌草凋落物分解速率快[47],导致土壤中烷氧碳含量偏低(表4),其固碳能力并不强;而常绿阔叶林中凋落物含量高,加之较高的生产力和惰性指数(表4),表现出很强的固碳能力。
本研究还发现,芳香碳对表层土壤ROC含量具有重要影响,同Ma 等[48]研究结果相似;同时,对不同土层惰性碳组分分析结果显示(图3),烷基碳对惰性指数影响较大,且烷基碳含量越高,惰性指数越大。因此增加凋落物中具有较高烷基碳、芳香碳、酚类化合物或它们的前体化合物含量的物种有助于提高森林土壤尤其是表层土壤固碳能力。在鹤山6种植被中,常绿阔叶林、乡土树林和马占相思林土壤中烷基碳含量(图3A)和惰性指数均处于较高水平,表明它们对SOC的保存能力较其他几种植被类型强。因此在选择物种对退化森林生态系统进行恢复时可以优先考虑选用,如在恢复初期选择固氮植物为先锋种,快速增加土壤表层有机质含量,再辅以乡土树种,构建稳定常绿阔叶林,在提高森林SOC储量同时,提升SOC稳定性,实现退化生态系统植被的持续性恢复。
4 结论
南亚热带鹤山森林SOC含量受季节、土壤深度和植被影响显著:干季SOC含量显著高于或具高于湿季趋势,0 ~ 10 cm土层SOC含量最高且随土层深度降低,马占相思林SOC含量最高,马尾松林和灌草最低;6种植被中,0 ~ 40 cm土层内SOC有43% 存在0 ~ 10 cm土层中,马占相思林0 ~ 10 cm土层较其他植被更容易积累SOC;常绿阔叶林、乡土树林和马占相思林烷基碳和ROC惰性指数高于桉树林和马尾松林,揭示这2种植被SOC具有更高稳定性;SOC与土壤TAN含量呈显著正相关。基于以上结果,建议在退化森林生态系统植被恢复实际中,可在初期营造豆科马占相思林,快速提高表层土壤SOC,中期辅以乡土树种营造常绿阔叶林,能利于提高森林SOC稳定性。
[1] Batjes N H. Total carbon and nitrogen in the soils of the world[J]. European Journal of Soil Science, 1996, 47(2): 151–163
[2] Pan Y, Birdsey R A, Fang J, et al. A large and persistent carbon sink in the world's forests[J]. Science, 2011, 333(6045): 988–993
[3] Lal R. Soil carbon sequestration to mitigate climate change[J]. Geoderma, 2004, 123(1/2): 1–22
[4] Davidson E A, Janssens I A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change[J]. Nature, 2006, 440(7081): 165–173
[5] Wei X R, Shao M G, Gale W, et al. Global pattern of soil carbon losses due to the conversion of forests to agricultural land[J]. Scientific Reports, 2014, 4(4): 4062
[6] Lewis S L. Tropical forests and the changing earth system[J]. Philosophical Transactions of The Royal Society B-Biological Sciences, 2006, 361(1465): 195–210
[7] Jobbagy E G, Jackson R B. The vertical distribution of soil organic and its relation to climate and vegetation [J]. Ecology Application, 2000, 10: 423–436
[8] Amend A S, Matulich K L, Martiny J B H. Nitrogen addition, not initial phylogenetic diversity, increases litter decomposition by fungal communities[J]. Frontiers in Microbiology, 2015 , 6 (109) : 109
[9] Zhang X, Zhang Y, Sha L, et al. Effects of continuous drought stress on soil respiration in a tropical rainforest in southwest China[J]. Plant Soil, 2015, 394(1/2): 343–353
[10] Zhou G Y, Liu S G, Li Z, et al. Old-growth forests can accumulate carbon in soils[J]. Science, 2006, 314(5804): 1417
[11] De Vos B, Cools N, Ilvesniemi H, et al. Benchmark values for forest soil carbon stocks in Europe: Results from a large scale forest soil survey[J]. Geoderma, 2015, 251: 33–46
[12] 杨帆, 黄来明, 李德成, 等. 高寒山区地形序列土壤有机碳和无机碳垂直分布特征及其影响因素[J]. 土壤学报, 2015, 25(6): 1226–1236
[13] 孔涛, 张德胜, 寇涌苹, 等. 浑河上游典型植被河岸带土壤有机碳、全氮和全磷分布特征[J]. 土壤, 2014, 46(5): 793–798
[14] Wang H, Liu S, Wang J, et al. Effects of tree species mixture on soil organic carbon stocks and greenhouse gas fluxes in subtropical plantations in China[J]. Forest Ecology and Management, 2013, 300: 4–13
[15] Mathers N J, Xu Z H. Solid-state C-13 Nmr spectroscopy: Characterization of soil organic matter under two contrasting residue management regimes in a 2-year-old pine plantation of subtropical Australia[J]. Geoderma, 2003, 114(1/2): 19–31
[16] Rovira P, Vallejo V R. Labile and recalcitrant pools of carbon and nitrogen in organic matter decomposing at different depths in Soil: An acid hydrolysis approach[J]. Geoderma, 2002, 107(1/2): 109–141
[17] Wang D, Wang B, Niu X. Effects of natural forest types on soil carbon fractions in north-east China[J]. Journal of Tropical Forest Science, 2014, 26(3): 362–370
[18] Wang Q K, Wang S L, Zhong M C. Ecosystem carbon storage and soil organic carbon stability in pure and mixed stands of cunninghamia Lanceolata and Michelia Macclurei[J]. Plant Soil, 2013, 370(1/2): 295–304
[19] 习丹, 李炯, 旷远文, 等. 鹤山不同植被类型土壤惰性碳含量及其季节变化特征[J]. 热带亚热带植物学报, 2013, 21(3): 203–210
[20] Cheng L, Leavitt S W, Kimball B A, et al. Dynamics of labile and recalcitrant soil carbon pools in a sorghum free-air CO2enrichment (Face) agroecosystem[J]. Soil Biology & Biochemistry, 2007, 39(9): 2250–2263
[21] 申卫军, 彭少麟, 周国逸, 等. 鹤山丘陵草坡的水文特征及水量平衡[J]. 植物生态学报, 2000, 24(2): 162–168
[22] 张晋京, 窦森, 朱平, 等. 长期施用有机肥对黑土胡敏素结构特征的影响—固态13C核磁共振研究[J]. 中国农业科学, 2009, 42(6): 2223-2228
[23] Ostertag R, Marin-Spiotta E, Silver W L, et al. Litterfall and decomposition in relation to soil carbon pools along a secondary forest chronosequence on Puerto Rico[J]. Ecosystems, 2008, 11(5): 701–714
[24] 傅声雷, 林永标, 饶兴权, 等. 中国生态系统定位观测与研究数据. 森林生态系统卷. 广州鹤山站:1998 ~ 2008[M]. 北京: 中国农业出版社, 2011: 101–103
[25] 余作岳, 彭少麟. 热带亚热带退化生态系统植被恢复生态学研究[M]. 广州: 广州科技出版社, 1996: 163
[26] Bae K, Lee D K, Fahey T J, et al. Seasonal variation of soil respiration rates in a secondary forest and agroforestry systems[J]. Agroforestry Systems, 2013, 87(1): 131–139
[27] Wang H Q, Cornell J D, Hall C A S, et al. Spatial and seasonal dynamics of surface soil carbon in the Luquillo experimental forest, Puerto Rico[J]. Ecological Modelling, 2002, 147(2): 105–122
[28] Brunn M, Spielvogel S, Sauer T, et al. Temperature and precipitation effects on delta13C depth profiles in SOM under temperate beech forests[J]. Geoderma, 2014, 235: 146–153
[29] Langley J A, Mckinley D C, Wolf A A, et al. Priming depletes soil carbon and releases nitrogen in a scrub-oak ecosystem exposed to elevated CO2[J]. Soil Biology and Biochemistry, 2009, 41(1): 54–60
[30] Bernal B, Mckinley D C, Hungate B A, et al. Limits to soil carbon stability; deep, ancient soil carbon decomposition stimulated by new labile organic inputs[J]. Soil Biology and Biochemistry, 2016, 98: 85–94
[31] Lorenz K, Lal R. The depth distribution of soil organic carbon in relation to land use and management and the potential of carbon sequestration in subsoil horizons[J]. Advances in Agronomy, 2005, 88: 35–66
[32] Grimm R, Behrens T, Marker M, et al. Soil organic carbon concentrations and stocks on Barro Colorado Island - Digital soil mapping using random forests analysis[J]. Geoderma, 2008, 146(1/2): 102–113
[33] 于贵瑞, 王秋凤, 方华军. 陆地生态系统碳-氮-水耦合循环的基本科学问题、理论框架与研究方法[J]. 第四纪研究, 2014, 34(4): 683–698
[34] Norby R J, Warren J M, Iversen C M, et al. CO2enhancement of forest productivity constrained by limited nitrogen availability[J]. Proceedings of the National Academy of Sciences of The United States of America, 2010, 107(45): 19368–19373
[35] Lebauer D S, Treseder K K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed[J]. Ecology, 2008, 89(2): 371–379
[36] Du Z, Wang W, Zeng W, et al. Nitrogen deposition enhances carbon sequestration by plantations in northern China[J]. Plos One, 2014, 9(2): 1–9
[37] Liu X, Zhang Y, Han W, et al. Enhanced nitrogen deposition over China[J]. Nature, 2013, 494(7438): 459–462
[38] Li H C, Hu Y L, Mao R, et al. Effects of nitrogen addition on litter decomposition and CO2release: Considering changes in litter quantity[J]. Plos One, 2015, 10(12): 1–18
[39] Nair R K F, Perks M P, Weatherall A, et al. Does canopy nitrogen uptake enhance carbon sequestration by trees?[J]. Global Change Biology, 2016, 22(2): 875–888
[40] Houle D, Marty C, Duchesne L. Response of canopy nitrogen uptake to a rapid decrease in bulk nitrate deposition in two eastern Canadian boreal forests[J]. Oecologia, 2015, 177(1): 29–37
[41] Zhang W, Shen W, Zhu S, et al. Can canopy addition of nitrogen better illustrate the effect of atmospheric nitrogen deposition on forest ecosystem?[J]. Scientific Reports, 2015, 5: 1–18
[42] Deng Q, Cheng X L, Yang Y H, et al. Carbon-nitrogen interactions during afforestation in central China[J]. Soil Biology and Biochemistry, 2014, 69: 119–122
[43] Jien S H, Chen T H, Chiu C Y. Effects of afforestation on soil organic matter characteristics under subtropical forests with low elevation[J]. Journal of Forest Research, 2011, 16(4): 275–283
[44] 陈小梅, 闫俊华, 林媚珍, 等. 南亚热带森林植被恢复演替中土壤有机碳组分及其稳定性[J]. 地球科学进展, 2016, 31(1): 86–93
[45] Mylotte R, Sutrisno A, Farooq H, et al. Insights into the composition of recalcitrant organic matter from estuarine sediments using Nmr spectroscopy[J]. Organic Geochemistry, 2016, 98: 155–165
[46] Iglesias Briones M J, Garnett M H, Ineson P. Soil biology and warming play a key role in the release of ‘old C’ from organic soils[J]. Soil Biology and Biochemistry, 2010, 42(6): 960–967
[47] Solly E F, Schoening I, Boch S, et al. Factors controlling decomposition rates of fine root litter in temperate forests and grasslands[J]. Plant Soil, 2014, 382(1/2): 203–218
[48] Ma W M, Li Z W, Ding K Y, et al. Stability of soil organic carbon and potential carbon sequestration at eroding and deposition sites[J]. Journal of Soils and Sediments, 2016, 16(6): 1705–1717
Characteristics of Soil Organic Carbon Under Different Forest Types in Heshan of Southern China
TANG Songbo1,2,3, XI Dan4, REN Wendan1,2,3, KUANG Yuanwen1,3*
(1 Key Laboratory of Vegetation Restoration and Management of Degraded Ecosystems, South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China; 2 University of Chinese Academy of Sciences, Beijing 100049, China; 3 Guangdong Provincial Key Laboratory of Applied Botany, Guangzhou 510650, China; 4 Forestry College, Fujian Agriculture and Forestry University, Fuzhou 350002, China)
The storage and dynamics of soil organic carbon (SOC) under different forest types is one of the important concerns of global change. We determined the seasonal (dry and wet seasons) and spatial (0–10, 10–20 and 20–40 cm depths) variations of SOC under six forest types (Shrub-grass forest, SF; Masson pine forest, PF; Eucalyptus forest, EF; Native-species forest, NF; Acacia forest, AF; Monsoon evergreen broadleaved forest, MF) in Heshan, southern China. Carbon recalcitrant indexes (RIC) and its relationships with total available nitrogen (TAN) in soils of the forest types were also detected. The results showed that: 1) the contents of SOC under 6 forest types were higher in wet season than in dry season with a declining trend with the increase of soil depth, SOC content was highest in AF while lowest in PF and SF; 2) 0–10 cm depth soils had highest proportion of SOC storage (more than 40%) for 0–40 cm depth soils; 3) RIC decreased with increase of soil depth. The higher contents of Alkyl-C and ROC in MF, NF and AF than in EF and PF implied that SOC in the former three forest types had higher stability; and 4) SOC contents significantly positively correlated with soil TAN contents in all forest types. The results suggested that in the process of vegetation restoration, the selection of legume species coupled with native species constructing evergreen broadleaved forest would improve potential carbon sink of forests.
Soil organic carbon; Total availability nitrogen; Forest types; Seasonal pattern; Carbon recalcitrant index
国家自然科学基金面上项目(No. 41471443)资助。
(kuangyw@scbg.ac.cn)
汤松波(1992 —),男,江西上饶人,硕士研究生,主要从事森林生态研究。E-mail: TangSongbo@scbg.ac.cn
10.13758/j.cnki.tr.2018.01.017
Q94;S154.1
A
