两种黑麦草砷吸收特征及其与茎叶营养元素积累的关系研究
2018-03-04李金波李诗刚宋桂龙濮阳雪华薛博晗北京林业大学草坪研究所北京00083深圳市铁汉生态环境股份有限公司广东深圳58040
李金波,李诗刚,宋桂龙*,濮阳雪华,薛博晗(.北京林业大学草坪研究所,北京 00083;.深圳市铁汉生态环境股份有限公司,广东 深圳 58040)
砷(arsenic,As)是一种剧毒物质,对生物体具有致癌、致突变的效应,是生物体的非必需元素,被世界卫生组织列为一级致癌物。砷在地壳中的含量较低,在自然界中多以砷酸盐[arsenate,As(Ⅴ)]和亚砷酸盐[arsenite,As(Ⅲ)]的形态存在,通过植物经食物链对人类健康构成极大威胁[1-2]。我国面临严重的砷污染问题,大量含砷农业药剂的施用以及矿山的开采均导致农田和矿山土壤中的砷含量大幅上升[3]。重金属污染的治理问题是目前世界各国共同面临的一大难题,我国也为此开展了大量研究,目前比较新兴的则是植物修复技术,利用植物吸收富集重金属,具有成本低、环境扰动小、易于操作等优点[4]。草本植物具有生长迅速,耐性强等特点,且能弥补超富集植物生物量小、种子不易收集等缺点。目前已有大量研究证明常见草本植物是植物修复工程中极具潜力的优势植物。
黑麦草(Loliumperenne)作为常见的禾本科草本植物,对重金属有较强的抗性和富集能力,具有极大的潜力应用于环境治理以及生态环境保护工程中[5]。同时黑麦草具有生长迅速、产量高、耐频繁修剪等特点,且有研究证明适宜的修剪或刈割能够提高植物对重金属污染土壤的修复效率[6],因此黑麦草是极具潜力的植物修复植物。袁敏等[7]研究了黑麦草、高羊茅(Festucaarundinacea)、早熟禾(Poapratensis)以及紫花苜蓿(Medicagosativa)对铅锌尾矿土壤重金属的抗性与吸收特性,结果表明3种禾本科草的抗性要大于豆科的紫花苜蓿,每种植物对Zn、Pb、Cu、Cd四种重金属的吸收与分布均表现为根系大于茎叶,且4种植物对重金属的吸收量为黑麦草>高羊茅>早熟禾>紫花苜蓿。徐卫红等[8]采用盆栽试验研究了Zn、Cd单一及复合污染对黑麦草的生长及生理的影响,结果表明黑麦草对Zn有较强的抗性和耐性,在16 mmol·kg-1的高浓度胁迫下也没有表现出抑制作用。黑麦草吸收重金属后在不同组织及不同细胞器的分配不同,张尧等[9]研究了黑麦草幼苗对Cd的耐性及吸收和细胞分布的特点,结果表明黑麦草在10和20 mg·kg-1的Cd胁迫下,富集系数>1,转运系数<1,说明黑麦草对Cd富集能力较强且主要积累在根部。
植物受到重金属毒害的原因之一是养分吸收受到干扰,体内的养分平衡遭到破坏[10]。吴福忠等[11]研究了Cd胁迫对桂花(Osmanthussp.)生长和养分积累、分配和利用的影响,结果表明植物C、N、P的积累量随着Cd浓度的增加而降低。高浓度的Cd处理(≥25 mg·kg-1)能明显抑制长春花(Catharanthusroseus)C、N、P、K的积累并显著改变其分配格局[12]。张骞等[13]对不同品种苋菜(Amaranthusspinosus)对砷的吸收和植株磷砷关系进行研究,得出在2 mg·L-1As(Ⅴ)处理下,苋菜地上部P含量和As含量有显著相关性(r=0.881),在4 mg·L-1As(Ⅴ)处理下,二者相关性不显著,但是相关系数达到0.816。研究植物在重金属胁迫下的养分吸收状况,能够为植物的实际应用提供理论基础。前人虽然对黑麦草在金属胁迫下耐性及富集进行了一些研究,但大多集中在Cd、Pb等金属元素,对营养元素的影响方面也大多集中在大量元素上,而黑麦草在As胁迫下对As元素的吸收及不同部位分配以及与养分吸收关系未见报道。因此,本研究以一年生黑麦草(Loliummultiflorum)和多年生黑麦草(Loliumperenne)为实验材料,研究其在As胁迫下各部位对砷以及营养元素的吸收特征,并分析茎叶部位营养元素与As的相关性,以期为黑麦草在植物修复工程上的应用以及养分管理提供理论参考。
1 材料与方法
1.1 供试材料
供试土壤:将北京林业大学草坪研究所昌平实验站表层土风干过5 mm筛,按照14∶1(w/w)将土壤和腐熟鸡粪进行充分混匀。供试土壤类型为潮土,质地中壤,全氮1.33 g·kg-1,有效磷13.9 mg·kg-1,速效钾70.2 mg·kg-1,有机质8.2 g·kg-1,pH值7.68,As含量0.337 mg·kg-1。
供试植物:多年生黑麦草和一年生黑麦草,品种分别为Mathilde和Idyll,种子均来自北京正道生态科技有限公司。
1.2 试验设计
采取完全随机区组设计,设置2个As浓度梯度,分别是0,100 mg·kg-1(以装填混合土重,纯As计),分别用DCK、D100、YCK、Y100(D代表多年生黑麦草,Y代表一年生黑麦草)表示。每个处理设3个重复,共计12盆。
实验处理于2016年6-10月在北京林业大学草坪研究所昌平实验站进行。采用盆栽实验,选用锥形塑料花盆(上下内径分别为20 和10 cm,高28 cm),每盆装入混合土3 kg。As施入采用NaAsO2配成200 mL溶液浇入土壤,静置2周。
黑麦草先在育苗盘里育苗,基质采用草炭∶蛭石=1∶1充分混合,待出苗10 d后选取长势一致的幼苗移栽至As处理花盆,生长适应2周后选择长势良好且一致的植物定株,每盆10株。定株后定期定量浇水培养60 d。
1.3 株高及根长的测定
将植株连带土壤整体取出浸入水中,清洗根部土壤以获得完整根系,蒸馏水冲洗数次后用直尺测量株高和根长。
1.4 植株生物量的测定
参考徐佩贤[14]的方法(稍作修改)将各处理的草按照根系、茎、老叶、功能叶、新叶五部位分解开,用蒸馏水洗净并擦干表面水分,分别收集置于105 ℃杀青30 min,然后80 ℃烘干至恒重,分别测定植株各部分的干重。
1.5 砷及营养元素含量测定
烘干的样品使用粉碎机粉碎,过60目(0.3 mm)筛后称取0.500 g,使用Hanon220s石墨消解仪,采用HNO3-H2O2消化完全后,定容至50 mL,采用ICP-MS(安捷伦 7700)测定As、P、K、Ca、Mg、Mn含量。然后称取0.200 g,采用H2SO4-H2O2消化完全后定容至100 mL,采用全自动流动分析仪(SEAL AA3)测定全N含量。
1.6 数据处理及分析
原始数据经Excel 2010软件整理,运用SPSS for Windows 17.0软件进行单因素方差分析及相关性分析,分析结果用Origin作图。
2 结果与分析
2.1 As胁迫对株高及根长影响
As处理下多年生黑麦草与一年生黑麦草的株高和根长见图1,当As浓度为 100 mg·kg-1时,两种黑麦草的株高和根长都受到了抑制。多年生黑麦草在砷胁迫下显著低于对照(P<0.05),一年生黑麦草与对照相比也受到抑制,但没有达到显著水平(P>0.05)。二者的株高分别降低了33.4%和13.3%,根长分别降低了14.9%和3.3%,说明一年生黑麦草在100 mg·kg-1As 胁迫下的耐性要优于多年生黑麦草。
2.2 As胁迫对5部位干重影响
两种黑麦草在砷胁迫下各部位的干重如图2所示。与对照相比,两种草各部位干重均呈现下降趋势,且除多年生黑麦草老叶以及一年生黑麦草根系外,其余部位都达到显著水平(P<0.05)。多年生黑麦草和一年生黑麦草的老叶分别降低了25.0%和44.7%,功能叶分别降低35.3%和39.8%,新叶分别降低57.7%和28.1%,茎分别降低44.5%和44.3%,根系分别降低44.0%和6.7%。As处理下多年生黑麦草和一年生黑麦草老叶的耐受系数(处理值/对照值)分别为75.0%和55.3%,功能叶耐受系数分别为64.7%和60.2%,新叶耐受系数分别为42.3%和71.9%,茎耐受系数分别为55.5%和55.7%,根系耐受系数分别为56.0%和93.3%。结果表明,一年生黑麦草根系耐性和新叶耐性优于多年生黑麦草,而多年生黑麦草老叶的耐性优于一年生黑麦草。
2.3 As胁迫下5个部位砷含量特征
As处理下多年生黑麦草和一年生黑麦草5个部位砷含量如图3。两种草5个部位As含量均呈现根系>老叶>茎>功能叶>新叶。在同样的As胁迫下,一年生黑麦草5个部位As含量均低于多年生黑麦草,且功能叶、新叶、茎、根系4部分达到显著水平(P<0.05),老叶、功能叶、新叶、茎、根系各部位分别是多年生黑麦草的95.5%、72.9%、84.2%、81.4%、83.0%,说明一年生黑麦草对As吸收能力要低于多年生黑麦草,这也可能是多年生黑麦草株高、根长被抑制更明显的原因。
两种黑麦草地下根系As含量都远远高于地上4部分,As处理下多年生黑麦草老叶、功能叶、新叶、茎4部位分别占其根系含量的4.6%、1.8%、1.0%、3.6%,一年生黑麦草分别是5.2%、1.6%、1.0%、3.5%,说明两种黑麦草对砷的转运能力都较低,体内砷主要集中在根系,同时两种草对砷的运输和分配较为一致。
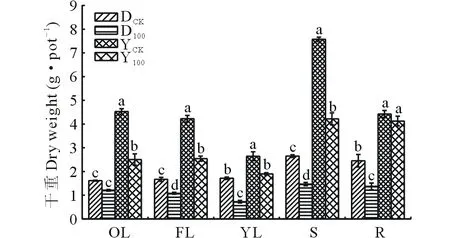
图2 As胁迫对两种黑麦草5个部位干重影响Fig.2 Effect of As on five tissues biomass of two ryegrass species

图3 As胁迫下两种黑麦草5个部位As含量Fig.3 As concentration of five tissues of two ryegrass species under As stress
OL:老叶 Old leaves;FL: 功能叶 Function leaves;YL:新叶 Young leaves;S:茎 Stem;R:根系 Root. 下同 The same below.
2.4 As胁迫下茎叶部位养分含量特征
As胁迫下多年生黑麦草和一年生黑麦草地上部分各部位的营养元素含量如图4。同一植物各个部位对养分元素的吸收情况各不相同。N元素,多年生黑麦草在砷处理下新叶部分表现出显著增高趋势(P<0.05),其余3部位均略有下降但都没有达到显著水平;一年生黑麦草则在老叶中表现出显著增高趋势(P<0.05),而在新叶中显著降低(P<0.05),茎中没有显著性差异。P元素,多年生黑麦草在砷处理下老叶和功能叶中表现出显著降低趋势(P<0.05),新叶中呈现显著增高(P<0.05),但茎中并无显著性差异;一年生黑麦草在老叶和新叶中呈现显著增高(P<0.05),但在功能叶和茎中没有显著性差异。As胁迫下两种黑麦草对K元素的吸收与分配则呈现出老叶和茎中没有显著性差异,但多年生黑麦草在功能叶中显著降低(P<0.05),两种草在新叶中都显著增高(P<0.05)。Ca元素,As胁迫使多年生黑麦草老叶和功能叶中显著增高(P<0.05),茎中也有增高但差异不显著,一年生黑麦草却在茎中显著增高(P<0.05),老叶和功能叶中也有增高但未达到显著水平,二者在新叶中都无显著性差异。Mg元素,两种草在老叶和功能叶中都呈现显著降低趋势(P<0.05),新叶中均无显著性差异,但在茎中,多年生黑麦草显著增高而一年生黑麦草显著降低(P<0.05)。Mn元素,多年生黑麦草在3类叶片中都有降低,在老叶以及功能叶中达显著水平(P<0.05),而一年生黑麦草则在叶片中没有显著性差异,在茎中多年生黑麦草显著增高(P<0.05),一年生黑麦草显著降低(P<0.05)。
无论是对照组还是处理组,两种黑麦草的3类叶片对于N、P、K三种元素的吸收都呈现新叶>功能叶>老叶,这说明植物的新叶新陈代谢速率较快,为维持正常的生长需要吸收消耗更多的大量元素。而Ca、Mg、Mn三种元素则呈现出相反的结果。总的来讲,两种黑麦草对于这6种营养元素的选择性由强到弱依次是K、N、Ca、Mg、P、Mn。两种草在不同部位间对养分的吸收与分配不同,在同一部位两种草之间也表现出较大的差异,说明植物对营养元素的吸收与分配与植物基因型有关。

图4 As胁迫下两种黑麦草4个茎叶部位N、P、K、Ca、Mg、Mn含量Fig.4 N, P, K, Ca, Mg, Mn concentration of four shoot tissues of two ryegrass species shoots under As stress
2.5 茎叶部位养分含量与砷的关系
对两种黑麦草地上部各部位养分含量与As含量做相关性分析,结果如表1。对于老叶而言,多年生黑麦草As与P、Mg呈显著负相关关系,与Mn呈极显著负相关关系;一年生黑麦草As与N、P、Ca呈显著正相关关系,与Mg呈极显著负相关关系。对于功能叶而言,多年生黑麦草As与N呈显著负相关关系,与K、Mg、Mn呈极显著负相关关系,与Ca呈显著正相关关系;一年生黑麦草As只与Mg呈极显著负相关关系。对于新叶而言,多年生黑麦草As与N、K呈极显著正相关关系;一年生黑麦草As与P呈显著正相关关系,与K 呈极显著正相关关系。对于茎而言,多年生黑麦草As与Mg呈显著正相关关系,与Mn呈极显著正相关关系;一年生黑麦草As与Ca呈显著正相关关系,与K呈极显著正相关关系,与Mn呈显著负相关关系。在两种黑麦草的叶片中,As与Mg在老叶和功能叶中呈现出一致的显著或极显著负相关关系,在新叶中两种元素的吸收没有相关性,说明两种黑麦草叶片对于As和Mg的吸收存在竞争关系。在一年生黑麦草的老叶和茎以及多年生黑麦草的功能叶中,As和Ca则呈现出一致的显著正相关关系,两种草对As和Ca的吸收在其他部位都没有显著或极显著负相关关系的呈现,说明黑麦草对于As和Ca的吸收可能存在协同作用。在两种黑麦草的新叶中,As与K都呈现极显著正相关关系,多年生黑麦草As与N 呈现极显著正相关关系,而一年生黑麦草As与P呈现显著正相关关系,说明两种黑麦草的新叶对于As以及N、P、K三种大量元素的吸收可能存在协同作用。
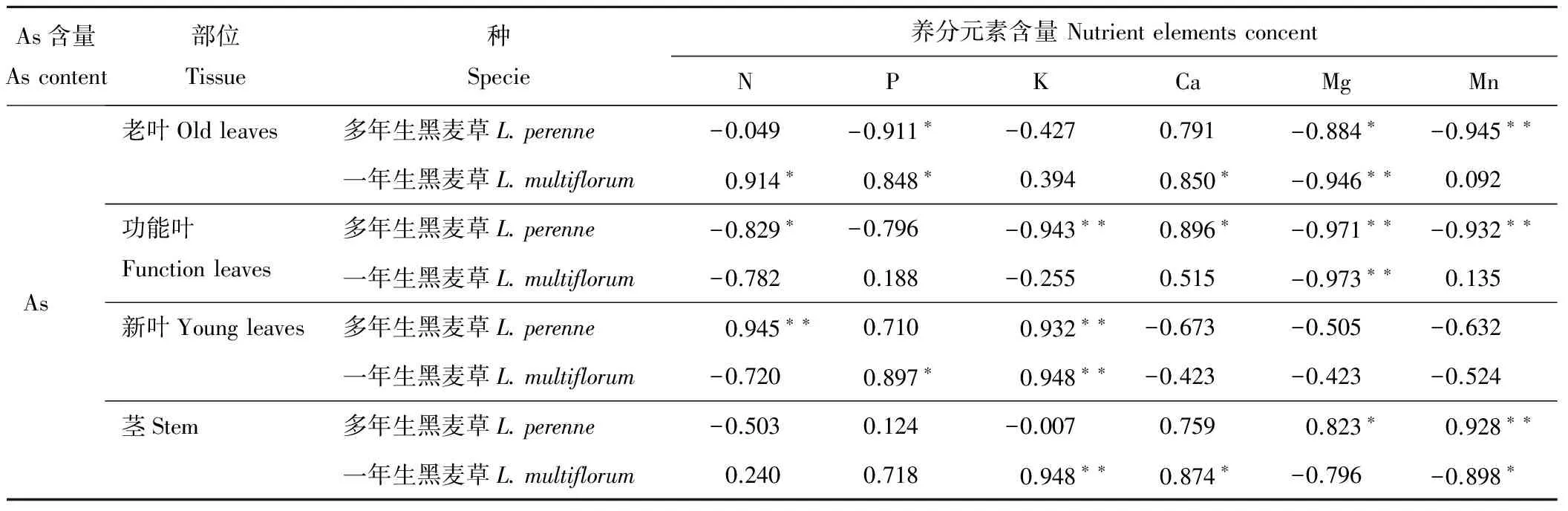
表1 两种黑麦草茎叶部分养分含量与砷的相关系数Table 1 Correlation coefficients between nutrient elements and As in the shoot of two ryegrass species
注:*,在 0.05 水平(双侧)上显著相关,**,在 0.01 水平(双侧)上显著相关。
Note: * means significant correlation (P<0.05), ** means highly significant correlation (P<0.01).
3 讨论
3.1 As对植物影响
As是植物生长的非必需元素,大量研究表明低浓度的As会促进植物的生长,而达到较高浓度时则会抑制植物的生长[15-16]。本研究中,两种黑麦草在100 mg·kg-1的高浓度下株高、根长、生物量都出现不同程度的降低,生长都受到抑制,这与前人的研究结果基本一致[17]。As对植物的毒害首先表现在叶上,导致叶片枯萎或脱落,其次会让植物根系伸长受到抑制,从而使植物整体的生长发育都受到严重抑制[18],本研究中一年生黑麦草根系抑制明显小于地上部,但多年生黑麦草却没有表现出类似结果,且多年生黑麦草老叶和一年生黑麦草根系和对照相比都没有显著性差异,这说明多年生黑麦草老叶耐性优于一年生黑麦草,而一年生黑麦草根系耐性优于多年生黑麦草,出现这一现象的原因可能是与植物基因型有关。一般认为,高浓度As胁迫导致植物出现生物量降低的根本原因是As使植物可溶性蛋白含量降低、过氧化酶活性下降、光合作用减弱等生理过程的变化[19]。
重金属在植物体内不同部位的积累和分布特征决定了植物受到重金属的毒性大小[20],同时也是研究植物吸收转运重金属以及对重金属耐性机制的基础[21]。有的植物为了防止重金属对自身光合作用的影响以及对叶片新陈代谢的毒害,当其吸收重金属后会将大量重金属富集在根部,只有较少一部分转运到茎叶,这是植物对逆境的一种适应机制[22]。本研究中两种黑麦草在As胁迫下都将大量的As元素富集在根部,说明黑麦草在重金属胁迫下有以上适应机制,类似结果在张尧等[9]的研究中也有体现。两种黑麦草对于As的地上分配都呈现老叶>茎>功能叶>新叶,这也是植物的一种保护机制,将转运到地上部的重金属离子贮存在老叶当中,随着老叶的逐渐脱落将重金属排出体外,从而可以减少新叶的吸收,降低自身的毒害,这与徐佩贤[14]在高羊茅和草地早熟禾上Cd分布的研究结果相类似,同时类似结果在蔡成翔等[23]在水葫芦(Eichhorniacrassipes)的研究结果中也有体现。两种黑麦草对于As的吸收能力差别较大,这与植物的基因型有关,李希铭[24]在草本植物Cd的耐性及富集特征研究中,黑麦草也表现出了种与品种之间的较大差异。
3.2 As与营养元素的关系
As对植物生长的影响可能并不是源于对植物与生物相互作用的干扰,而是源于As与植物营养或者植物代谢的相互作用,As可能通过影响营养元素的吸收从而间接影响到植物的生长状况[25-26]。当土壤环境发生变化时,植物会主动调节养分需求来适应土壤环境,从而调整体内的元素丰度[12]。前人对于植物吸收As与N的相互关系的研究较少,有研究报道As会影响烤烟(Nicotianatabacum)的N代谢过程,使总N含量以及硝酸还原酶(NR)活性降低[27]。本研究中两种黑麦草在功能叶中N含量都呈现降低趋势,但多年生黑麦草新叶以及一年生黑麦草老叶中As与N却呈现显著正相关关系,与前人结果相反,这可能与植物的基因型不同有关,也说明不同植物对于逆境的适应机制不同。一年生黑麦草各部位对于P的吸收都有增高趋势,在老叶和新叶中呈显著正相关关系,这可能与磷和砷两种元素共用一个转运通道和受同一基因控制有关,这在张骞等[13]的研究中也有体现。两种黑麦草在As胁迫下新叶中K含量都显著增高,这可能是因为K是植物适应逆境的抗逆元素[28],植物为降低新叶的毒害而向新叶中分配更多的K元素。As对于冬小麦(Triticumaestivum)根系中的Ca含量影响较小[29],本研究结果却表明As与Ca在一年生黑麦草的老叶和茎以及多年生黑麦草的功能叶中呈现显著正相关关系,类似结果不适用于蜈蚣草(Pterisvittata),有研究表明蜈蚣草中的砷积累量同时受钙水平和砷水平的限制[30]。Mg是组成叶绿素的重要成分,而叶绿素含量又是光合作用的物质基础,As在植物体内能取代叶绿素分子中的Mg离子,阻碍叶绿素的合成,从而影响植物的光合作用。本研究中两种黑麦草3种叶片中Mg含量都出现降低趋势,并且在老叶和功能叶中呈现极显著负相关关系。植物在砷胁迫下对Mn、Fe、Zn等微量元素的影响大于对大量元素的影响,本研究中多年生黑麦草老叶与功能叶中As与Mn都呈极显著负相关关系,类似结果在刘全吉等[29]在冬小麦上的研究结果中也有体现。综上,植物吸收重金属与营养元素之间的交互作用机理十分复杂,植物的基因型不同也会导致重金属与营养元素间的较大差异[31],本研究中两种黑麦草也表现出较大差异。重金属与营养元素相互作用的机理,还有待在分子水平上进一步研究。
4 结论
1)砷胁迫下,一年生黑麦草株高和根长受抑制程度显著小于多年生黑麦草;二者的地上和地下生物量都呈现降低趋势,只有多年生黑麦草老叶和一年生黑麦草根系没有达到显著水平。
2)二者对于砷元素的分配情况较为一致,都表现为根系>老叶>茎>功能叶>新叶,但多年生黑麦草吸收能力要优于一年生黑麦草。二者地上部养分含量都受到了砷胁迫的影响,同一部位不同植物种之间以及同一植物种不同部位之间都表现出较大差异,比较一致的是二者在叶片中的镁含量都出现降低趋势,在老叶和功能叶中的钙含量都有增高趋势。二者对于N、P、K三种元素的吸收均呈现新叶>功能叶>老叶,对于Ca、Mg、Mn三种元素的吸收则呈现老叶>功能叶>新叶。
3)两种黑麦草在同浓度砷胁迫下对于砷以及营养元素吸收的相关性表现出较大差异。As与Mg在两种黑麦草的老叶和功能叶中都呈现出一致的显著或极显著负相关关系;As与Ca则在一年生黑麦草的老叶和茎以及多年生黑麦草的功能叶中呈现出一致的显著正相关关系;As与K在两种黑麦草的新叶中都呈现出极显著正相关关系。
References:
[1] Jiang H M, Deng T L, Lai D M,etal. Progresses on the effect of arsenic for plant and plant tolerant mechanism. Guangdong Trace Elements Science, 2009, (11): 1-5.
蒋汉明, 邓天龙, 赖冬梅, 等. 砷对植物生长的影响及植物耐砷机理研究进展. 广东微量元素科学, 2009, (11): 1-5.
[2] Kamnev A A, Van D. Chemical and biological parameters as tools to evaluate and improve heavy metal phytoremediation. Bioscience Reports, 2000, (20): 239-258.
[3] Nriagu J O, Pacyna J M. Quantitative assessment of worldwide contamination of air, water and soils by trace metals. Nature, 1988, 333(6169): 134-139.
[4] Leduc D L, Terry N. Phytoremediation of toxic trace elements in soil and water. Journal of Industrial Microbiology Biotechnology, 2005, (32): 514-520.
[5] Liao M, Huang C Y. Effects of organic acids on the toxicity of cadmium during ryegrass growth. Chinese Journal of Applied Ecology, 2002, (1): 109-112.
廖敏, 黄昌勇. 黑麦草生长过程中有机酸对镉毒性的影响. 应用生态学报, 2002, (1): 109-112.
[6] Li N, Sun N X, Song G L,etal. Effects of cutting frequency on cadmium uptake and physiological responses ofMedicagosativaunder cadmium stress. Acta Prataculturae Sinica, 2017, 26(5): 109-117.
李娜, 孙宁骁, 宋桂龙, 等. 刈割次数对紫花苜蓿镉吸收影响及生理响应. 草业学报, 2017, 26(5): 109-117.
[7] Yuan M, Tie B Q, Tang M Z,etal. Resistance and uptake of heavy metals by four herbaceous plants from soil polluted lead/zinc mine tailings. Acta Prataculturae Sinica, 2005, 14(6): 57-62.
袁敏, 铁柏清, 唐美珍, 等. 四种草本植物对铅锌尾矿土壤重金属的抗性与吸收特性研究. 草业学报, 2005, 14(6): 57-62.
[8] Xu W H, Wang H X, Wang Z Y,etal. Response of hyperaccumulator ryegrass (LoliurnperenneL.) to cadmium, zinc and their combined pollution. Chinese Agricultural Science Bulletin, 2006, (6): 365-368.
徐卫红, 王宏信, 王正银, 等. 重金属富集植物黑麦草对锌、镉复合污染的响应. 中国农学通报, 2006, (6): 365-368.
[9] Zhang Y, Tian Z G, Cao C L,etal. Characteristics of cadmium tolerance, accumulation and cellular distribution inLoliumperenneseedlings. Journal of Agro-Environment Science, 2010, (11): 2080-2086.
张尧, 田正贵, 曹翠玲, 等. 黑麦草幼苗对镉耐性能力及吸收积累和细胞分布特点研究. 农业环境科学学报, 2010, (11): 2080-2086.
[10] Xu W H, Wang H X, Wang Z Y,etal. Effects of zinc, cadmium and their combined pollution on nutrient uptake and Zn, Cd accumulation in ryegrass (LoliumperenneL.). Asisn Journal of Ecotoxicology, 2006, (1): 70-74.
徐卫红, 王宏信, 王正银, 等. 锌、镉复合污染对重金属蓄集植物黑麦草养分吸收及锌、镉积累的影响. 生态毒理学报, 2006, (1): 70-74.
[11] Wu F Z, Yang W Q, Zhang J,etal. Effects of cadmium stress on growth and nutrient accumulation, distribution and utilization inOsmanthusfragransvar.thunbergii. Chinese Journal of Plant Ecology, 2010, (10): 1220-1226.
吴福忠, 杨万勤, 张健, 等. 镉胁迫对桂花生长和养分积累、分配与利用的影响. 植物生态学报, 2010, (10): 1220-1226.
[12] Liu S L, Shi X S, Pan Y Z,etal. Effects of cadmium stress on growth, accumulation and distribution of biomass and nutrient inCatharanthusroseus. Acta Prataculturae Sinica, 2013, 21(3): 154-161.
刘柿良, 石新生, 潘远智, 等. 镉胁迫对长春花生长, 生物量及养分积累与分配的影响. 草业学报, 2013, 21(3): 154-161.
[13] Zhang Q, Zeng X B, Su S M,etal. Arsenic uptake by different species of amaranth and the relationship between arsenic and phosphate in amaranth. Journal of Agro-Environment Science, 2016, 35(10): 1888-1894.
张骞, 曾希柏, 苏世鸣, 等. 不同品种苋菜对砷的吸收能力及植株磷砷关系研究. 农业环境科学学报, 2016, 35(10): 1888-1894.
[14] Xu P X. Studies on Cadmium Tolerance and Detoxification in Tall Fescue and Kentucky Bluegrass. Shanghai: Shanghai Jiaotong University, 2014.
徐佩贤. 高羊茅和草地早熟禾对镉的耐受能力和解毒机制研究. 上海: 上海交通大学, 2014.
[15] Shri M, Kumar S, Chakrabarty D,etal. Effect of arsenic on growth, oxidative stress, and antioxidant system in rice seedlings. Ecotoxicology and Environmental Safety, 2009, 72(4): 1102-1110.
[16] Ding F H, Liu S X, Luo D,etal. Arsenic toxicity to nineteen vegetable species in solution culture. Environmental Chemistry, 2010, (3): 439-443.
丁枫华, 刘术新, 罗丹, 等. 基于水培毒性测试的砷对19种常见蔬菜的毒性. 环境化学, 2010, (3): 439-443.
[17] Liu Q, Zheng C, Hu C X,etal. Effects of high concentrations of soil arsenic on the growth of winter wheat (TriticumaestivumL.) and rape (Brassicanapus). Plant Soil Environment, 2012, 58(1): 22-27.
[18] Li F Y, Qu X R, Wu L H. The Theoretical Basis and Technology of Bioremediation for Contaminated Soil. Beijing: Chemical Industry Press, 2006: 60.
李法云, 曲向荣, 吴龙华. 污染土壤生物修复理论基础与技术. 北京: 化学工业出版社, 2006: 60.
[19] Liu Q J, Sun X C, Hu C X,etal. Growth and photosynthesis characteristics of wheat (TriticumaestivumL.) under arsenic stress condition. Acta Ecologica Sinica, 2009, (2): 854-859.
刘全吉, 孙学成, 胡承孝, 等. 砷对小麦生长和光合作用特性的影响. 生态学报, 2009, (2): 854-859.
[20] Lefevre I, Vogel-Mikus K, Jeromel L,etal. Differential cadmium and zinc distribution in relation to their physiological impact in the leaves of the accumulatingZygophyllumfabagoL. Plant Cell and Environment, 2014, 37(6): 1299-1320.
[21] Baxter I, Dilkes B P. Elemental profiles reflect plant adaptations to the environment. Science, 2012, 336: 1661-1663.
[22] Gao F F. The Effects of Cu, Zn, Cd and Pb on the Growth of Three Leguminous Plants and the Metal-absorption Abilities of the Plants. Shenyang: Northeastern University, 2008.
高菲菲. Cu、Zn、Cd、Pb对三种豆科植物生长的影响及其吸附性能的研究. 沈阳: 东北大学, 2008.
[23] Cai C X, Wang H M, Zhang Z M. Studies on beneficiation mechanism and speed rate of removal of five heavy metalic ions by water gourd. Journal of Youjiang Teachers College for Nationlities Guangxi, 2002, (6): 48-51.
蔡成翔, 王华敏, 张宗明. 水葫芦对五种重金属离子的去除速率与富集机制研究. 广西右江民族师专学报, 2002, (6): 48-51.
[24] Li X M. Tolerance and Accumulation Characteristics of Cadmium in Herbaceous Plants. Beijing: Beijing Forestry University, 2016.
李希铭. 草本植物对镉的耐性和富集特征研究. 北京: 北京林业大学, 2016.
[25] Finnegan P M, Chen W. Arsenic toxicity: the effects on plant metabolism. Frontiers in Physiology, 2012, 3: 182.
[26] Verbruggen N, Hermans C, Schat H. Mechanisms to cope with arsenic or cadmium excess in plants. Current Opinion in Plant Biology, 2009, 12(3): 364-372.
[27] Chang S M, Ma X M, Zhang G L,etal. Effects of arsenic toxicity on carbon and nitrogen metabolism and the yield and quality of flue-cured tobacco. Journal of Plant Ecology, 2006, (4): 682-688.
常思敏, 马新明, 张贵龙, 等. 砷对烤烟碳氮代谢及其产量和品质的影响. 植物生态学报, 2006, (4): 682-688.
[28] Cakmak I. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. Journal of Plant Nutrition and Soil Science, 2005, 168: 521-530.
[29] Liu Q J, Sun X C, Tan Q L,etal. Effect of As on the concentration of nutrient elements and root growth of winter wheat(TriticumaestivumL.). Journal of Anhui Agricultural Sciences, 2011, 39(32): 19695-19697.
刘全吉, 孙学成, 谭启玲, 等. 砷对冬小麦根系生长和养分吸收的影响. 安徽农业科学, 2011, 39(32): 19695-19697.
[30] Chen T B, Huang Z C, Huang Y Y,etal. The distribution characteristics of arsenic and nutrient element in the leaf ofPterisvittata. Science In China (Serices C: Life Sciences), 2004, (4): 304-309.
陈同斌, 黄泽春, 黄宇营, 等. 蜈蚣草羽叶中砷及植物必需营养元素的分布特点. 中国科学(C辑: 生命科学), 2004, (4): 304-309.
[31] Bhowrnik S, Datta B K, Saha A K. Determination of mineral content and heavy metal content of some traditionally important aquatic plants of tripura, India using atomic absorption spectroscopy. Journal of Agricultural Technology, 2012, 8(4): 1467-1476.
