麋鹿采食与非采食区群落重要值和狼尾草种群的差异
2018-03-17贾媛媛安玉亭孙大明沈华陆立林原宝东
贾媛媛安玉亭孙大明沈 华陆立林原宝东
(1.盐城市麋鹿研究所,盐城,224136;2.江苏省大丰麋鹿国家级自然保护区管理处,盐城,224136;3.商丘师范学院生物与食品学院,商丘,476000)
麋鹿(Elaphurusdavidianus),国家Ⅰ级保护动物,是适应湿地环境的大型食草动物,野生麋鹿种群的恢复对世界野生动物保护具有重要的科研与应用价值[1]。截至目前,麋鹿相关的国内外研究大致可分为历史研究[2]、食物及栖息地研究[3-10]、疾病防治[11]、种群管理[12]、行为研究[13-16]、生理生化[17]等。英国、美国等国家的野生动物专家研究较多的主要是麋鹿生活习性[18]、驯养与繁殖和栖息地恢复[19-24]等,这些研究为我们进一步了解该物种的濒危机制和生态价值提供了重要的基础资料。
江苏大丰麋鹿国家级自然保护区自1986年成立以来,麋鹿数量从39头递增为2016年的3 223头,种群数量和密度均有大幅度增加,高密度的麋鹿的采食、代谢废物以及践踏作用会影响植被的更新[10],保护区的植被群落[25]也随鹿群增多在不断演替,并呈现出较为明显的带状分布,麋鹿采食区与非采食区植被群落差异明显。只有适应力强的植物才能够更新,狼尾草(Pennisetumalopecuroides)是麋鹿采食区和非采食区内少数能更新的植物,基于这一现实问题,在2015年和2006年6月对麋鹿采食与非采食区群落重要值和麋鹿主要食物狼尾草种群的差异进行研究,旨在揭示麋鹿采食对栖息地的影响,研究结果可为进一步保护麋鹿及其栖息地植被群落提供基础信息。
1 研究区概况
大丰麋鹿保护区位于江苏省中部的南黄海湿地(E 120°47′~120°53′,N 32°59′~33°03′),保护区内生物多样性丰富,原生态以草本种类居多。在第一放养区内,由于麋鹿的种群密度过高,造成植物群落更新缓慢,只有生长速度快、耐旱、抗倒伏、无病虫害、适应力较强狼尾草能够更新,狼尾草3月开始发芽,6月是生长的茂盛期,由于采食区其他植被更新困难,麋鹿采食植物主要以狼尾草为主[26]。当地气候以潮湿、多雨、多风为主,无霜期217 d,年均降雨量1 068 mm,年均气温14.1℃,1月平均气温-0.8℃,7月平均气温26.8℃。地势平坦,海拔2~4 m,土壤偏碱性,pH 7.7~8.4,0~40 cm土壤中NaCl含量为0.04%~1.13%[26]。经过30 a(1986~2016年)的有效保护,已形成846头的野外麋鹿种群。随着科研保护力度的不断加大,大丰麋鹿种群得到有效保护的同时,鸟类等其他生物也得到了保护和发展[27]。
2 研究方法
根据狼尾草的季节性生长特性和麋鹿对狼尾草的适口性喜好程度[28],分别于2015年、2016年的6月下旬,调查麋鹿主要生活区狼尾草的密度、高度、盖度和地上干重;并在麋鹿主要放养区代表性地段选定4个样点,即东区近水源草地、西区近水源草地、东区林下、西区林下,对其植物种类、盖度、密度、频度、各物种的平均高度等参数进行调查。采取随机“五点样方法”,先选取20 m×20 m的大样方,然后在每个大样方的4个角落和样方正中心选取1 m×1 m的小样方进行调查,并收集数据。每项调查重复30次。有关计算方法如下:
(1)物种重要值(IV):IV=[相对频度(RF)+相对盖度(RC)+相对密度(RD)]/3
(2)多样性指数:Shannon指数:H=-H=-∑Piln(Pi)
式中:H为Shannon指数;Pi为第i种植物的重要值占所有植物重要值总和的比值。计算公式表明,群落中生物种类增多代表了群落的复杂程度增高,即H值愈大,群落所含的信息量愈大。
(3)均匀度指数:J=H/lnS
式中:J为均匀度指数;S为群落中的植物种类数。
(4)生态优势度:C=∑(ni/Ns)2
式中:C为生态优势度;S为群落中植物种类数;Ns为群落中所有种的重要值总和;ni为第i种植物的重要值。
指标变量是否符合正态分布,本研究首先对各指标变量做了Kolmogorov-Smirnov正态分布检验,显著水平选α=0.05。对于不符合正态分布的数据进行比较时,采用非参数统计学检验方法的Mann-WhitneyU检验法。符合正态分布的数据采用Studentt检验法。显著性差性水平采用3个水平即显著(α=0.05)、非常显著(α=0.01)和极显著(α=0.001)。所有统计学检验采用SPSS 21.0完成。
3 结果与分析
3.1 植被群落结构调查
本研究主要调查位于老海堤西南侧的第一放养区,属于较早成陆地带,植物多样性相对丰富。该区分为东区和西区,东区为核心区;西区为实验区,开展生态旅游。
表1 各样点植被群落重要值
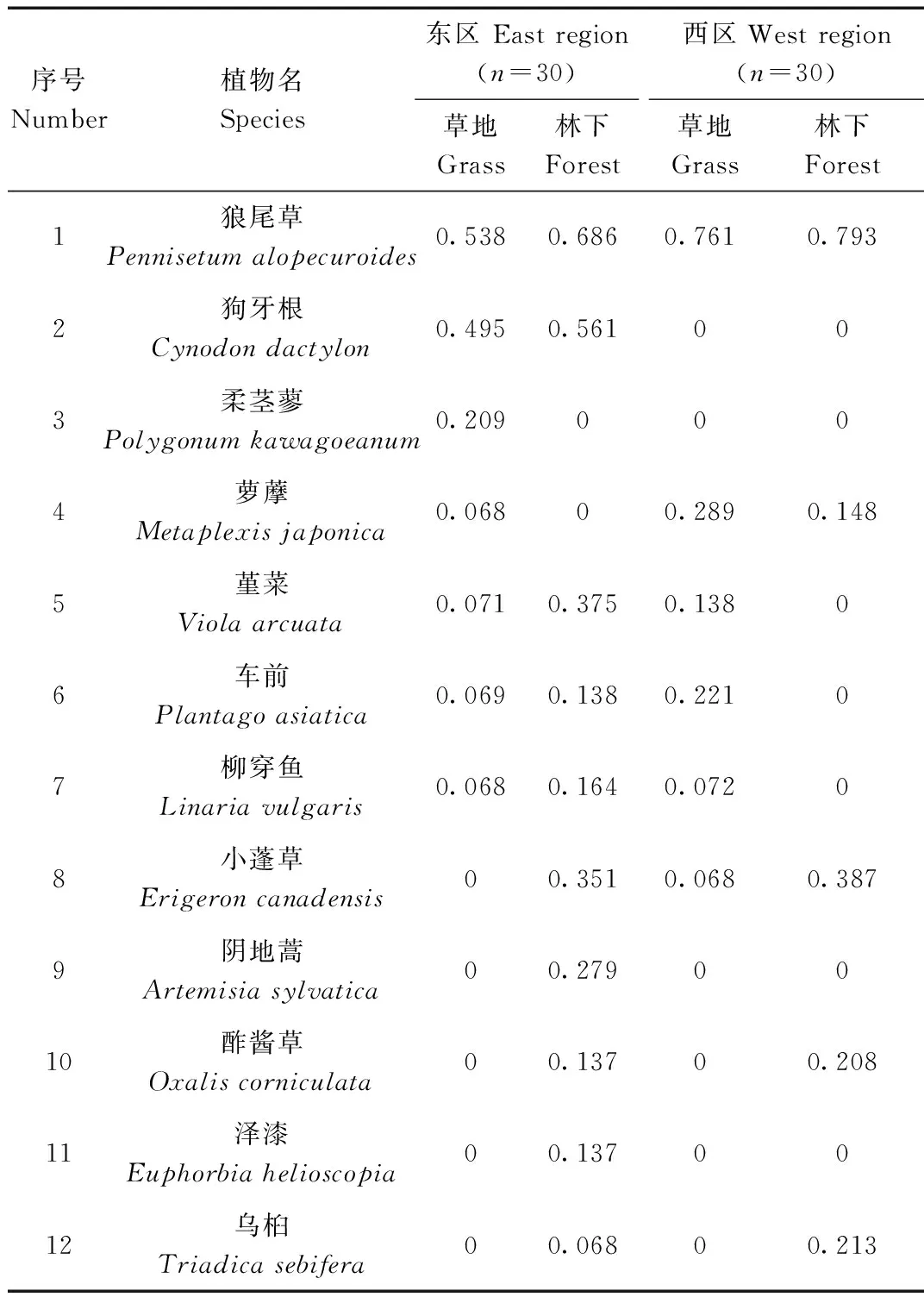
Tab.1 Important values in different survey spots
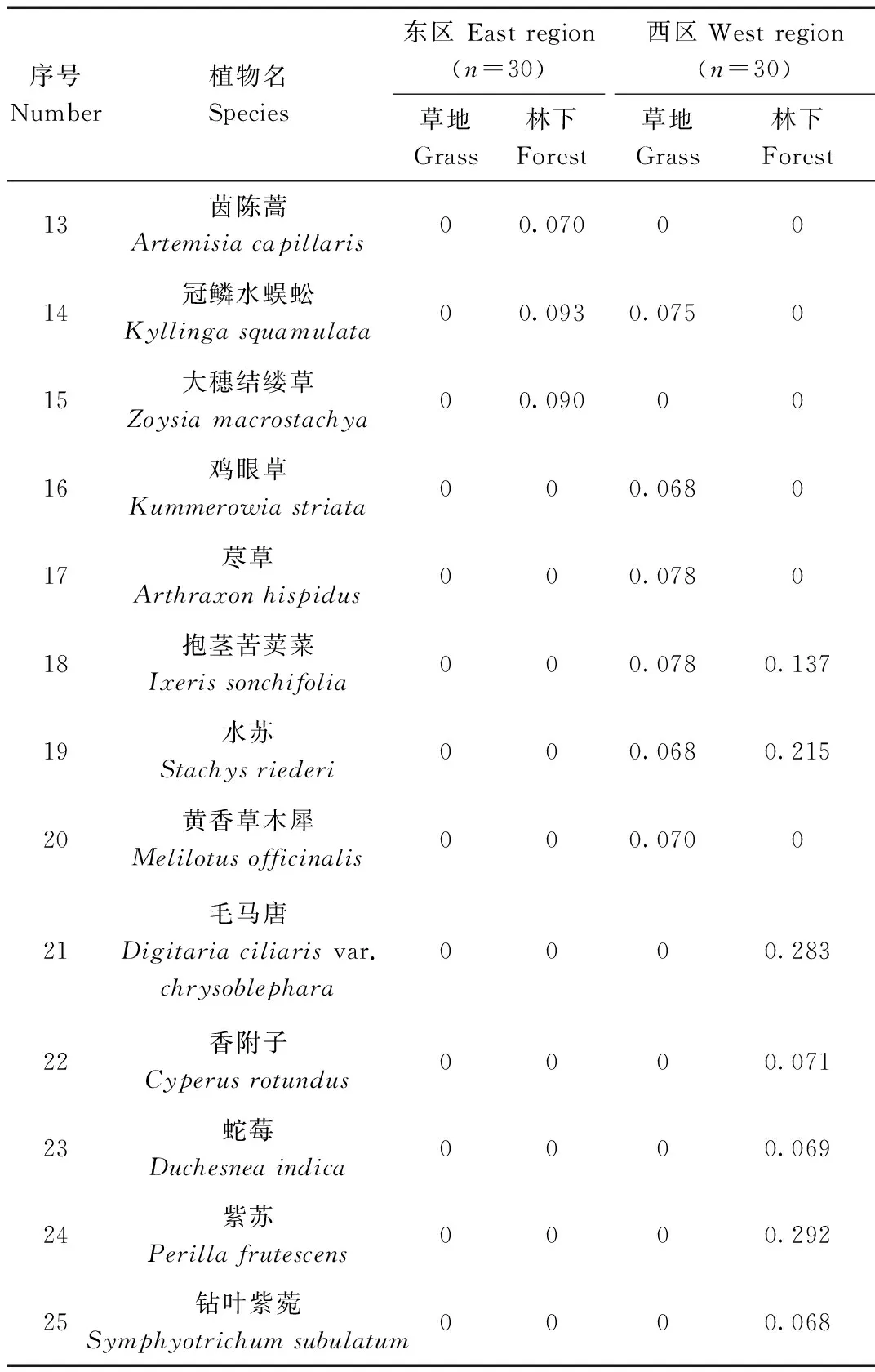
续表1
上述共调查到25种植物,近水源草地调查到14种,林下调查到21种,其中狼尾草重要值最高,属于绝对优势物种。其他植物物种的重要值都处于较低水平。
表2 各样点植被群落特征
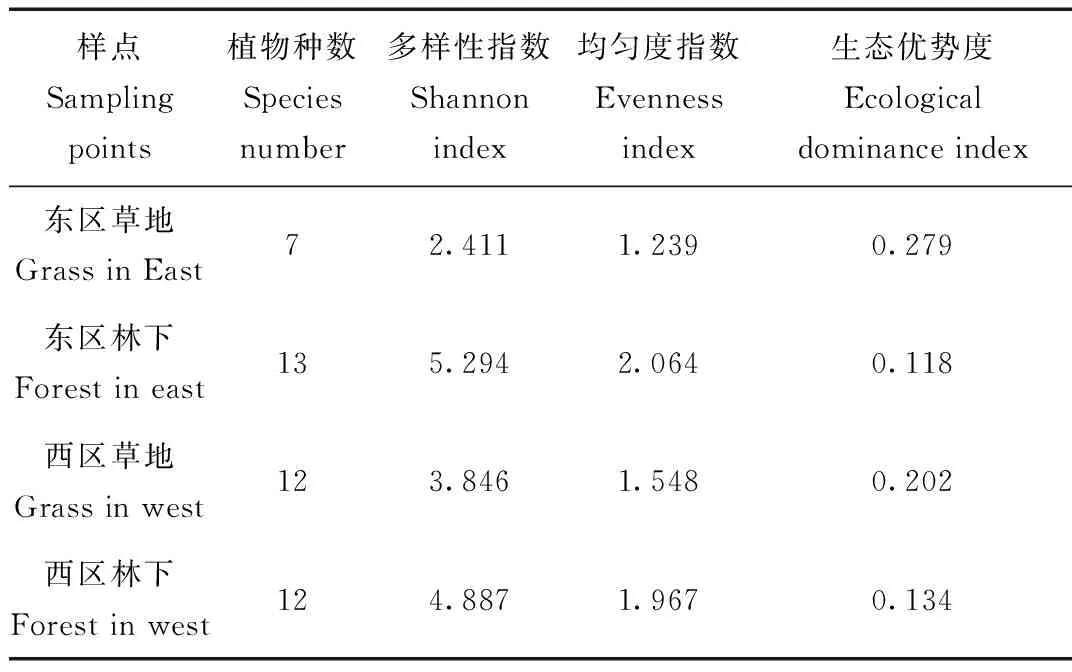
Tab.2 Vegetation characteristics of all sampling points
通过东西区植被群落的基本特征对比(表2)得出,麋鹿活动较少的林下多样性指数大于活动较多的近水源草地(东区林下多样性指数(EHF)=5.294,东区草地多样性指数(WHW)=4.887;西区林下多样性指数(EHF)=2.411,西区草地多样性指数(WHW)=3.846);均匀度指数林下大于近水源草地(东区林下均匀度指数(EJF)=2.064,东区草地均匀度指数(WJW)=1.967;西区林下均匀度指数(EJF)=1.239,西区草地均匀度指数(WJW)=1.548),而生态优势度指数小于近水源草地(东区林下生态优势度指数(ECF)=20.118,东区草地生态优势度指数(WCW)=20.134;西区林下生态优势度指数(ECF)=20.279,西区草地生态优势度指数(WCW)=20.202)。
3.2 麋鹿主要采食植物样方调查
主要是测定1 m2样方内狼尾草的生物量,将样方中地上部分进行平地刈割称重,研究其地上生物量。
表3 采食区与非采食区的狼尾草参数对比

Tab.3 Parameters’ comparison of Pennisetum alopecuroides (L.)Spreng.in the feeding area and non-feeding area
调查发现狼尾草是麋鹿优先采食的植物,利用Mann-Whitney-U对非采食区和采食区狼尾草的各参数值进行检验(表3),非采食区的密度、盖度、均高、地上干重分别为13.30株/m2、94.80%、146.46 cm、2 443.97 g/m2;采食区的密度、盖度、均高、地上干重分别为6.77株/m2、25.30%、16.21 cm、243.07 g/m2,非采食区参数值分别是采食区的1.99倍、3.75倍、9.04倍和10.06倍。利用Mann-Whitney-U对非采食区和采食区狼尾草的密度、盖度、均高、地上干重进行检验,都具有显著性差异(Z=14.717,P=0.001;Z=-70.500,P=0.001;Z=-49.505,P=0.001;Z=-17.839,P=0.001)。
4 讨论
4.1 群落特征分析
林下重要值高于或者接近于近水源草地(表1),麋鹿属于平原湿地动物[1],角枝庞大,密集的树林对麋鹿觅食与行动造成障碍;其次,近水源草地既能满足麋鹿的采食,又便于麋鹿的饮水,因此麋鹿更偏向于在近水源草地觅食。同一植物物种的重要值西区基本上均高于东区,原因分析为麋鹿天性胆小[26],西区游客及观光车的来来往往导致该调查范围内麋鹿白天觅食减少,偏向于在无干扰的地段和夜间采食;而东区为核心区,无人为干扰,麋鹿全天候都可以自由采食。
林下植被群落的多样性指数和均匀度指数均大于近水源草地的植被群落(表2),表明林下的植物物种数量更多,且每物种的个体分布更均匀。根据生态优势度统计结果,东西两区近水源草地明显大于林下植被群落,表明草地群落内物种数量分布非常不均匀,狼尾草的优势种的地位突出。总的来看,东区林下的植被多样性、个体数量的均匀度较高,植被群落质量较高。可能由于东区为核心区,西区为实验区并已开发为旅游景区,频繁的人为活动干扰是造成东西区植被群落分布差异的原因之一[29]。
4.2 采食区狼尾草差异性分析
植物资源的种类、植被的生长情况等生境因子与麋鹿的生存关系密切[29]。本研究发现,麋鹿采食区与非采食区狼尾草在密度、盖度、高度和地上部分干重几个指标上P<0.001(表3),差异极显著,此种现象可能由于麋鹿的采食行为等对其栖息地植物的生长产生了影响。麋鹿春夏季喜食植物的叶茎,秋季常采食植物的花穗籽粒,冬季善采食植物的根[26]。狼尾草为调查区麋鹿的主要采食植物,因为调查期正值夏初,狼尾草叶嫩,含水量高,木质化程度低,适口性好,遭到麋鹿的过度采食。据测算,每头麋鹿大概需要2.6 hm2的肥沃土地,才能够满足自身的觅食和栖息需要[30],保护区总面积为2 667 hm2,截至2016年底,大丰麋鹿种群为3 223头,每头麋鹿平均土地拥有量为0.8 hm2,仅占理论占有值的30.8%,因此,采食力度也扩大到理论值的3.25倍。采食压力的加大造成麋鹿采食区狼尾草的生物量远小于未采食区。其次,由于麋鹿种群密度过高,对栖息地的踩踏力度很大,湿地上麋鹿采食的地方都能够看到明显的足迹。践踏极易造成土壤板结,土壤孔隙度减小,抑制生命力弱的植物根部的呼吸作用,影响采食区植被的更新。这一结果与湖北石首麋鹿种群密度过高会影响植被更新相类似[10]。麋鹿产生的代谢废物,如粪便、尿液等,适量的代谢物能够提高土壤有机质含量,改善土壤肥力;但是由于种群密度过高容易导致土壤pH提高[31],因此土壤粪尿会间接导致大丰麋鹿栖息地土壤盐渍化程度加剧,同时麋鹿大规模、高密度的集群活动造成土壤板结的同时,也降低了土壤有机质的含量。即使土壤未板结地段,大量的麋鹿排泄物经微生物分解后,也容易造成土壤肥力陡增,出现栖息地植物“烧苗”现象。
因此,为缓解麋鹿种群快速增长和栖息地急剧退化的矛盾,可以采取一些措施,如:扩大保护范围、轮牧放养、人工投喂、异地野放、合理开发等,以此来降低麋鹿的种群密度,促进麋鹿种群健康可持续发展。
致谢:本研究野外调查工作和论文写作过程中得到了江苏省大丰麋鹿国家级自然保护区诸多同事的热心帮助,在此一并致以诚挚的谢意。
[1] 曹克清,陈彬.关于野生麋鹿绝灭原因的再探讨[J].四川动物,1990,9(1):41-42.
[2] 白加德,张林源,钟震宇,等.中国麋鹿种群发展现状及其研究进展[J].中国畜牧兽医,2012,39(11):225-230.
[3] 李钢铁,秦富仓,贾守义,等.旱生灌木生物量预测模型的研究[J].内蒙古林学院学报:自然科学版,1998,20(2):25-31.
[4] 孟玉萍,李坤,张林源,等.北京麋鹿苑麋鹿采食量测定分析[J].特产研究,2010,32(4):39-42.
[5] 何振,杨道德,马建章,等.湖北石首麋鹿的冬季生境选择[J].四川动物,2007,26(4):764-768.
[6] 赵小雷,凌云,张光富,等.大丰麋鹿保护区不同生境梯度下滩涂湿地植被的群落特征[J].生态学杂志,2010,29(2):244-249.
[7] 陈星,张林源,钟震宇,等.湖北麋鹿保护区麋鹿产仔期植被分布及种群概况[J].四川动物,2011,30(5):820-824.
[8] 丁宁宁,王保松,梁珍海,等.江苏大丰麋鹿保护区不同改良措施对滩涂土壤的改良效应研究[J].土壤,2011,43(3):487-492.
[9] 纪一帆,吴宝镭,丁玉华,等.大丰野放麋鹿生境中芦苇和互花米草的营养对比分析[J].生态学杂志,2011,30(10):2240-2244.
[10] 李鹏飞,温华军,沙平,等.石首麋鹿国家级自然保护区湿地生境退化与保护对策[J].绿色科技,2012(6):249-251.
[11] 钟震宇,张林源,唐宝田.2000-2009年北京南海子麋鹿出血性肠炎发生特点分析[J].畜牧与兽医,2013,45(4):56-58.
[12] 周宇虹,黄佳怡.大丰自然保护区麋鹿种群密度制约增长的模型[J].南京林业大学学报:自然科学版,2013,37(5):172-174.
[13] 蔡桂全,谢家华,Mathur N.麋鹿发情期主要活动的时间分配及行为研究[J].兽类学报,1988,8(3):166-171.
[14] 蒋志刚.麋鹿行为谱及PAE编码系统[J].兽类学报,2000,20(1):1-12.
[15] 曾岩,蒋志刚,李春旺,等.麋鹿幼仔的活动同步性与同性聚群倾向[J].兽类学报,2004,24(1):78-81.
[16] 程志斌,张林源,刘艳菊,等.北京南海子麋鹿体型的性二型及生长发育[J].四川动物,2015,34(3):330-337.
[17] 段艳芳,陈付英,李玉峰,等.麋鹿雌性生殖系统的组织学观察[J].河南农业科学,2012,41(8):180-184.
[18] Sargeant G A,Eberhardt L E,Peek J M.Thermoregulation by mule deer (Odocoileushemionus) in arid rangelands of southcentral Washington[J].Journal of Mammalogy,1994,75(2):536-544.
[19] Palmer S C F,Truscott A M.Seasonal habitat use and browsing by deer in Caledonian pinewoods[J].Forest Ecology and Management,2003,174(1/3):149-166.
[20] Linnell J D C,Nilsen E B,Andersen R.Selection of bed-sites by roe deerCapreoluscapreolusfawns in an agricultural landscape[J].Acta Theriologica,2004,49(1):103-111.
[21] Adrados C,Baltzinger C,Janeau G,et al.Red deerCervuselaphusresting place characteristics obtained from differential GPS data in a forest habitat[J].European Journal of Wildlife Research,2008,54(3):487-494.
[22] Borkowski J,Ukalska J.Winter habitat use by red and roe deer in pine-dominated forest[J].Forest Ecology and Management,2008,255(3/4):468-475.
[23] Brodie J F,Brockelman W Y.Bed site selection of red muntjac (Muntiacusmuntjak) and sambar (Rusaunicolor) in a tropical seasonal forest[J].Ecological Research,2009,24(6):1251-1256.
[24] Torres R T,Carvalho J C,Panzacchi M,et al.Comparative use of forest habitats by roe deer and moose in a human-modified landscape in southeastern Norway during winter[J].Ecological Research,2011,26(4):781-789.
[25] 梁崇岐,李渤生.我国半散放麋鹿生境植被及采食植物种类的研究[J].林业科学,1991,27(4):425-434.
[26] 蒋志刚,丁玉华.大丰麋鹿与生物多样性[M].北京:中国林业出版社,2011.
[27] 刘彬,丁玉华,任义军,等.大丰麋鹿国家级自然保护区鸟类多样性[J].野生动物,2012,33(1):11-17.
[28] 张国斌,薛建辉,吴永波.半圈养状态下麋鹿对生境的影响[J].中国农学通报,2007,23(7):180-184.
[29] 刘金根,薛建辉,王磊,等.江苏大丰麋鹿自然保护区栖息地退化特征[J].生态学杂志,2011,30(8):1793-1798.
[30] 徐殿波,丁玉华.大丰麋鹿保护区生境改造的探讨[J].农村生态环境,1997,13(1):10-12.
[31] 朱明淏,刘艳菊,张婷婷,等.不同栖息地环境下麋鹿活动对土壤理化特性的影响[J].环境化学,2016,35(1):208-217.
