次生阔叶林冬季鸟类混合群生态位的相互关系
2018-03-17曹长雷邓秋香王海涛由玉岩
曹长雷邓秋香王海涛由玉岩
(1.长江师范学院生命科学与技术学院,重庆,408100;2.北华大学化学与生物学院,吉林,132013;3.东北师范大学生命科学学院,长春,130024;4.北京动物园,北京,100044)
鸟类混合群在世界范围内普遍存在,在鸟类集群研究中,主要集中在2个问题上:一是集群的功能,二是混合群内部鸟类的竞争性[1]。
国内外大量的研究描述了鸟类混合群的组成[2-8]。从集团功能研究上,主要集中在2个模式上:抵抗被捕食和捕食效率模式。抗捕食模式指出,一个种群加入集群主要是减少被捕食的机会,这种模式可以分解成2个假说:(1)“许多眼睛”假说,即当种群加入集群时会增加“很多眼睛”以提高警惕,从而减少被捕食的概率;(2)“数量安全”假说,即加入种群对集群种群是一个缓冲,可减少被捕食的概率[9-12]。更深入地说,集群可能支配着一种几何学规律从而保护内部种群免于受到捕食者的攻击[13-14]。捕食效率模式也与“眼睛”假说有关,种群增大势必使种群“眼睛”增多,利于对食物的定位,投入捕食时间增多[15-18]。然而,在降低被捕食机会的同时,捕食效率也相应降低,集群种群对资源的竞争会使捕食效率降低,因而,潜在需要调整捕食行为以适应集群中其他种群的运动模式。国内对山地次生林冬季鸟类集群做过初步研究,提出了集群的物种组成、群的个体数、集群行为、集群的时间和空间变化以及气候因子对集群形成及种群行为的影响等[19]。在对小兴安岭人工落叶松林中冬季山雀类集群的研究指出,混合群由普通鳾(Sittaeuropaea)、旋木雀(Certhiafamiliaris)、沼泽山雀(Paruspalustris)和银喉长尾山雀(Aegithaloscaudatus)组成,其中19个混合群中沼泽山雀和银喉长尾山雀的数量分别为178只和118只,占总个体数的53.1%和35.2%[20]。在对两种山雀捕食生态位和竞争的研究指出,在12个群体中,沼泽山雀和银喉长尾山雀是混合群的主体[21]。
在以往混合群内部鸟类竞争研究中,混合群内部鸟类之间的竞争报道很少。许多研究认为,种间竞争可以通过捕食行为的改变而减弱乃至避免,但种间差异以及较低的生态位重叠并不能完全说明种间竞争的重要性。对大斑啄木鸟(Dendrocoposmajor)、小斑啄木鸟(Dendrocoposminor)和灰头绿啄木鸟(Picuscanus)的取食树种、取食基质、取食高度、冠层取食位置4个资源位的生态位宽度和生态位重叠的研究指出,在4个资源位上都有分离,且与鸟的体重有明显的关系[22]。
我们此次研究的主要目的在于揭示山地次生林冬季鸟类的集群行为,找出混合群中的核心种群,明确混合群中核心种群间的关系及各自在混合群中扮演的角色与地位。
1 研究区域自然概况
研究地点为吉林省左家自然保护区,为长白山地的丘陵地带,沟壑较多。位于E 126°00′~126°04′,N 43°07′~44°06′,全区总面积为87 km2,海拔高度240~460 m(平均300 m)。保护区内主要生境是山地次生阔叶林,树龄大多50~60 a。保护区现有植物40多科,300余种,主要由14种乔木构成,优势树种为蒙古栎(Quercusmongolica)、黑桦(Betuladahurica)、辽椴(Tiliamandshurica)、白皮柳(Salixpierotii)、山杨(Populusdavidiana)、紫椴(Tiliaamurensis)、黄檗(Phellodendronamurense)等7种(表1),还有水曲柳(Fraxinusmandshurica)、山槐(Albiziakalkora)、色木槭(Acerpictum)、稠李(Prunuspadus),以及部分针叶林树种,如油松(Pinustabuliformis)、樟子松(Pinussylvestris)、黄花落叶松(Larixgmeliniivar.olgensis)等。林下灌木丛生,主要有紫丁香(Syringaoblata)、剌五加(Eleutherococcussenticosus)、山荆子(Malusbaccata)、山刺枚(Rosadavurica)、榛(Corylusheterophylla)等。除次生林外,还有草甸、农田、林间空地和水域等生态景观。
2 研究方法
2.1 样地选取
在研究地点选择5块样地,每块 10 hm2左右,用全球定位系统GPS测量,并作标号。5块样地林型均为山地次生阔叶林。
选取1号、2号样地进行优势树种的测量,每块样地中设10个随机点(标准树),以标准树为中心,以30 m为半径,调查该范围内的树种、树高、胸径和每种树的株数。据调查,研究区域优势树种的详细参数列表如下。
表1 优势树种的测量

Tab.1 Survey on dominance species of tree
2.2 鸟类调查
采用Morrison等的方法[23],在每块样地选择10棵标准树,树间距为80 m,以标准树为中心30 m为半径,每点观察5 min。记录看到的群数、种类和个体数,同时记录集群行为(单种群、多种群和单独个体)。每月观察2次,两冬季(11~2月)观察16次,计26.7 h。
鸟类捕食的位置分为树冠层、树干部、地面和空中。依据距离树干远近,树冠分为3部分:树冠内侧1/3为内侧,树冠内侧以外的1/3为中侧,树冠最外侧的1/3为外侧。树冠从下向上再划分为下1/3、中1/3、上1/3,3个垂直等级。对每种鸟的捕食位置都记录千次以上。
2.3 数据处理

3 结果
3.1 山地次生林冬季鸟类集群行为
5块样地共观察133.5 h,2003年11月~2004年2月记录60群,2004年11月~2005年2月记录98群。调查结果表明,2003~2004年混合群物种密度最大,达7.1只/群,高于2004~2005年(表2)。与混合群相比,单种群出现次数不多,且每群的物种数量明显低于混合群。冬季鸟类物种活动主要以集群方式为主。
3.2 核心种的确定
对混合群中鸟类出现频率及丰富度进行分析发现,出现频率较多的(>50%)鸟类有3种:普通鳾、大山雀、沼泽山雀。普通鳾在混合群中出现频率最高达到100%,其次是大山雀,出现频率为67%(表3)。3种鸟类出现频率及丰富度都占绝对优势,在混合群形成中起重要作用,普通鳾、大山雀、沼泽山雀为核心种。
表2 冬季鸟类集群中的个体数和群数

Tab.2 Number of individuals and flock in wintering mixed-species flocks

续表2
注:n代表该物种在混合群或单种群中出现的群数,数字代表该物种在混合群或单种群中出现的平均个体数
Note:nnumber of flock for the specie in mixed-species or single species flock.Number of table 2:Number of average individuals for the specie in mixed-species or single species flock
表3 混群鸟类中优势种的出现频率和丰富度
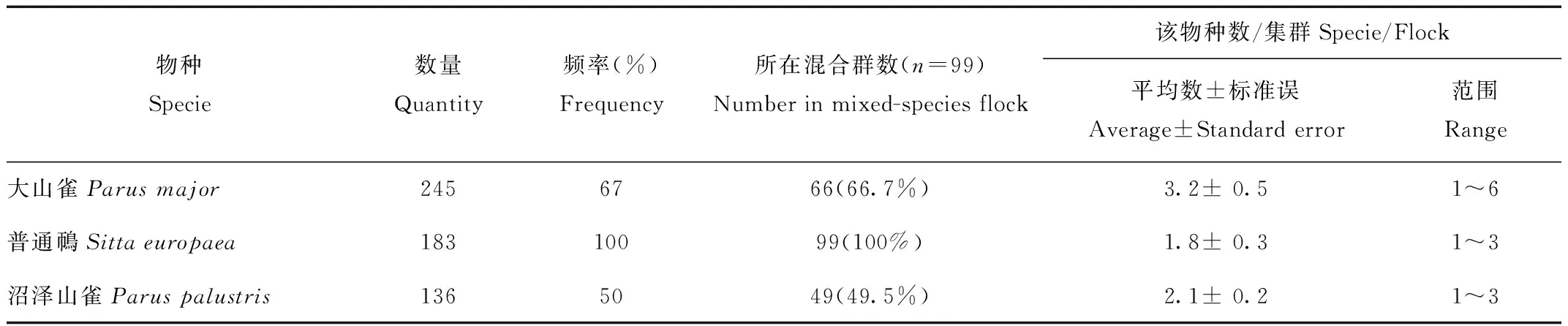
Tab.3 The frequency and richness of core species in mixed-species flocks
3.3 核心种的相互关系
3.3.1 凝聚指数
调查发现,普通鳾与大山雀的凝聚系数最大(1.26),与沼泽山雀次之(表4),因此,普通鳾与大山雀之间协同性较好,社会性程度较高,组成混合群的机率最大。
表4 核心种群的凝聚指数

Tab.4 The coherence index of core population
3.3.2 核心种的关联系数
从关联系数来看,与核心种的凝聚指数一样,普通鳾与大山雀、沼泽山雀的关联系数和凝聚指数都较高(表5),关系较为密切,是形成混合群的主体。
表5 核心种的关联系数

Tab.5 The association coefficient of core population
3.4 核心种取食位置的相互关系
调查发现,大山雀取食主要集中在地面、树冠中上层以及中外侧,沼泽山雀取食主要集中在树冠中上层以及中外侧,普通鳾取食主要集中在树干、树冠中下层以及中内侧(图1)。
Duncan多重比较表明,内侧、中侧、上层、下层以及地面等捕食位置中各种群的捕食没有差异,外侧捕食位置中沼泽山雀与普通鳾之间存在显著差异,中层捕食位置中大山雀与沼泽山雀之间存在显著差异,树干捕食位置中普通鳾与沼泽山雀、大山雀之间都存在显著差异(表6)。
表6 核心种间对捕食位置利用差异性的duncan多重比较

Tab.6 Duncan multiple comparisons for utilization difference ofpredation position by core specie in mixed-species flocks
核心种在长期种间互相作用下形成了独有的生态位,以降低生态位重叠,减少种间竞争,协同进化。
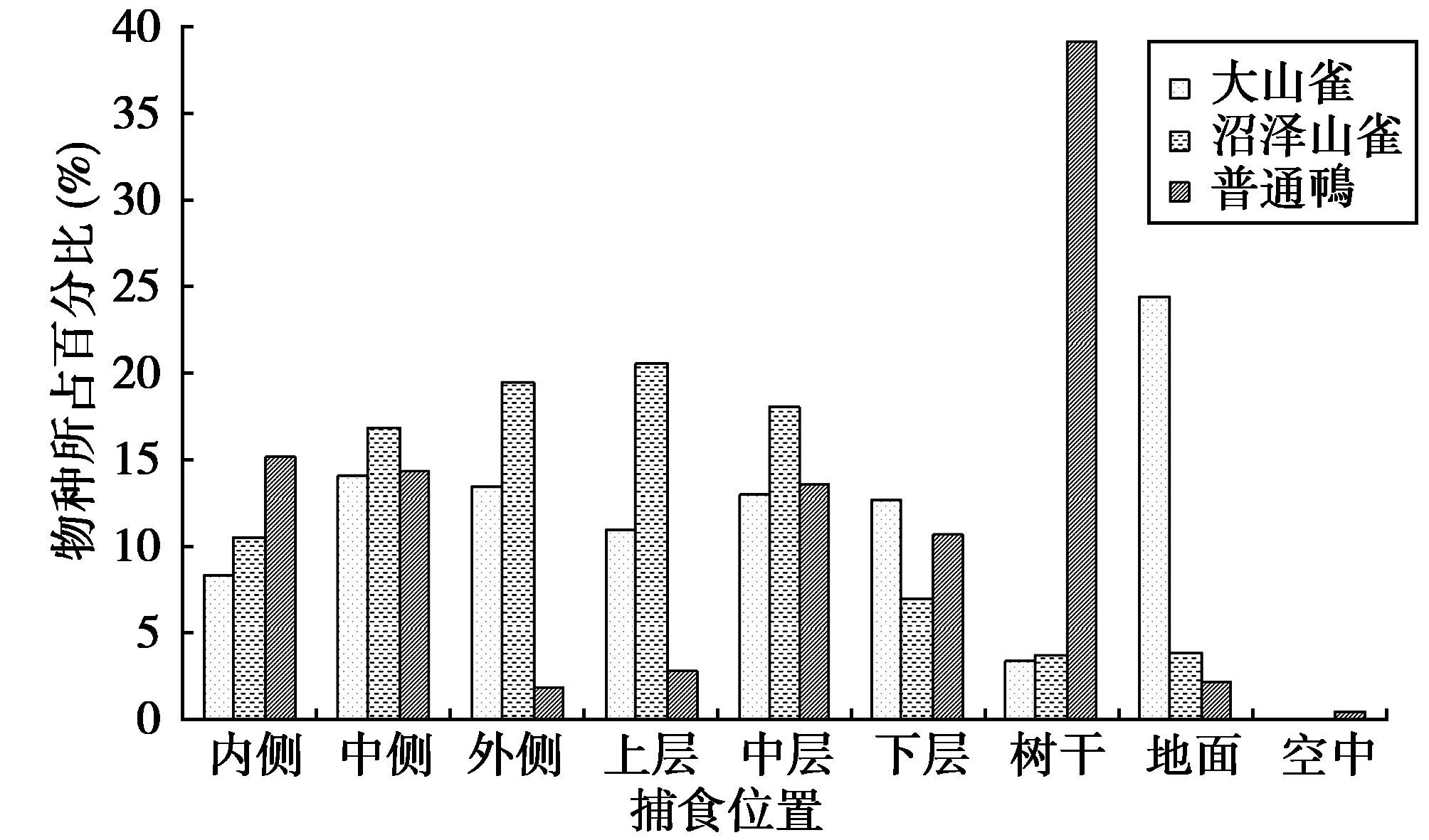
图1 混群中核心种的捕食位置Fig.1 Predation position of core specie in mixed-species flocks
3.5 核心种生态位的相互关系
3.5.1 核心种群的生态位宽度
调查结果表明,在9个捕食位置中,大山雀的生态位宽度值最大,取食位置较广;普通鳾生态位宽度值最小,取食位置比较单一(表7)。单从生态位宽度来看,大山雀的生态适应能力相比于普通鳾较强。
表7 核心种的生态位宽度
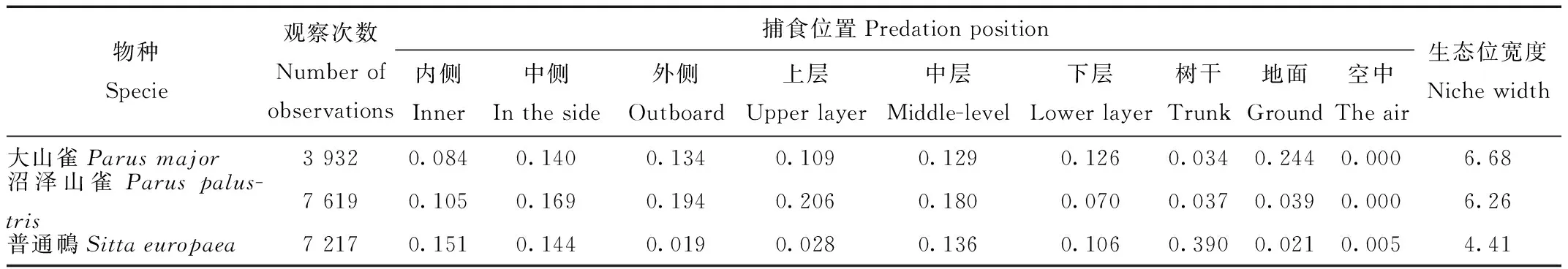
Tab.7 The niche width of core specie
3.5.2 核心种的生态位重叠
运用Hulbert生态位重叠计算[25],结果表明,大山雀与沼泽山雀生态位重叠比较大,取食位置相近;普通鳾与大山雀、沼泽山雀的生态位重叠都较小(表8)。
表8 核心种之间的捕食位置生态位重叠

Tab.8 The niche overlap for predation position in core species
4 讨论
4.1 核心种在混合群形成中的作用
4.2 核心种在混合群内部竞争的重要性
每种鸟类都活动在各自的捕食位置中,但对捕食位置的利用各不相同。某些鸟类物种对捕食位置有一定的偏爱性,如普通鳾主要取食于树干,大山雀主要取食于地面(图1)。某些物种对捕食位置的利用存在显著差异(表6),如外侧捕食位置中沼泽山雀与普通鳾、中层捕食位置大山雀与沼泽山雀、树干捕食位置中沼泽山雀与普通鳾之间都存在显著差异。

从核心种的相互关系中可以看出,普通鳾与大山雀、沼泽山雀的关联系数和凝聚指数最强(表4,表5),普通鳾与大山雀、沼泽山雀生态位重叠相同,说明普通鳾在混合群中起着更为关键的作用。大山雀与沼泽山雀的凝聚指数和关联系数最小,生态位宽度相似而生态位重叠最大,说明大山雀和沼泽山雀在核心种群中处于次要地位。
[1] LaGory K E,LaGory M K,Meyers D M,et al.Niche relationships in wintering mixed-species flocks in western Washington[J].The Wilson Bulletin,1984,96(1):108-116.
[2] Moynihan M.Some adaptations which help to promote gregariousness[C].Proceedings of the 12th International Ornithological Congress,Helsinki,vol,1960,1958(523):541.
[3] Morse D H.Ecological aspects of some mixed-species foraging flocks of birds[J].Ecological Monographs,1970,40(1):119-168.
[4] Morse D H.Feeding behavior and predator avoidance in heterospecific groups[J].BioScience,1977,27(5):332-339.
[5] Wing L.Size of bird flocks in winter[J].The Auk.1941,58(2):188-194.
[6] Smith S M.An ecological study of winter flocks of black-capped and Chestnut-backed chickadees[J].The Wilson Bulletin,1967,79(2):200-207.
[7] Austin G T,Smith E L.Winter foraging ecology of mixed insectivorous bird flocks in oak woodland in southern Arizona[J].The Condor,1972,74(1):17-24.
[8] Powell G V N.Experimental analysis of the social value of flocking by starlings(Sturnusvulgaris)in relation to predation and foraging[J],Animal Behaviour,1974,22(2):501-505.
[9] Vine I.Risk of visual detection and pursuit by a predator and the selective advantage of flocking behaviour[J].Journal of Theoretical Biology,1971,30(2):405-422.
[10] Vine I.Detection of prey flocks by predators[J].Journal of Theoretical Biology,1973,40(2):207-210.
[11] Siegfried W R,Underhill L G.Flocking as an anti-predator strategy in doves[J].Animal Behaviour,1975,23(75):504-508.
[12] Treisman M.Predation and the evolution of gregariousness.Ⅱ.An economic model for predator-prey interaction[J].Animal Behaviour,1975,23:801-825.
[13] Hamilton W D.Geometry for the selfish herd[J].Journal of Theoretical Biology,1971,31(2):295-311.
[14] Pulliam H R.On the advantages of flocking[J].Journal of Theoretical Biology,1973,38(2):419-422.
[15] Krebs J R.Social learning and the significance of mixed-species flocks of chickadees(Parusspp.)[J].Canadian Journal of Zoology,1973,51(12):1275-1288.
[16] Krebs J R.MacRoberts M H,Cullen J M.Flocking and feeding in the great tit parus major-an experimental study[J].Ibis,1972,114(4):507-530.
[17] Caraco T.Time budgeting and group size:a theory[J].Ecology,1979,60(3):611-617.
[18] Caraco T.Time budgeting and group size:a test of theory[J].Ecology,1979,60(3):618-627.
[19] 高玮,冯贺林,相桂权,等.山地次生林冬季鸟类集群的初步研究[J].动物学报,1993,39(4):385-391.
[20] 杨志杰,高玮.小兴安岭冬季山雀类集群的初步研究[J].野生动物,1988,9(5):28-29.
[21] 高玮,杨志杰.两种山雀在人工落叶松林中冬季捕食生态位和竞争[J].东北师大学报:自然科学版,1987(4):101-106.
[22] 高玮,李万超.三种啄木鸟的生态位和竞争[J].东北师大学报:自然科学版,1997(1):78-81.
[23] Morrison M L,With K A,Timossi I C,et al.Composition and temporal variation of flocks in the Sierra Nevada[J].The Condor,1987,89(4):739-745.
[24] MacArthur R H.Geographical ecology[M].New York:Harper and Row,1972.
[25] Hurlbert S H.The measurement of niche overlap and some relatives[J].Ecology,1978,59(1):67-77.
[26] Diamond J M.Assembly of species communities[C]//Cody M L,Diamond J M.Ecology and evolution of communities.Harvard Univ.Press,1975:342-444.
[27] Ase T J,Sidell R.Pattern and chance in the structure of model and natural communities?[J].Evolution,1983,37(4):832-849.
[28] Drake J A.Communities as assembled structures:do rules govern pattern?[J].Trends in Ecology & Evolution,1990,5(5):159-164.
[29] Keddy P A.Assembly and response rules:two goals for predictive community ecology[J].Journal of Vegetation Science,1992,3(2):157-164.
[30] Tirado R,Pugnaire F I.Community structure and positive interactions in constraining environments[J].Oikos,2005,111(3):437-444.
[31] 高玮.中国东北地区鸟类及其生态学研究[M].北京:科学出版社,2006:402-407.
