温度对江南油杉种子萌发特性的影响
2018-03-12,,,,,
,,,, ,
(1.广西壮族自治区林业科学研究院广西优良用材林资源培育重点实验室,南宁 530002;2.中南林业科技大学生命科学与技术学院,湖南 长沙 410004)
种子繁殖是植物繁衍的重要手段,种子萌发潜在影响着植物种群的动态和组成,任何不利于种子萌发的因素都会直接影响到种群的稳定性[1]。温度是植物种子萌发的主要因素,温度高低直接影响种子萌发和出苗,适宜的温度是种子萌发所需的必要条件,掌握适宜种子萌发的温度对指导苗木生产实践具有重要意义[2-3]。
江南油杉(Keteleeriafortuneivar.cyclolepis)隶属松科(Pinaceae)油杉属(Keteleeria),是我国特有常绿大乔木,现块状分布或散生于我国广西、云南、贵州、广东、湖南、福建、浙江、江西等省区海拔1 000 m以下的丘陵和低山,球果圆柱形或椭圆状圆柱形,长7~15 cm,径3~6 cm,种子椭圆形,正面褐色有光泽,背面淡黄色,10月中旬至11月中旬成熟[4]。江南油杉具有很强的抗逆性和生态适应性,加之其树干高大通直,材质优良,是山地造林、用材林和园林绿化的理想树种。由于长期人为砍伐和自生自灭过程,江南油杉天然种质资源流失严重,现处于渐危状态[5]。据调查,江南油杉林下幼苗严重缺乏,种群更新困难,在100 m2范围内,株高<20 cm的幼苗仅有6~10株。

表1 江南油杉种子大小和采集地基本信息
注:LY为广西凌云县;BB为广西博白县;FN为云南富宁县;WM为贵州望谟县。
目前,对江南油杉的研究主要集中在系统分类[6],地理分布[7-8],群落结构特征[8]、细胞生物学[9]、育苗技术[10]等方面,迄今未见有关江南油杉种子萌发特性的研究报道。为此,笔者设置温度梯度开展种子萌发实验,以了解温度变化对江南油杉种子萌发的影响,揭示江南油杉不同种源间种子萌发对温度响应的差异以及与生境条件的关系,并探明江南油杉种子最佳发芽温度,为江南油杉资源开展合理保护以及引种栽培提供参考。
1 材料与方法
1.1 供试种子
实验材料分别来源于广西博白县、凌云县、云南富宁县、贵州望谟县4个江南油杉天然种群,其中广西为江南油杉中心分布区,云南和贵州为边缘分布区。采集地基本信息如表1所示。果实采收于2014年10月,球果采回后堆放在室内数天,待种子充分成熟后摊晒于阳光下,种鳞自然开裂脱出种子,种子晾干后,每个种群随机挑选30粒种子,测定种子长、种子宽和种子厚,重复3次;每个种群随机挑选100粒饱满种子用电子天平称重,重复5组,计算种子千粒重(表1)。
1.2 种子萌发
实验前待试种子用0.2%K2MnO4溶液消毒10 min,再用蒸馏水冲洗干净,之后置于蒸馏水中浸泡24 h催芽。种子催芽后每个种群随机选取30粒种子均匀放入铺有2层滤纸的10 cm培养皿内,每种群重复3次。每个培养皿内加入适量蒸馏水,以不使种子移动为适。实验共设置5个恒温(15,20,25,30,35 ℃)和2个变温(15 ℃/25 ℃、20 ℃/30 ℃)(12/12 h)条件,所有萌发处理在1 000 lx,12 h/d的光照,90%RH下进行。实验中当胚根达到种子长1/2时视为发芽,每24 h观察记录萌发状况,同时补充适量水分。实验持续30 d。
1.3 发芽指标计算公式
1) 萌发率(%)=发芽种子总数/供试种子总数×100%;
2) 发芽势(%)=10 d内正常发芽种子数/供试种子总数×100%;
3) 发芽指数=∑(Gt/Dt),式中,Gt为t天内的发芽种子数,Dt为发芽天数;
4) 萌发时滞(d):第1粒种子萌发所需天数;
5) 萌发历期(d):第1粒种子和最后1粒种子萌发时间差;
6) 胚芽和胚根长度(mm):种子播种的第20天,从每个培养皿选取10粒萌发的种子分别测定胚芽和胚根长度(不足10粒则全部测量);

注:同一种源不同字母表示差异显著(p<0.05)。图1 温度对不同种源江南油杉种子萌发指标的影响
7) 萌发值=[(∑DGS)/N]×Gr×10,DGS为日累积萌发数除以相应的萌发天数所得商,N为从萌发开始到结束的天数,Gr为最终萌发率[11-13]。
1.4 数据处理
用Excel 2007进行常规数据统计以及作图;用SPSS 19.0对不同种源和不同温度条件下江南油杉种子萌发指标进行方差分析,多重比较(采用邓肯氏新复极差法),通过相关分析分析江南油杉种子萌发指标与种子形态、大小以及种源地环境因子的相关性,显著性水平均设定为α=0.05。
2 结果与分析
2.1 温度对江南油杉种子萌发率、发芽势和发芽指数的影响
不同温度条件下4个种源江南油杉种子萌发率、发芽势和发芽指数见图1。恒温条件下,随着温度升高,4个种源种子的萌发率呈先增加后减小的趋势。25 ℃条件下,4个种源种子萌发率均达到最大值,分别为富宁53.3%、博白70.0%、望谟60.0%、凌云48.3%。富宁和望谟种源均在15 ℃条件下萌发率最低,而博白和凌云种源均在35 ℃条件下萌发率最低。变温条件下,除了在20 ℃/30 ℃下富宁的种子萌发率略高于30 ℃,其余种源种子在20 ℃/30 ℃下的萌发率均低于30 ℃;4个种源在15 ℃/25 ℃下的萌发率均低于25 ℃。多重比较结果表明,4个种源江南油杉种子在不同温度条件下的萌发率部分结果存在显著差异。
恒温条件下,江南油杉种子的发芽势和萌发率一样,随着温度升高其发芽势呈先增加后减小的趋势,博白和凌云种源种子的发芽势随着温度升高,在25 ℃下达到最大值,而富宁和望谟种源种子的发芽势随着温度升高,在30 ℃下达到最大值,凌云种源种子在15,20,35 ℃下发芽势均为0。变温条件下,博白种子在20 ℃/30 ℃下发芽势大于30 ℃下发芽势,凌云种子在20 ℃/30 ℃下发芽势与30 ℃下发芽势相等。
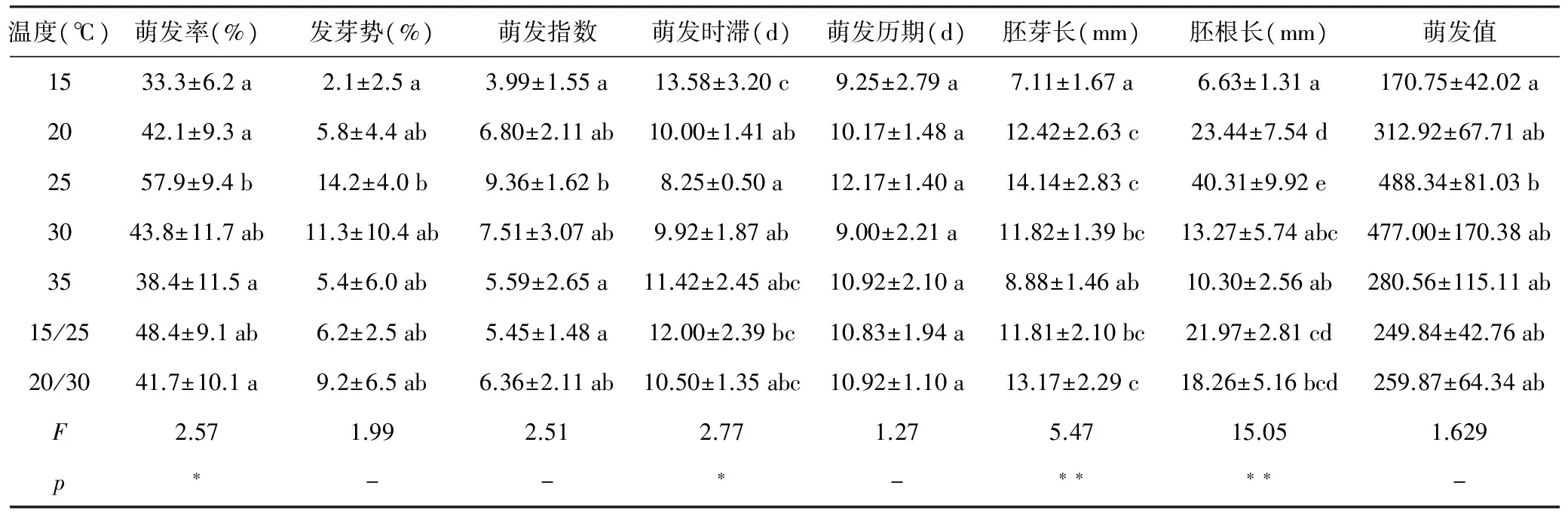
表2 不同温度条件下江南油杉种子萌发指标的差异
注:同一列数据不同字母表示差异显著;“-”表示差异不显著,“*”p<0.05;“**”p<0.01。
4个种源江南油杉种子发芽指数各不相同,表现为随着温度升高发芽指数呈先增加后减小的趋势。富宁、博白、凌云种源种子均在25 ℃条件下发芽指数达到最大值;望谟种源种子在35 ℃条件下发芽指数达到最大值。富宁和望谟种子在15 ℃下发芽指数最小;博白和凌云种子在35 ℃下发芽指数最小。变温条件下,博白种子在20 ℃/30 ℃下发芽指数大于30 ℃。
2.2 温度对江南油杉种子萌发时间、胚芽和胚根生长的影响
不同温度条件下,江南油杉种子萌发时间、胚芽和胚根生长见图1。4个种源江南油杉种子在15 ℃条件下,萌发时滞均最长,分别为富宁10.33 d、博白11.33 d、望谟16.33 d、凌云16.33 d。25 ℃条件下,4个种源江南油杉种子萌发时滞均最短,且25 ℃下各种源种子萌发时滞均显著小于15 ℃下种子萌发时滞。温度对各种源种子萌发历期的影响无明显规律,表现为:富宁种源在15 ℃条件下萌发历期最长,在35 ℃下最短;望谟和凌云种源在15 ℃下萌发历期最短,在25,30 ℃下最长;博白种源在35 ℃下萌发历期最长,在30 ℃下最短,为6.00 d。
恒温条件下,4个种源江南油杉种子胚芽和胚根长度均随着温度升高呈先增加后减小的趋势,均在15 ℃条件下最小,在25 ℃条件下达到最大值,且25 ℃下各种源种子胚根长显著大于其它温度下各种源种子胚根长。4个种源中,富宁种源在15 ℃下胚芽和胚根长均最短,凌云种源在25 ℃下胚芽长最长,望谟种源在25 ℃下胚根长最长。变温条件对江南油杉种子萌发时间、胚芽和胚根生长均无明显促进作用(图1中上部分为胚根长,下部分为胚芽长)。
2.3 不同温度条件下江南油杉种子萌发指标差异
由表2可知,江南油杉种子的萌发率、萌发时滞、胚芽长、胚根长在不同温度条件下存在显著或极显著差异;发芽势、萌发指数、萌发历期、萌发值在不同温度条件下差异不显著。25 ℃条件下,江南油杉种子萌发率、发芽势、萌发指数、萌发历期、胚芽长、胚根长以及萌发值均达到最大值,萌发时滞值最小。15 ℃条件下,江南油杉萌发率、发芽势、萌发指数、胚芽长、胚根长和萌发值均最小,萌发时滞最大。30 ℃条件下,江南油杉种子萌发历期最小。以上结果说明,江南油杉种子最适宜在25 ℃条件下萌发和生长,温度过低(15~20 ℃)种子萌发和生长缓慢,且萌发率低,温度过高(30~35 ℃)则会抑制种子萌发和生长。
2.4 江南油杉萌发指标与种子大小及环境因子的相关性
以每个江南油杉种群在所有温度条件下种子各萌发指标的平均值与种子大小、环境因子进行相关性分析,结果见表3。由表3可知,江南油杉种子的萌发值与千粒重呈显著正相关,与种子宽呈显著负相关。种子萌发的胚芽长度与千粒重呈显著负相关。其它萌发指标与种子大小、环境因子无显著相关性。
2.5 温度与萌发值的函数关系
相关研究表明,萌发值是较为理想的检验种子萌发能力的指数,可以综合种子的发芽率、萌发速率和萌发时间[11-13]。以温度为自变量,萌发值为因变量进行回归分析,确定温度与萌发值在[15,35]区间内的函数关系。
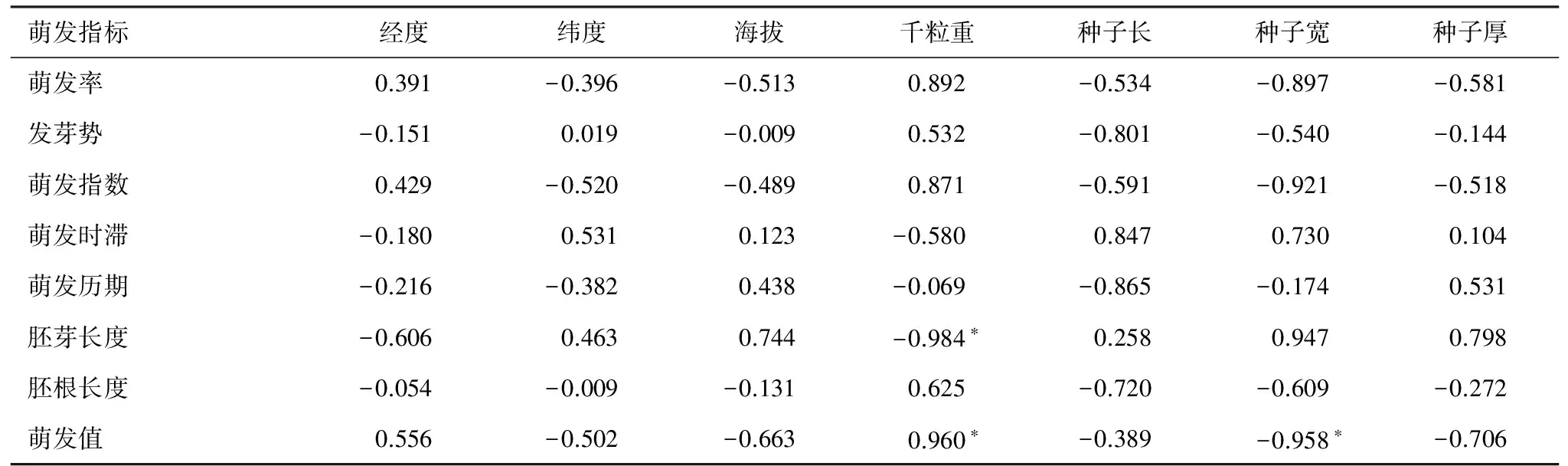
表3 江南油杉萌发指标与种子大小、环境因子的相关性
通过回归分析得出温度与萌发值的回归模型为:
y=0.014 778x4-1.623 380x3+62.231 223x2-971.303 008x+5 469.041 822,R2=1。
利用函数,分别代入原点数据检测函数准确性,结果见表4。

表4 拟合模型实测值与预测值间的比较
由表4可知,函数对种子萌发值的预测值与实测值相差不大,函数拟合效果较好,能准确反映江南油杉种子萌发温度与萌发值的关系。通过函数模型计算,在15~35 ℃范围内,当温度达到27.3 ℃时,江南油杉种子萌发值达到最大值(511.370 55)。因此,预测江南油杉种子最适萌发温度为27.3 ℃。
3 讨 论
温度是植物种子萌发的必要条件,不同种子的最适萌发温度不同[14-15]。本研究结果表明,温度变化对江南油杉种子萌发有重要影响。7个温度条件中,江南油杉种子在25 ℃条件下萌发率、发芽势、发芽指数、胚芽长、胚根长和萌发值均最大,这说明种子在25 ℃条件下出苗快且整齐,苗木质量高,这可能是因为在25 ℃下,种子吸水、种皮软化得到促进,且酶促反应速度加快,呼吸作用加强,为种子萌发和胚的生长提供足够能量和物质基础[16-17]。15 ℃条件下,江南油杉种子萌发率、发芽势、萌发指数、胚芽长和胚根长均最小,可能是因为种子积温不足,不能在一定时间里积累到种子萌发所需的热量,种子相继出现活力降低、腐烂和发霉等现象[18]。当温度升高到30 ℃和35 ℃时江南油杉种子萌发率、发芽势、萌发指数等指标明显降低,可能是由于温度过高影响种子内部酶变性、膜的透性以及膜结合的活性所致[19]。
相关研究表明,变温条件能显著提高某些植物种子萌发,因为高低温互变可促进种子内部各类酶系统的生理活动,致使种皮收缩,促进种子与外界气体交换[20-21]。4个种源江南油杉种子中只有少部分萌发指标显示变温条件下比恒温条件下萌发效果好,如富宁的种子在20 ℃/30 ℃下萌发率略高于20 ℃和30 ℃;博白的种子在20 ℃/30 ℃下发芽势大于30 ℃;凌云的种子在20 ℃/30 ℃下发芽势与30 ℃下相等。这说明变温对江南油杉种子萌发的影响不是绝对的,依种源而异。相对而言,恒温比变温更适合江南油杉种子萌发,说明在野外,埋藏于地表层的种子比裸露于地面的种子易萌发[22]。
最适萌发温度一直都是研究植物种子萌发的重点,确定物种种子最适萌发温度不仅可大大提高种子萌发率,而且对苗木的生长发育具有重要作用[23]。Kurtar[24]通过建立数学模型对一些瓜类植物种子最适萌发温度、萌发率和萌发速率进行了精确预测,该模型有助于模拟植物生长发育。本实验以综合种子萌发率、萌发时间和萌发速率的萌发值为因变量,温度为变量建立相关模型,通过模型预测江南油杉种子最适萌发温度,这不仅为提高江南油杉种子萌发率,促进萌发效果提供理论依据,而且对提高江南油杉苗木出苗率和壮苗率,充分利用江南油杉种质资源方面具有重要作用。
物种间种子大小与其萌发有着紧密的相关性[25-26]。江南油杉种子萌发值与千粒重呈显著正相关,种子越重,萌发值越大,其萌发效果越好。这可能一方面是由于大种子内部营养物质较多,酶活性较高,抗病性较强,在萌发时有充足养分提供[27];另一方面是因为大种子萌发时具有较强的竞争力和抵御风险的能力,而小种子的质量较低,死亡风险较高[28-29]。总体来说,江南油杉种子萌发率偏低,萌发效果不佳,这可能是其种群更新困难的重要原因,此外,种子传播范围狭小、高虫害率、高空瘪率以及幼苗高死亡率也在一定程度上影响种群更新。
[1]Jusaitis M,Polomka L,Sorensen B.Habitat specificity,seed germination and experimental translocation of the endangered herbBrachycomemuelleri(Asteraceae)[J].Biological Conservation,2004,116(2):251-266.
[2]刘小金,徐大平,杨曾奖,等.温度对越南黄花梨种子萌发的影响[J].林业科学研究,2014,27(5):707-709.
[3]Maria SO,Leticoap LG,Rosaura G.Germination of four species of the genus Mimosa,(Leguminosae) in a semi-arid zone central Mexico[J].Journal of Arid Environments,2003,55(1):75-92.
[4]牟凤娟,戴兴芬,李双智,等.油杉属植物研究动态[J].西部林业科学,2012,41(6):92-99.
[5]蒋燚,王勇,刘菲,等.江南油杉种质资源与苗木繁殖研究动态与展望[J].广西林业科学,2014,43(3):301-305.
[6]Lin J,Liang E,Farjon A.The occurrence of vertical resin canals inKeteleeria,with reference to its systematic position in Pinaceae[J].Botanical Journal of the Linnean Society,2001,134(4):567-574.
[7]王崇云,马绍宾,吕军,等.中国油杉属植物的生态地理分布与系统演化[J].广西植物,2012,32(5):612-616.
[8]何国生.福建江南油杉4种天然林群落物种结构特征[J].西南林业大学学报,2011,31(5):1-5.
[9]李林初,徐炳声.江南油杉和台湾油杉核型的比较研究[J].广西植物,1984,4(4):277-280.
[10]杨广文,杨广军,潘新军,等.江南油杉育苗技术试验[J].林业建设,2014(5):76-78.
[11]Gzabator FJ.Germination value: an index combining speed and completeness of pine seed germination[J].Forest Science,1962,8(4):386-396.
[12]杜国祯,马锦荣.十五种野生草本植物种子在不同温度下萌发能力的研究[J].草业学报,1994,3(1):18-24.
[13]闫兴富,周立彪,思彬彬,等.不同温度下PEG-6000模拟干旱对柠条锦鸡儿种子萌发的胁迫效应[J].生态学报,2016,36(7):1-8.
[14]柴胜丰,蒋运生,韦霄,等.濒危植物合柱金莲木种子萌发特性[J].生态学杂志,2010,29(2):233-237.
[15]Kumar B,Verma SK,Singh HP.Effect of temperature on seed germination parameters in Kalmegh (Andrographispaniculata,Wall.ex Nees.)[J].Industrial Crops & Products,2011,34(1):1 241-1 244.
[16]李雄,尹欣,杨时海,等.温度对高山植物紫花针茅种子萌发特性的影响[J].植物分类与资源学报,2014,36(6):698-706.
[17]徐恒恒,黎妮,刘树君,等.种子萌发及其调控的研究进展[J].作物学报,2014,40(7):1 141-1 156.
[18]张鹏,孙红阳,沈海龙.温度对经层积处理解除休眠的水曲柳种子萌发的影响[J].植物生理学通讯,2007,43(1):21-24.
[19]宋兆伟,郝丽珍,黄振英,等.光照和温度对沙芥和斧翅沙芥植物种子萌发的影响[J].生态学报,2010,30(10):2 562-2 568.
[20]Meulebrouvk K,Ameloot E,Assche JA,et al.Germination ecology of the holoparasiteCuscutaepithymum[J].Seed Science Research,2008,18(1):25-34.
[21]Qu XX,Baskin JM,Wang L,et al.Effect of cold stratification,temperature,light and salinity on seed germination and radical growth of of the desert halophyte shrub,Kalidiumcaspicum(Chenopodiaceae)[J].Plant Growth Regulation,2008,101(2):293-299.
[22]和子森,陈苏依勒,程明,等.濒危植物羽叶丁香种子休眠与萌发特性研究[J].植物生理学报,2016,52(4):560-568.
[23]Yilmaz M.Optimum germination temperature,dormancy,and viability of stored,non-dormant seeds ofMalustrilobata(Poir.)C.K.Schneid[J].Seed Science & Technology,2008,36(3):747-756.
[24]Kurtar ES.Modelling the effect of temperature on seed germination in some cucurbits[J].African Journal of Biotechnology,2010,9(9):1 343-1 353.
[25]Eriksson O.Seed size variation and its effect on germination and seedling performance in the clonal herb Convallaria majalis[J].Acta Oecologica,1999,20(1):61-66.
[26]张蕾,张春辉,吕俊平,等.青藏高原东缘31种常见杂草种子萌发特性及其与种子大小的关系[J].生态学杂志,2011,30(10):2 115-2 121.
[27]张世挺,杜国祯,陈家宽,等.不同营养条件下24种高寒草甸菊科植物种子重量对幼苗生长的影响[J].生态学报,2003,29(9):1 737-1 744.
[28]Pearson TRH,Dalling JW.Germination ecology of neotropical pioneers:Interacting effects of environmental conditions and seed size[J].Ecology,2002,83(10):2 798-2 807.
[29]王慧慧,王普昶,赵钢,等.干旱胁迫下白刺花种子大小与萌发对策[J].生态学报,2016,36(2):1-8.
(本栏目责任编辑:周介雄、戴 燚)
