低氮胁迫对烟草生物学性状、生长素及NtPINs基因家族的影响
2018-03-09王立志魏跃伟黄明月朱彦铭丁永乐贾宏昉
王立志,魏跃伟,黄明月,朱彦铭,丁永乐,贾宏昉
河南农业大学,烟草学院,烟草行业烟草栽培重点实验室,河南郑州文化路95号 450002
在所有必需营养元素中,氮素是影响烟草产量和品质形成的首要因素。根系作为植物吸收氮的主要器官,其形态和生理特性的适应性变化是植物高效利用氮的重要基础[1]。低氮胁迫下,植物根系的数量、总根长、干重、总吸收面积及活性均明显降低[2]。生长素参与了植物胁迫反应中根系构型的变化,在调节植物根系发育过程中起关键作用[3-5]。根系的生长发育与根系内部生长素浓度密切相关,玉米在高硝酸盐处理条件下,根系的生长明显受到抑制,在高硝酸盐条件下,施加外源生长素(萘乙酸和吲哚乙酸)能够促进根系的伸长[6]。生长素的浓度梯度对器官发育和组织分化有决定性的影响,生长素极性运输是导致生长素浓度产生梯度的主要原因。生长素极性运输蛋白由PINs家族基因编码,水稻、拟南芥等作物PINs蛋白家族对生长素的信号传导系统、极性运输起重要作用[7-9]。
腺毛是烟叶表面化学中重要的组织类型,是合成烟草致香物质的主要场所,对烟叶的香气质和香气量有重要影响。生长素、赤霉素、水杨酸、茉莉酸和细胞分裂素的生物合成或信号传导途径均参与腺毛的发育[10-11]。生长素参与植物腺毛的起始调控[12],外源生长素能增加植物腺毛的数量和密度[13]。
植物根系的发育对低氮胁迫的响应与体内激素水平的变化密切相关[14],有关低氮胁迫下烟草根系及腺毛研究已有报道,但多集中单一研究根系形态与激素[15-17],氮素与腺毛发育等[18-19]方向,对低氮胁迫下烟草地上部及根系构型改变与生长素分布之间的关系研究较少。利用生长素响应元件DR5融合GUS报告基因研究植物生长素的分布已有报道[20-21],但未见利用该材料研究烟草氮素的相关的报道。本试验以可标记生长素分布的DR5::GUS转基因单拷贝纯合株系D-2为材料,从生长发育、生长素分布及基因表达方面研究低氮胁迫对烟草生物学性状(根系及腺毛)、生长素及NtPINs基因家族的影响,旨在揭示生长素在低氮调控烟株发育过程中的关键作用。
1 材料与方法
1.1 试验设计
沙培试验于2016年在河南农业大学许昌科教园区的人工气候室内进行,温度昼28℃、夜24℃,光周期昼 16h、夜 8h,光强 300μmol·m-2·s-1。供试材料为响应生长素的启动子DR5引导GUS报告基因的云烟87转基因株系。采用格盘漂浮育苗法,内装蛭石和珍珠岩(1∶1),播种50d后取长势一致的烟苗,将根部冲洗干净,移栽至60盆(高22cm、直径15cm)内装等量过60目筛石英砂的花盆中,使用改进后的Hoagland 营养液[22]培育21d,再从中选取60株长势一致的烟苗进行营养处理。其大量元素配方见表1,其他元素同Hoagland 营养液,每天定量将营养液倒入沙培底部托盘。
试验设置正常供氮(CK)处理(NH4NO3:2.5mmol/L)和低氮胁迫(-N)处理(NH4NO3:0.25mmol/L),每个处理设置3个重复,每个重复10株烟苗。缺氮处理时间为20d。
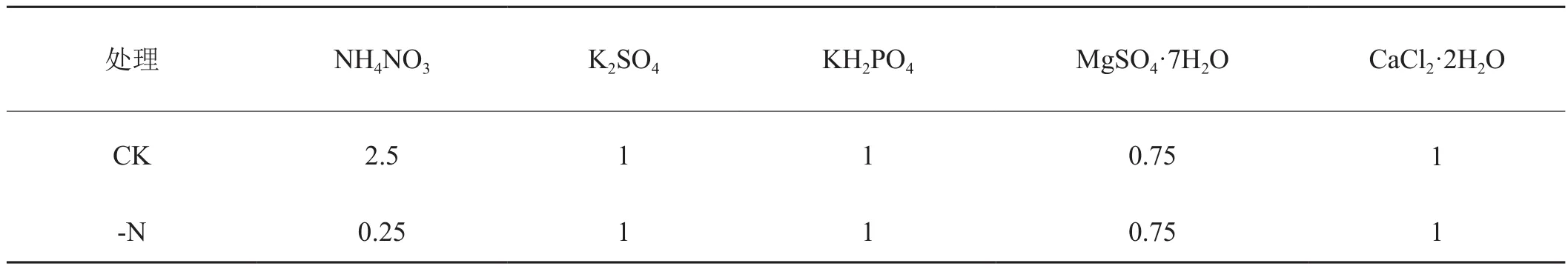
表1 试验处理方案Tab.1 Test treatment program mmol /L
1.2 样品采集与分析
生物量与全氮含量的测定:烟苗处理20d后每个重复取3株长势一致的烟苗,先用水小心冲出根系,将烟苗分上部叶(1~5叶位),中部叶(6~10叶位)、下部叶(11~15叶位)、根、茎各器官分开,拍照并称其鲜重,统计生物量,然后在110℃烘箱烘15min,调至70℃至样品恒重,采用德国Seal AA流动分析仪测定全氮含量[23]。
腺毛密度统计:烟苗处理20d后,每个重复取3株长势一致的烟苗,分别取烟苗的第3片叶(上部),第8片叶(中部)、第13片叶(下部),选取从叶尖向叶基数第6~8支脉间,主脉至右侧叶缘之间的中央位置,取5mm×5mm大小的叶片组织5-10个,将其迅速转移至4%戊二醛固定液固定,经过乙醇脱水后,固定喷金,Olympus SZX16体式显微镜观察拍照。腺毛密度统计时选取5个视野作为重复,并进行数据的统计分析[24]。
GUS染色和生长素测定:烟苗处理20d后,每个重复取3株长势一致的烟苗,分别取烟苗的第3片叶(上部),第8片叶(中部)、第13片叶(下部),从叶片中间沿主脉一分为二,一半选取从叶尖向叶基部数第6~8支脉间,主脉至叶缘中间取0.5cm×0.5cm大小的叶片组织,同时取多片用于叶片GUS染色[25],另一半液氮冷冻保存,用于生长素的测定,其方法为取样品0.2g,在石英砂、抗氧化剂(BHT)的液氮中快速研磨至粉末,然后加入80%的乙醇25mL,超声浸提15h。浸提液经旋转蒸发浓缩至5mL,再加等体积石油醚浸提。将下层液体pH调至8.5,加0.2g聚乙烯吡咯烷酮,震荡0.5h,缓冲液和样品经0.22μm的滤膜过滤[26-28]后,进行液相色谱Waters 600-2487(Waters,Milford,MA)分析,色谱柱 RT(250×4.6mm);Purospher STAR RP-18(5μm);柱温45°C;流动相:甲醇-1%乙酸(v/v,40/60),等度洗脱;液体流速:0.6mLmin−1;紫外线检测器,波长269nm;进样量20μL。IAA标准品购自Sigma-Aldrich。
NtPINs家族基因的表达:烟苗处理20d后,每个重复取3株长势一致的烟苗,将根部冲洗干净,取长1~2cm的根尖,Trizol法提取总RNA,反转录成cDNA,用于PIN家族基因qRT-PCR分析,以NtL25为内参基因,引物序列如表2。

表2 qRT-PCR引物序列Tab.2 Primer sequences used in reverse-transcription PCR analysis
1.3 数据处理
采用图像处理软件Photoshop、专业图像分析软件Image-ProPlus6.0进行腺毛密度分析、Excel 2010软件进行方差分析和多重比较。
2 结果
2.1 低氮处理下烟株表型及生物量的变化
由图1A可知,与CK相比,低氮胁迫烟苗下部叶黄化,并逐渐向上部叶扩展,植株矮小,叶片小而薄,根系除主根较粗外其他侧根纤细白嫩,主根明显伸长,侧根尤其是二级根增多;由图1B可知,与CK的烟株相比,低氮处理下的烟株除中部叶和下部叶的生物量差异没有统计学意义外,其它部位生物量与对照相比差异有统计学意义(P<0.05),分别降低了57.1%,16.79%和39.77%。
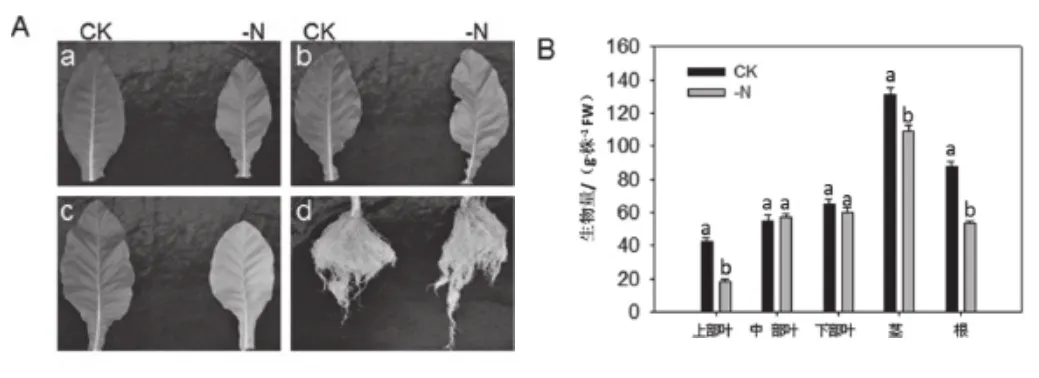
图1 低氮处理下烟株不同组织表型及生物量统计Fig.1 Different tissue phenotypes and biomass statistics of tobacco plants under total and low nitrogen treatment
2.2 低氮处理下烟株不同组织全氮含量分析
由表3可知,与对照相比,低氮胁迫下上部叶、中部叶、下部叶及根系全氮含量均显著下降,叶片全氮含量降低幅度约为33%,尤其是下部叶,全氮含量仅为1.36%,这与图1A照片中反映出的下部叶叶片变黄相一致;根部全氮含量下降幅度约为27.5%,差异有统计学意义。说明低氮胁迫下烟株叶片和根系氮素积累量明显减少。

表3 低氮处理下烟株不同组织全氮含量分析Tab.3 Analysis of total nitrogen content in different tissues of tobacco plants under low nitrogen treatment %
2.3 低氮处理下不同部位叶片的腺毛密度及生长素含量分析
将两种处理烟株不同叶位叶片进行GUS染色(GUS染色为蓝色的是新生腺毛),从图2A可以看出,低氮处理的烟苗上部叶片新生腺毛密度比CK烟苗显著增加,中部叶片新生腺毛密度降低;在两种氮素处理下的下部叶片均未被染色。低氮处理下不同部位的总腺毛密度及生长素含量的检测分析结果如图2B、2C所示;从上部叶来看,与CK相比,低氮处理下的烟株的上部叶总腺毛密度增加13.9%,差异有统计学意义有统计学意义,生长素含量也显著高于CK;对于中下部叶片,整叶总腺毛密度显著降低,分别降低40%和31.9%,生长素含量也显著低于CK。因此,腺毛的发生与生长素含量密切相关。
2.4 低氮处理对根部生长素分布及含量的影响
为确定低氮胁迫处理中生长素是否参与根系构型改变,分别检测两个处理烟苗根部的生长素浓度。由图3可知,与CK相比,低氮烟株根部的生长素浓度降低15.6%,差异有统计学意义。检测两种处理的烟株根系DR5::GUS的表达情况:与CK相比,低氮处理下的根部DR5::GUS的颜色较浅,这表明低氮处理下生长素在根系分布较少。

图2 正常供氮素与低氮处理下烟株不同部位烟叶腺毛密度及生长素含量的变化Fig.2 Changes of trichome density and auxin content in different leaves at different parts of tobacco plant under low nitrogen treatment

图3 低氮处理下烟株根部生长素分布及含量Fig.3 Auxin distribution and content in roots of tobacco plants under low nitrogen treatment
2.5 低氮处理下生长素极性运输蛋白NtPINs家族基因表达分析
为探究低氮胁迫下烟草根系中生长素含量减少的原因,采用qRT-PCR分析了CK和低氮处理下烟草根系中生长素极性运输蛋白NtPINs家族基因的表达。从表4可以看出,与CK相比,低氮处理下,除NtPIN3的表达水平降低没有统计学意义外;NtPIN1、NtPIN4、NtPIN9的基因表达水平分别降低了40%,55%,65%,差异均有统计学意义。在低氮胁迫下,生长素运输和NtPINs基因的表达水平降低,说明生长素的极性运输被抑制。

表4 低氮处理下烟叶根部NtPINs家族基因相对表达量分析Tab.4 Gene expression analysis of NtPINs family genes in tobacco root under low nitrogen treatment
3 讨论
氮素与植物的生长发育密切相关。低氮胁迫下,烟株内蛋白合成受阻,导致蛋白合成酶的数量下降,同时叶绿体的结构遭到破坏,使烟叶叶片黄化,地上部生长受阻而显得矮小[29],这可能是低氮胁迫地上部生物量降低的主要原因。根系形态在很大水平上决定植物从土壤中吸收养分的能力[30]。低氮胁迫可以改变植物根系形态以适应外界的低氮环境[31],在水稻中的研究表明,低氮胁迫下种子根长度增加,侧根密度降低,根冠比增加[32];而在拟南芥中的研究表明,低氮胁迫促进了侧根分发育,侧根变长,密度增加[33-34]。本研究中低氮(0.5mmol/L)胁迫处理的烟苗地上部和根系生物量显著下降,这表明低氮改变了烟草的生物学性状(地上部和根系构型)。
烟叶腺毛密度与发育情况很大程度上影响了烟叶香气质量,而腺毛密度又受生态环境、遗传因素和栽培条件的影响。孔光辉[35]等人的试验结果,烤烟上部叶腺毛密度最大,中部叶次之,下部叶最小,这与本试验的结果相一致。另外在不同的环境和栽培措施处理时,史宏志[36]研究在施氮量较高的情况下,腺毛密度比较低,而低施氮量时腺毛密度则较高,在中等施氮量下,较高比率的无机氮对提高烟叶表面蜡质含量更有利。本研究中,上部叶腺毛密度最大,生长素含量也相对最高,中下部叶的腺毛密度和生长素含量均明显低于上部叶,这说明烟草中生长素仍然大量产生并富集在生长活跃的区域,并且生长素在通过极性运输向中下部叶和根部的运输时受到抑制。生长素主要在植株幼嫩部位合成,能够影响细胞的伸长。Liu[37]等同位素的结果表明,低氮胁迫促进了玉米体内生长素从地上部向根部的运输。本试验条件下,与CK相比,低氮胁迫处理的烟株上部叶生长素含量明显提高,而中部叶、下部叶和根系中的生长素含量则下降,这与时向东[38]不同施氮量处理烟叶、根系中生长素的变化规律一致,说明低氮胁迫下烟草中生长素从叶片向根部的极性运输受到抑制。RT-PCR检测结果显示,与CK对照相比,NtPIN1,NtPIN3,NtPIN4和NtPIN9在根部的表达降低,NtPIN3的表达在两个处理之间差异不大。生长素从叶片向根部的极性运输主要被生长素极性运输蛋白 NtPINs家族基因所限制,其中NtPIN1,NtPIN3,NtPIN4和NtPIN9对于生长素从叶片到根部运输可能发挥了关键作用。前人研究表明生长素几乎参与植物生长发育的所有方面[39-40],DR5启动子包含生长素响应TGTCTC元件重复序列,这些元件位于CaMV35S的最小启动子和GUS报告基因的上游,具有较高的生长素反应活性;通过DR5::GUS转基因植株的GUS染色,可以反映生长素在体内的分布模式,即GUS报告基因表达的部位就是生长素分布积累的部位[41-43]。利用DR5::GUS转基因材料,将组织中的生长素通过染色拍照直观显示出来,通过GUS染色的图片更能直观的描述试验结论;并可与组织中的生长素含量的测定结果进行比较,进行相互验证。另外,本实验中统计的腺毛密度低于张亚婕[44]等的统计结果,推测一方面是因为烟株的生育期不相同,另一方面是因为烟株所处的外界环境(光照、温度等)不一致,因此导致了本实验腺毛密度统计结果与前人结果不尽一致。
4 结论
在低氮胁迫下,烟草叶片和根系的生物量减少,全氮含量降低;烟株根系发生受到抑制;低氮胁迫下上部叶腺毛密度最大,生长素含量也相对最高,而生长素由地上部向根部的极性运输明显减少。因此,低氮胁迫下烟草内生长素从新叶向根部极性运输被抑制是烟草根系对低氮胁迫响应的生理机制之一。
[1] Lynch J.Root Architecture and Plant Productivity.[J].Plant Physiology,1995,109(1):7-13.
[2] 杨中义,张发明,李永智,等.不同烤烟品种对氮素胁迫响应差异的研究[J].云南农业大学学报(自然科学版),2011,26(2):240-245.YANG Zhongyi,ZHANG Faming,LI Yongzhi,et al.Evaluation of Low Nitrogen Stress for Diff erent Tobacco Varieties[J].Journal of Yunnan Agricultural University,2011.
[3] 黄荣,孙虎威,刘尚俊,等.低磷胁迫下水稻根系的发生及生长素的响应[J].中国水稻科学,2012,26(5):563-568.HUANG Rong,SUN Huwei,LIU Shangjun,et al.Rice Root Growth and Auxin Concentration in Response to Phosphate Deficiency[J].Chinese Journal of Rice Science,2012,26(5):563-568.
[4] 刘国顺,秦菲,王彦亭,等.烟草生长过程中根系内源激素含量的变化规律[J].中国农学通报,2005,21(4):179-181.LIU Guoshun,QIN Fei,WANG Yanting,et al.Law of Endogenous Hormone Variation of Growing Tobacco Root[J].Chinese Agricultural Science Bulletin,2005,21(4):179-179.
[5] Fang B,Li J.Evidence that the auxin signaling pathway interacts with plant stress response[J].植物学报(英文版),2002,44(5):532-536.
[6] Tian Q,Chen F,Liu J,et al.Inhibition of maize root growth by high nitrate supply is correlated with reduced IAA levels in roots.[J].Journal of Plant Physiology,2008,165(9):942-951.
[7] ang,JiRong,Hu,et al.Expression of PIN genes in rice(Oryza sativa L.):tissue specificity and regulation by hormones.[J].Molecular Plant,2009,2(4):823-31.
[8] Benková E;Michniewicz M;Sauer M;Teichmann T;Seifertová D;Jürgens G;Friml J.Local,Efflux-Dependent Auxin Gradients as a Common Module for Plant Organ Formation[J].Cell,2003,115(5):591-602.
[9] 胡一兵,刘炜,徐国华.生长素与乙烯信号途径及其相互关系研究进展[J].植物学报,2011,46(3):338-349.HU Yibing,LIU Wei,XU Guohua,et al.Research Advances in Auxin and Ethylene Signaling and Eff ects of Auxin on Ethylene Response of Plants[J].Chinese Bulletin of Botany,2011,46(3):338-349.
[10] An L,Zhou Z,Yan A,et al.Progress on trichome development regulated by phytohormone signaling[J].Plant Signaling &Behavior,2011,6(12):1959.62.
[11] 刘金秋,陈凯,张珍珠,等.外施GA、MeJA、IAA、SA和KT对番茄表皮毛发生的作用[J].园艺学报,2016,43(11):2151-2160.LIU Jinqiu,CHEN Kai,ZHANG Zhenzhu,et al.Effects of Exogenous GA,MeJA,IAA,SA and KT on Trichome Formation in Tomato[J].ACTA HORTICULTURAE SINICA,2016,43(11):2151-2060
[12] Deng W,Yang Y,Ren Z,et al.The tomato SlIAA15,is involved in trichome formation and axillary shoot development[J].New Phytologist,2012,194(2):379.
[13] Traw M B,Bergelson J.Interactive effects of jasmonic acid,salicylic acid,and gibberellin on induction of trichomes in Arabidopsis.[J].Plant Physiology,2003,133(3):1367-1375.
[14] 李鹏飞,周冀衡,张建平,等.氮、磷、钾、镁亏缺对烤烟生长和叶片腺毛发育的影响[J].烟草科技,2009(12):49-54.LI PengFei,ZHOU Jiheng,ZHANG jianping,et al.Effects of N,P,K,Mg Deficiency on Plant Growth and Leaf Glandular Trichome Development of Flue-cured Tobacco[J].Tobacco Science&Technology,2009(12):49-54.
[15] 赵珺,陈丹,安鹏,等.烟草花发育过程中花药和花粉IAA分布规律的研究[J].武汉植物学研究,2009,27(5):541-547.ZHAO Jun,CHEN Dan,AN Peng,et al.IAA Distribution of Anther and Pollen during Flower Development of Nicotiana tabacum L.2009,27(5):541-547.
[16] 邢瑶,马兴华.氮素形态对烟苗根系生长及氮素利用的影响[J].中国烟草学报,2016,22(4):52-61.XING Yao,MA Xinghua.Effect of different nitrogen forms on tobacco seedling root growth and nitrogen utilization[J].Acta Tabacaria Sinica,2016,22(4):52-61.
[17] Itai C,Ben-Zioni A,Ordin L.Correlative Changes in Endogenous Hormone Levels and Shoot Growth Induced by Short Heat Treatments to the Root[J].Physiologia Plantarum,2010,29(3):355-360.
[18] 史宏志,韩锦峰,刘卫群,等.氮素营养对烤烟类脂物含量和脂肪氧化酶活性的影响[J].中国烟草学报,1997,3(2):41-48.SHI Hongzhi,HAN Jinfeng,LIU Weiqun,et al.Eff ects of Nitrogen Nutrition on Lipid Content and Lipoxygenase Activity in Fulecured Tobacco Leaves[J].Acta Tabacaria Sinica,1997,3(2):41-48.
[19] 梁志敏,翁梦苓,崔红.施用有机肥对烟草腺毛形态结构及分泌物的影响[J].厦门大学学报(自然科学版),2008,47(S2):149-152.Liang Zhimin,Weng Mengling,Cui Hong.Effects of Applying Organic Fertilizer on Morphology and Structure of Chloroplast and Exudates of Flue-cured Tobacco Trichomes[J].Journal of Xiamen University,2008,47(s2):149-152.
[20] Song W,Liu S,Meng L,et al.Potassium deficiency inhibits lateral root development in tobacco seedlings by changing auxin distribution[J].Plant and Soil,2015,396(1):163-173.
[21] Rubio V,Bustos R,Irigoyen M L,et al.Plant hormones and nutrient signaling.[J].Plant Molecular Biology,2009,69(4):361.
[22] Chen A,Hu J,Sun S,et al.Conservation and divergence of both phosphate- and mycorrhiza-regulated physiological responses and expression patterns of phosphate transporters in solanaceous species[J].New Phytologist,2007,173(4):817-831.
[23] 张丽萍,王久荣,袁红朝,等.植物全氮的AA3型流动分析仪测定方法研究[J].湖南农业科学,2016(10):83-86.ZHANG Liping,WANG Jiurong,YUAN Hongchao,et al.The Determination of Total Kjeldahl Nitrogen in Plants by AA3 Flow Analyzer[J].Hunan Agricultural Sciences,2016(10):83-86.
[24] 杨永霞,魏跃伟,冯琦,等.典型生态区烟叶腺毛发育及其分泌物比较研究[J].中国烟草学报,2014,20(5):38-42.YANG Yongxia,WEI Yuewei,FENG Qi,et al.Comparative study in leaf trichome development and exudates in typical ecological regions[J].Acta Tabacaria Sinica,2014,20(5):38-42.
[25] Song W,Liu S,Meng L,et al.Potassium deficiency inhibits lateral root development in tobacco seedlings by changing auxin distribution[J].Plant &Soil,2015,396(1-2):163-173.
[26] Walchliu P,Neumann G,Engels C.Response of shoot and root growth to supply of different nitrogen forms is not related to carbohydrate and nitrogen status of tobacco plants.[J].Journal of Plant Nutrition &Soil Science,2001,164(1):97-103.
[27] Takei K,Sakakibara H,Taniguchi M,et al.Nitrogen-Dependent Accumulation of Cytokinins in Root and theTranslocation to Leaf:Implication of Cytokinin Species that Induces GeneExpression of Maize ResponseRegulator[J].Plant &Cell Physiology,2001,42(1):85-93.
[28] Xiong J,Lu H,Lu K X,et al.Cadmium decreases crown root number by decreasing endogenous nitric oxide,which is indispensable for crown root primordia initiation in rice seedlings.[J].Planta,2009,230(4):599-610.
[29] 张柳,王铮,张亚婕,等.烟草叶片衰老期过程中的蛋白质组学分析[J].植物生理学报,2014,50(4):488-500.ZHANG Liu,WANG Zheng,ZHANG Yajie,et al.Proteomic analysis of senescing leaf of tobacco[J].Plant Physiology Journal,2014,50(4):488-500.
[30] Vidal E A,Moyano T C,Riveras E,et al.Systems approaches map regulatory networks downstream of the auxin receptor AFB3 in the nitrate response of Arabidopsis thaliana roots[J].Proceedings of the National Academy of Sciences of the United States of America,2013,110(31):12840-12845.
[31] Walchliu P,Ivanov I I,Filleur S,et al.Nitrogen Regulation of Root Branching[J].Annals of Botany,2006,97(5):875-881.
[32] 赵学强,施卫明.水稻根系生长对不同氮形态响应的动态变化[J].土壤,2007,39(5):766-771.ZHAO Xueqiang,SHI Weiming.Dynamic Responses of Rice in Root Growth to Diff erent NH_4~+ and NO_3~- Supply[J].Soils,2007.34(3):530-535.
[33] Linkohr B I,Williamson L C,Fitter A H,et al.Nitrate and phosphate availability and distribution have different effects on root system architecture of Arabidopsis[J].Plant Journal for Cell &Molecular Biology,2002,29(6):751-760.
[34] Gruber B D,Giehl R F,Friedel S,et al.Plasticity of the Arabidopsis root system under nutrient deficiencies[J].Plant Physiology,2013,163(1):161-179.
[35] 孔光辉,宗会.不同部位成熟烟叶腺毛密度及其分泌物的研究[J].中国农学通报,2006,22( 12):108-110.KONG Ganghui,ZONG Hui.Study on the Density and Exudates of Glandular Trichome of Mature Tobacco Leaves with Diff erent Stalk Positions[J].Chinese Agricultural Science Bulletin,2006,22( 12):108-110.
[36] 史宏志,韩锦峰,刘卫群,等.氮素营养对烤烟类脂物含量和脂肪氧化酶活性的影响[J].中国烟草学报,1997,3(4):41-48.SHI Hongzhi,HAN Jinfeng,LIU Weifeng,et al.Effects of Nitrogen Nutrition on Lipid Content and Lipoxygenase Activity in Fule-cured Tobacco Leaves[J].Acta Tabacaria Sinica,1997,3(4):41-48.
[37] Liu J,Xia A,Lei C,et al.Auxin transport in maize roots in response to localized nitrate supply [J].Annals of Botany,2010,106(6):1019-1026.
[38] 时向东,刘艳芳,文志强,等.施N水平对雪茄外包皮烟叶片生长发育和内源激素含量的影响[J].西北植物学报,2007,27(8):1625-1630.SHI Xiangdong,LIU Yanfang,WEN Zhiqiang,et al.Effects of nitrogen levels on growth and content of endogenous hormones of cigar wrapper leaves[J].Acta Botanica Boreali-Occidentalia Sinica,2007,27(8):1625-1630.
[39] Mockaitis K,Estelle M.Auxin receptors and plant development:a new signaling paradigm.[J].Annu Rev Cell Dev Biol,2008,24(24):55-80.
[40] Weijers D,Friml J.SnapShot:Auxin Signaling and Transport[J].Cell,2009,136(6):1172-1172.
[41] Sabatini S,Beis D,Wolkenfelt H,et al.An auxin-dependent distal organizer of pattern and polarity in the Arabidopsis root.[J].Cell,1999,99(5):463-472.
[42] Ulmasov T,Murfett J,Hagen G,et al.Aux/IAA proteins repress expression of reporter genes containing natural and highly active synthetic auxin response elements[J].Plant Cell,1997,9(11):1963-1971.
[43] Miura K,Lee JY,Gong QQ,et al.SIZ1 Regulation of Phosphate Starvation-Induced Root Architecture Remodeling Involves the Control of Auxin Accumulation [J].Plant Physiology,2011,155(2),1000-1012.
[44] 张亚婕,徐琼华,毛自朝,等.4个烤烟品种不同采收期烟叶腺毛密度和形态比较[J].云南农业大学学报(自然科学版),2015,30(1):35-43.ZHANG Yajie,XU Xionghua,MAO Zichao,et al.Comparison of Density and Morphology of Glandular Trichome of Tobacco Leaves of Four Species at Diff erent Collection Periods[J].Journal of Yunnan Agricultural University,2015.
