乳源性益生元对三线闭壳龟生长性能和营养成分消化率的影响
2018-03-17凯陈春山琰卢
魏 凯陈春山* 耿 琰卢 岩
(1.北京市水生野生动植物救护中心,北京,102100;2.北京动物园,北京,100044)
三线闭壳龟(Cuoratrifasciata),又称金钱龟,隶属龟鳖目(Testudines)、龟科(Emydidae)、闭壳龟属,国内主要分布于海南、广东、广西等地,国外主要分布于老挝和越南[1],国内很多学者对其生物学特性[2-4]、疾病防治[5-6]和养殖模式[7-8]等方面开展了大量研究。但随着三线闭壳龟药用保健和观赏价值被逐渐挖掘,市场需求剧增,对其野生种群的过度利用情况相当严重,从而导致其资源量在短时间内显著下降。目前,三线闭壳龟的野生资源已濒临灭绝,在平原及丘陵地区已经绝迹[9-10]。为了更好地保护三线闭壳龟的种群资源,我国于1988年将其列为国家Ⅱ级重点保护野生动物,《中国濒危动物红皮书》也将其列入极危等级[11],对其野生资源的利用进行了严格的限制。在此背景下,我国南方地区的部分养殖户从上世纪90年代开始尝试人工饲养三线闭壳龟,随着养殖技术的不断成熟,养殖个体数量也得到了较大提升,很大程度上缓解了市场对野生资源的消耗。可以说,人工养殖技术的发展是解决三线闭壳龟濒危状态的有效途径之一。但除现有的技术成果之外,仍存在一些问题,例如没有针对三线闭壳龟的专用饲料,而其他常见的龟鳖饲料在粗蛋白含量和营养物质配比方面也无法满足其日常需求。因此,如何通过改良饲料配比来实现健康养殖目的依然是我们努力的方向。目前,通过在水产饲料中添加植物提取物[12]、微生态制剂[13]等成分来促进动物生长和提高免疫能力的研究逐渐受到人们的重视,这对龟鳖养殖中的饲料配比合成也具有指导意义。
微生态制剂主要包括益生菌(Probiotics)、益生元(Prebiotics)及合生元(Synbiotics)。其中,益生元一般是指能够选择性地促进肠道有益菌群的增殖,对宿主健康及生长起到直接或间接作用的非消化性食物成分。目前常用的益生元主要是寡糖、多糖和酸化剂,常用来提高动物对饲料的消化和营养物质的吸收利用,从而降低饲料原料中主要蛋白饲料的用量。近年来,通过在水产饲料中添加益生元来促进个体生长,提高蛋白酶活性,增强宿主非特异性免疫力的研究逐渐成为热点,例如对草鱼生长和非特异性免疫能力的影响[14];对大菱鲆幼鱼生长性能、体组成的影响[15];对克氏原螯虾生长的影响[16]等研究。但目前还缺乏对三线闭壳龟个体生长、免疫能力以及饲料配比的相关研究。本研究采用在三线闭壳龟的饲料中添加一定比例的乳源性益生元(百泰®-A,其有效组分为35%的粗蛋白和53%的寡糖),以了解其对三线闭壳龟生长性能以及营养成分表观消化率的影响,为今后筛选最适宜的添加比例提供依据。
1 材料与方法
1.1 试验饲料
本试验所用饲料由中国农业科学院饲料研究所提供,基础饲料中粗蛋白含量为44.78%、粗脂肪为7.78%、总能为17.61MJ/kg,基本可以满足三线闭壳龟的日常营养需求[17-18]。在2个试验组中,分别添加0%(对照组)和1%(试验组)的益生元(百泰®-A),以G0、G1组表示,本实验所用的益生元(百泰®-A)是由美国国际原料公司提供,由自溶性啤酒酵母、奶组分和晒干的发酵产物组成的天然产品,其有效组分为35%的粗蛋白和53%的寡糖。饲料原料粉碎后经挤压制成2.0 mm的膨化颗粒饲料,原料混合时每组饲料中添加0.1% Y2O3作为指示剂,用于测定各类营养成分的表观消化率。2组饲料配方及营养成分见表1,氨基酸组成见表2。
表1 基础日粮配方和营养水平(%)
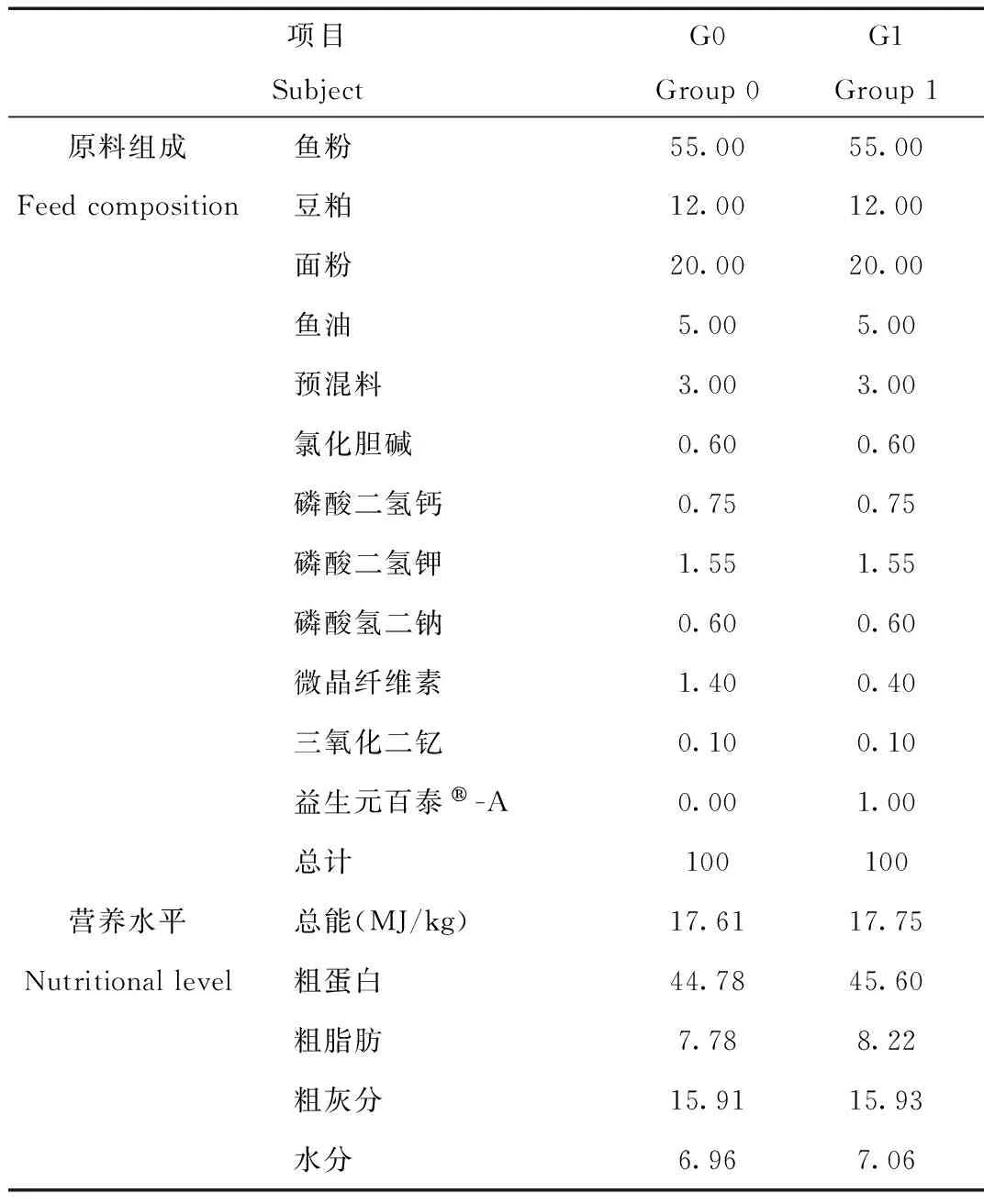
Tab.1 Diet formula and nutrition level
表2 2组饲料中的氨基酸组成(%)

Tab.2 Amino acids composition of two groups
1.2 试验龟及其饲养管理
1.2.1 试验分组
试验用龟购自广东某养殖场,均为3龄雌性个体。将30只体质健康且无疾病的三线闭壳龟,随机分为2组,经单因子方差分析(One-way ANOVA)2组间的初始体重无显著差异(P>0.05)。
1.2.2 试验时间、地点和养殖设施
试验在北京市水生野生动植物救护中心爬行动物饲养室进行,养殖时长为150 d。饲养设施采用“爬行天下”的PB-120型智能箱,规格为120 cm×50 cm×50 cm,具备UVA、UVB、陶瓷加热及通风网等设备,并采用控温器保持水温恒定(±0.1℃),试验期间水温控制在30℃。每箱养殖实验龟3只。试验开始前用20%高锰酸钾溶液刷洗缸底和缸壁,5%聚维酮碘溶液浸泡1 d后待用。
1.2.3 饲养管理
试验期间每天10:00定时投喂1次,2 h后清理残余饵料。残余饵料烘干后称重,以准确记录每组龟的摄食量,每隔30 d称量实验龟体重1次。
1.3 粪样收集
本试验的粪便样品收集方法参考沈建伟等[19],待粪样收集完毕后置于70℃烘箱烘干,-20°C冰箱备检。
1.4 分析检测
饲料中的粗蛋白、粗脂肪、水分和灰分的测定分别按照国家标准方法GB/T6432-1994、GB/T6433-2006、GB/T6435-2006和GB/T6438-1992进行检测。总能采用氧弹燃烧法测定。
饲料和粪便中的氨基酸测定按照国家标准GB/T 18246-2000进行检测。饲料和粪便中三氧化二钇(Y2O3)含量测定依据王赤龙等[20]方法,采用等离子发射光谱ICP-AES法测定。
1.5 数据计算及统计分析
1.5.1 各营养物质表观消化率
各营养物质表观消化率(Apparent digestibility coefficients,ADCS)的计算公式如下:
ADCS%=[1-(F/D×DY/FY)]×100
式中:D-饲料中某种营养成分的含量(%);F-粪便中相应营养成分的含量(%);DY-饲料中三氧化二钇(Y2O3)的含量(%);FY-粪便中三氧化二钇(Y2O3)的含量(%)。
1.5.2 生长性能的评价指标及计算方法
试验龟体重净增量、日增重量、相对增重率、饲料系数、摄食率计算如下:
体重净增量(g)=终末体重-初始体重;日增重量(g/d)=体重净增量/饲养天数;增重率(%)=(体重净增量/初始体重)×100;饲料系数=摄食量/体重净增量;摄食率(%)=摄食量/[(初始体重+终末体重)/2×饲养天数]×100;蛋白质效率(%)=(体重净增量/粗蛋白质摄入量)×100。
1.5.3 数据处理
所有数据采用“平均值±标准差”表示。所有数据经SPSS19.0软件进行方差齐性检验(Homogeneity test for variance),如果方差齐性,采用单因子方差分析(One-way ANOVA);如果方差不齐,则进行非参数检验(K-W分析),P<0.05时差异有统计学意义。
2 结果
2.1 饲料中添加乳源性益生元后对三线闭壳龟生长性能的影响
研究结果表明,饲料中添加乳源性益生元后G1组的终末体重、相对增重率、日增重量、蛋白质效率均显著高于G0组(P<0.05),而饲料系数及摄食率均显著低于G0组(P<0.05)(表3)。
表3 三线闭壳龟的生长性能(平均值±标准差)

Tab.3 Growth index of Cuora trifasciata
注:同一行数据肩标字母不同,表示存在显著差异(P<0.05)。下表同
Note:The same line of data is different from the superscript letter,which means there is a significant difference(P<0.05).The following table is the same
2.2 饲料中添加乳源性益生元后对蛋白质和干物质表观消化率的影响
结果表明,饲料中添加乳源性益生元后对G0组和G1组的蛋白质和干物质消化率分别为,G1组均显著高于G0组(P<0.05)(表4)。
表4 蛋白质和干物质表观消化率结果(平均值±标准差)(%)

Tab.4 Apparent digestibility of protein and dry matter
2.3 饲料中添加益生元后对氨基酸消化率的影响
结果表明:G0组和G1组间总氨基酸消化率没有显著差异(P>0.05)。必需氨基酸中,G1组的蛋氨酸、组氨酸、赖氨酸、亮氨酸、苏氨酸的消化率均显著高于G0组(P<0.05);非必需氨基酸中,G1组的胱氨酸、天门冬氨酸和谷氨酸的消化率显著高于G0组(P<0.05)(表5),其余的氨基酸消化率没有显著差异(P>0.05)。由于分析检测过程中色氨酸被破坏,故无法检出比较。
表5 两组饲料的氨基酸表观消化率(平均值±标准差)(%)

Tab.5 Apparent digestibility of amino acids of two groups
3 讨论
3.1 饲料中添加益生元对促进动物生长的作用
益生元作为一种微生态调节剂,具有选择性地促进有益菌的增殖、降低病原菌的致病力以及调节机体免疫系统等功能[21],相比使用传统抗生素所带来的耐药性、药物残留等风险,益生元是一种更为安全有效的促生长剂。目前,在很多水产饲料中都添加了各类益生元成分,周小秋等[22]研究了在基础饵料中添加不同比例甘果寡糖对鲤鱼生长的影响,其结果表明寡聚糖对鲤鱼有显著的促生长作用。苏纯阳等[23]研究了在基础饲料中添加一定比例的益康素(寡聚糖类产品)对南美白对虾生长性能的影响,试验组在增重、成活率、饲料系数等指标上均与对照组有显著差异。肖明松等[24]研究了在饲料中添加不同种类和含量的益生元对中华鳖生长、免疫功能和消化酶活力的影响,结果表明益生元有一定的促生长作用,并可以提高多种酶活力。本文所添加的乳源性益生元(百泰®-A)是由自溶性啤酒酵母、奶组分和晒干的发酵产物组成。Li等[25]研究了在杂交条纹鲈饵料中添加百泰®-A能提高其增重率。本试验中,通过在基础饲料中添加百泰®-A可以显著提高三线闭壳龟的增重率和蛋白质效率,同时使饲料系数显著降低。
3.2 益生元对促进营养物质吸收的作用
龟鳖和其他动物一样,为满足生理需要,要从饲料中摄取蛋白质、脂肪、碳水化合物、维生素和无机盐等营养物质,其中蛋白质是最重要的营养物质。从其生理作用来看,蛋白质是决定动物生长快慢的关键因素,Burr[26]研究发现,饲料中添加1%的百泰®-A能显著提高美国红鱼(Sciaenopsocellatus)对高豆粕日粮蛋白质的表观消化率,并能显著促进其前肠上皮细胞和微绒毛发育,这是促进其对营养物质吸收的重要因素。而龟鳖对蛋白质的需要量,其实质是对氨基酸的需要,周小秋等[27-29]研究指出赖氨酸和蛋氨酸对水生生物蛋白质的合成起到关键作用,赖氨酸缺乏会降低鳖的生活力,导致其食欲下降、生长受阻,赖氨酸对鳖的增重、饲料系数、体蛋白沉积等指标影响极显著(P<0.01);而蛋氨酸水平对特定生长率、摄食量、饲料系数有极其显著或显著影响(P<0.01或P<0.05)。因此,赖氨酸或蛋氨酸缺乏引起摄饵量下降可能是导致蛋白合成能力下降的主要原因之一。本试验中,在饲料中添加益生元虽然对总氨基酸消化率没有影响,但对赖氨酸和蛋氨酸消化率却造成显著差异(P<0.05),这可能是造成2组三线闭壳龟蛋白质合成、吸收及其他生长指标出现差异的重要原因。
3.3 单一性使用益生元的局限性
虽然很多研究[30-32]已证实益生元可以促进动物生长,有效提高动物的非特异性免疫。但也有很多国内外文献[33-34]指出益生元对试验对象的生长没有起到促进作用,除了物种之间存在的生长差异,还可能与益生元本身的作用机制有关。益生元是选择性地促进肠道有益微生物菌群的生长繁殖和代谢以及优化肠道有益微生物菌群的结构和丰度的。因此,如果没有相应益生菌的参与,动物肠道内缺乏有益微生物菌群的话,单纯使用益生元是无法竞争性抑制病原微生物的生长,而只能促进已有的有益菌群生长。因此,考虑到益生元的局限性,很多研究[35-37]已将益生菌和益生元的复合制剂(也称为合生元)应用在动物养殖环节中,它的特点是高效配伍特定的益生菌和益生元,通过有针对性地促进外源性益生菌在动物肠道中定植和生长,并选择性刺激内源有益菌群的增殖,调节免疫系统,改善动物肠道微生态群系平衡,起到益生菌和益生元的双重作用,从而更好地促进动物的生长和健康。下一步,我们也将在现有基础上,重点研究饲料中添加合生元对三线闭壳龟生长、非特异性免疫等方面的作用。
3.4 健康养殖模式的推广对野生种群资源恢复的重要意义
物种的濒危主要由环境因素、遗传因素、经济和社会发展因素所决定。对于濒危物种的资源恢复来说,开展人工养殖是一种有效的保育手段,目前所倡导的健康养殖模式则是未来人工养殖技术的主要发展方向。它是一个系统工程,是一种可持续的发展模式,可以保障动物健康和经济效益的同步发展,其主要涉及养殖配套设施、饲料管理、动物福利、疾病防治等方面。经过20多年的发展,三线闭壳龟的人工养殖已经逐渐走出了庭院式养殖,并向规模化、专业化、生态化方向发展。随着健康养殖模式的不断完善、推广,已能够满足市场对三线闭壳龟的食用数量、质量安全性和营养价值的多重需求,减少对野生种群的利用和破坏。本文从饲料配比方面入手,初步研究了益生元对三线闭壳龟的作用,有助于提高生长性能,增强其抗病能力,并减少对抗生素的依赖,在一定程度上可以缓解经济和社会发展因素对野生三线闭壳龟的压力。该成果还可以应用于物种保育技术研究,对野生种群资源恢复有一定意义。
[1] 魏成清,朱新平,陈永乐,等.三线闭壳龟的研究进展及发展对策[J].广东农业科学,2009(9):170-173.
[2] 洪美玲,王力军,史海涛.三线闭壳龟的生物学特性及人工养殖[J].海南师范学院学报:自然科学版,2003,16(3):78-82.
[3] 朱新平,陈永乐,刘毅辉,等.黄喉拟水龟、三线闭壳龟、鳄龟的生长比较研究[J].水产学报,2001,25(6):507-511.
[4] 陈春山,刘康,魏凯,等.温度和食物组成对三线闭壳龟生长的影响[J].野生动物,2012,33(2):77-80.
[5] 魏凯.人工养殖下金钱龟常见疾病及防治模式[J].野生动物,2012,33(1):34-36.
[6] 张明.金钱龟病害的防治[J].农村新技术,2011(23):27.
[7] 周婷.三线闭壳龟的饲养技术[J].四川动物,2000,19(3):184.
[8] 魏成清,朱新平,陈永乐.三线闭壳龟的人工养殖与繁育技术[J].广东农业科学,2004(4):67-69.
[9] 李贵生,唐大由.三线闭壳龟的人工保育[J].四川动物,2000,19(3):149-150.
[10] 周婷,李丕鹏.中国龟鳖物种多样性及濒危现状[J].四川动物,2007,26(2):463-467.
[11] 汪松,赵尔宓,国家环境保护局,等.中国濒危动物红皮书:两栖类和爬行类[M].北京:科学出版社,1998:29-37.
[12] 吕小文,郝倩,陈业渊,等.饲料添加芒果叶黄酮浸膏促进鱼类生长[J].农业工程学报,2013,29(18):277-283.
[13] 洪徐鹏,段燕君,周佳楠,等.微生态制剂在改善水产养殖动物免疫力中的应用[J].安徽农业科学,2014,42(22):7431-7434.
[14] 尚卫敏,朱根廷,陈任孝,等.酸化剂对草鱼生长和非特异性免疫能力的影响[J].淡水渔业,2011,41(5):56-61.
[15] 蔡胜昌.饲料中添加壳寡糖和低聚木糖对大菱鲆幼鱼生长性能、体组成、血液生化指标及非特异性免疫的影响[D].上海:上海海洋大学,2015:14-16.
[16] 洪徐鹏,夏思瑶,唐嘉荩,等.黄芪多糖对克氏原螯虾生长和非特异性免疫指标的影响[J].上海海洋大学学报,2013,22(4):571-576.
[17] 陈春山,魏凯,刘康,等.人工条件下不同饲料饲养金钱龟效果的研究[J].四川动物,2011,30(4):586-589.
[18] 李贵生,唐大由,方堃.三线闭壳龟肌肉氨基酸分析[J].四川动物,2000,19(3):165-166.
[19] 沈建伟,张永普,柳劲松,等.食物类型对平胸龟幼体摄食和消化影响的初步研究[J].淡水渔业,2008,38(2):27-30.
[20] 王赤龙,王嘉,薛敏,等.乳源性益生元(百泰®-A)对西伯利亚鲟(Acipenserbacriibrandt)生长性能和营养成分消化率的影响[J].饲料工业,2011,32(10):11-15.
[21] 吕耀平,李铁民.益生元在水产品养殖中应用的研究进展[J].饲料工业,2005,26(16):8-13.
[22] 周小秋,邝声耀,唐凌.不同剂量甘果寡糖对鲤生长和运输的影响[J].水产科学,2002,21(2):13-14.
[23] 苏纯阳,刘亚力,杨育才,等.寡聚糖对对虾生长、体成分及饲料效率的影响[J].饲料工业,2002,23(4):40-42.
[24] 肖明松,王志耕,崔峰,等.果寡糖和糖萜素对中华鳖生长性能及免疫功能的影响[J].水利渔业,2004,24(3):20-22.
[25] Li P,Gatlin D M.Dietary brewer’s yeast and the prebiotic GroBioticTM-AE influence growth performance,immune responses and resistance of hybrid striped bass(Moronechrysops×M.saxatilis)toStreptococcusiniaeinfection[J].Aquaculture,2004,231(1):445-456.
[26] Burr G S.Effects of prebiotics on growth performance,nutrient utilization and the gastrointestinal tract microbial community of Hybrid striped bass((Moronechrysops×M.saxatilis)and red drum(Sciaenopsocellatus)[M],Texas A&M University,2007:111-113.
[27] 周小秋,杨凤,周安国.赖氨酸和蛋氨酸对水生动物蛋白质合成能力的影响[J].国外畜牧科技,2001,28(3):2-5.
[28] 周小秋,杨凤,周安国,等.稚鳖赖氨酸营养的需要量[J].水产学报,2001,25(5):454-459.
[29] 周小秋,杨凤,周安国,等.稚鳖蛋氨酸的营养需要量[J].水生生物学报,2003,27(1):69-73.
[30] 李晓晶.壳寡糖对肉仔鸡促生长、免疫调节和抗氧化作用的研究[D].北京:中国农业大学,2007:28-30.
[31] 张德瑞,王际英,张利民,等.乳制品-酵母益生元(Grobiotic-A)对星斑川鲽(Platichthysstellatus)幼鱼生长、体组成及非特异性免疫的影响[C]//第九届世界华人鱼虾营养学术研讨会论文集,厦门,2013.
[32] 覃川杰,汪成竹,陈晓辉,等.茯苓多糖对中华鳖非特异性免疫功能的免疫调节作用[J].淡水渔业,2006,36(6):40-43.
[33] Pryor G S,Royes J B,Chapman F A,et al.Mannanoligosaccharides in fish nutrition:effects of dietary supplementation on growth and gastrointestinal villi structure in gulf of mexico sturgeon[J].North American Journal of Aquaculture,2003,65(2):106-111.
[34] 张荣斌,曹俊明,黄燕华,等.饲料中添加低聚木糖对奥尼罗非鱼生长性能和血清生化指标的影响[J].动物营养学报,2011,23(11):2000-2008.
[35] 王熙涛,李淑英,王丽丽,等.合生元的作用机制及其在水产养殖中的应用前景[J].饲料研究,2016(12):6-14,25.
[36] 王雪飞,李霞,付文艳,等.合生元作用机制及生理功能研究进展[J].中国兽医杂志,2010,46(9):53-56.
[37] 肖世玖,陈雁南,孙亚楠,等.合生素对团头鲂生产性能、肠道菌群及肠道形态的影响[J].中国粮油学报,2010,25(5):73-76,80.
