南通沿海潮间带土-气界面CO2交换特征
2018-03-02王一丹王晨昕华祖林王胜艳河海大学环境学院江苏南京20098河海大学浅水湖泊综合治理与资源开发教育部重点实验室江苏南京20098南京水利科学研究院江苏南京20029水文水资源与水利工程科学国家重点实验室江苏南京20098江苏省水文水资源勘测局泰州分局江苏泰州22300
王 鹏,王一丹,申 霞,王晨昕,华祖林,2,王胜艳 (.河海大学环境学院,江苏 南京 20098;2.河海大学浅水湖泊综合治理与资源开发教育部重点实验室,江苏 南京 20098;3.南京水利科学研究院,江苏 南京20029;4.水文水资源与水利工程科学国家重点实验室,江苏 南京 20098;.江苏省水文水资源勘测局泰州分局,江苏 泰州 22300)
随着全球气候变暖,大气中的CO2和CH4等关键活动温室气体浓度在2011年分别增加到了391×10-6和1803×10-9,比工业化之前增加40%和150%[1].碳循环成为全球研究的热点问题.近年来,作为全球“蓝色碳汇”的主要贡献者,有着高生产力的河口海岸潮间带湿地的碳循环受到了极大的关注[2-3].潮间带是众多湿地的一种,尽管全球潮间带总面积比陆地森林要小1~2个数量级[4],但是其单位面积的碳封存能力却不容忽视[5-6].据统计,全球范围内滨海湿地的总碳封存速率超过100Tg/a (以碳计),因此潮间带湿地在减缓含碳温室气体排放,降低全球温室效应方面具有重要潜力[5].
目前,对滨海潮间带湿地碳通量的研究逐渐深入[3].大量研究表明,潮间带湿地土-气界面CO2通量具有显著的季节性变化特征.受温度的影响,夏季地下生物活跃,CO2排放速率较高,冬季生物活性低,CO2释放速率达到最低值[7-8].除季节性变化外,潮间带湿地CO2通量在一天之内也会呈现明显的波动过程[7],Van Der Nat等[9]发现湿地系统的CO2通量在一天内通常呈现单峰型变化的趋势,从早晨开始慢慢上升,在午后出现最大值后再缓慢下降.由于湿地碳通量的周期性变化,导致碳循环的输入和输出过程受季节因素的影响,沼泽湿地中的生产者在春季和夏季固定大量的碳,而在夏末和秋季,沼泽会向河口和海洋输送大量的碳源[10].
湿地土壤CO2通量包含吸收和呼吸两部分[11].吸收部分是土壤自养微生物同化CO2过程,释放部分主要是土壤微生物代谢有机物及植物根际呼吸作用[12],因此土壤碳通量容易受自然环境影响,包括水文条件、温度、土壤有机质含量、土壤生物等[13].宋长春等[14]研究发现,潮间带湿地碳通量与土壤温度呈正相关关系,温度升高,土壤呼吸速率加强[15],同时促进盐沼植物的生长,使湿地固碳能力增强[16-17].水位的变化通过潮汐的周期性淹水实现,从而影响土壤呼吸和植物生长分布,影响碳循环过程[18-19].潮间带湿地CO2通量与地下水水位呈负相关关系, Freeman等[20]研究威尔士某泥炭地,发现水位下降,CO2通量从646mg/(m2·d)增加到1590mg/(m2·d).
我国有1.8万km的海岸线,仅高程在海图0m以上的潮间带就有2.17×104km2,相当于我国1/4的湖泊湿地面积[21].江苏海岸线长约954km,沿海滩涂湿地面积约占全国沿海滩涂湿地总面积的1/4.1987~2007年,整个江苏省沿海滩涂湿地土地资源的组成发生了显著的变化.耕地、建设和居民用地以及人工湿地面积比例持续增加,米草沼泽、碱蓬沼泽和芦苇沼泽为代表的自然湿地面积持续显著减少,主要转化为养殖用地、盐田和耕地,导致无植被覆盖的光滩面积逐年增加,土-气界面CO2交换特征发生了很大的变化[22].此外,大部分针对河口湿地碳循环的研究是将湿地植被与土壤碳通量合并作为生态系统整体加以考虑,然而土地覆被的改变导致江苏沿海潮间带大部分区域形成光滩,因此研究无植被覆盖下的土壤碳循环是揭示该地区湿地生态系统碳循环的基础,是研究局地气候变化的关键,在相关领域开展深入研究显得尤为迫切.本文选择长江口北侧启东沿海光滩潮间带作为研究对象,采用现场观测、室内实验和机理分析相结合的方法,阐明了潮间带土-气界面的CO2交换特征,揭示了碳循环的影响因素及其作用机制.研究成果为预测及评估潮间带土壤碳循环对滨海湿地生态系统的影响提供了科学依据,为受损潮间带湿地碳汇功能保育和修复措施制定提供了数据支撑.
1 材料与方法
1.1 研究地区概况
研究区域选在江苏省南通市海岸带,其位于江苏省东南部、长江口以北、东临黄海,见图1.
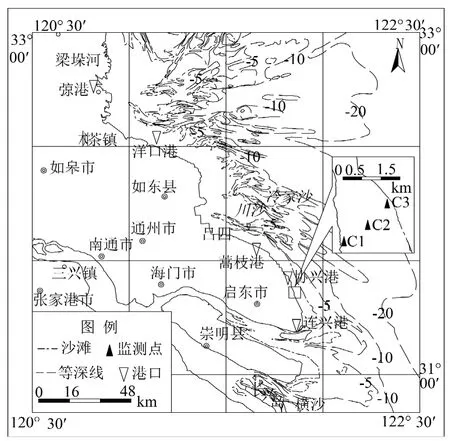
图1 研究区域位置Fig.1 The location of the study area
由于岸外辐射沙脊群的掩护,南通沿海多为淤涨型潮滩[22],沿岸地形低平,潮间带浅滩宽8~10km,其中潮上带宽约2km,潮间带宽4~6km,中潮位以下为粉砂带.南通近海岸湿地约36.6万hm2,潮滩呈明显的分带性,自陆向海依次为草滩带、盐蒿泥滩带、泥-砂混合带及粉砂滩带.根据南通海岸潮间带的地形地貌、潮波动力、泥沙沉积、生物组成和分布等特征,拟选择受人类活动和底栖动物扰动相对较小、土壤有机质含量相对较高的盐蒿泥滩带光滩土壤为研究对象.
1.2 实验设计及样品采集
选择启东协兴港附近潮间带(31°47'19"N,121°54'16"E),进行土-气界面碳通量现场观测.于2017年4月沿垂直海岸线方向在高潮带、中潮带、低潮带分别布设监测点C1、C2、C3,见图1.落潮之后,采用WEST便携式土壤通量测量系统进行土-气界面碳通量观测.以高潮带、中潮带和低潮带的顺序,测量各测点处土-气界面的CO2浓度变化过程,每次测量连续采样4min,采样频率为1次/s,每个测点重复测量5次.
同时采集监测点处土壤垂向分层样本,每层5cm,共5层(表层至底层依次编号为:1、2、3、4、5),置于洁净的封口袋中,于-18℃冷冻保存.分析测定土壤颗粒级配、土壤总有机碳(TOC)及叶绿素a含量.
1.3 样品分析
1.3.1 颗粒级配 样品进行冷冻干燥,捣碎研磨后,采用美国贝克曼库尔特有限公司生产的全自动激光粒度分析仪(LS13320),进行土样粒度分析.
1.3.2 总有机碳 取适量冻干土样,经酸化前处理去除杂质,于65℃烘干后,采用燃烧氧化-非分散红外法(Multi N/C2100)检测土壤中的有机碳含量.
1.3.3 叶绿素a 对冷冻保存的样品解冻,取2g左右放入150mL磨口具塞碘量瓶中,加入9mL 90%的丙酮及少量碳酸镁,塞紧瓶塞,避光超声15min,随后置于冰箱冷藏室萃取24h,经冷冻离心15min(转速4000r/min)后[23],按《海洋调查规范》(GB 12763-2007)荧光法测量土壤中的叶绿素a含量[24].
1.4 数据处理
使用WEST便携式土壤通量测量系统进行原位监测,该系统由红外分析仪、空气泵、掌上电脑及土壤呼吸气室组成,其中CO2传感器采用LI-COR公司生产的LI-840A.
对CO2浓度监测结果进行线性回归分析,按式(1)计算土-气界面CO2浓度的时间变化速率.计算公式如下:
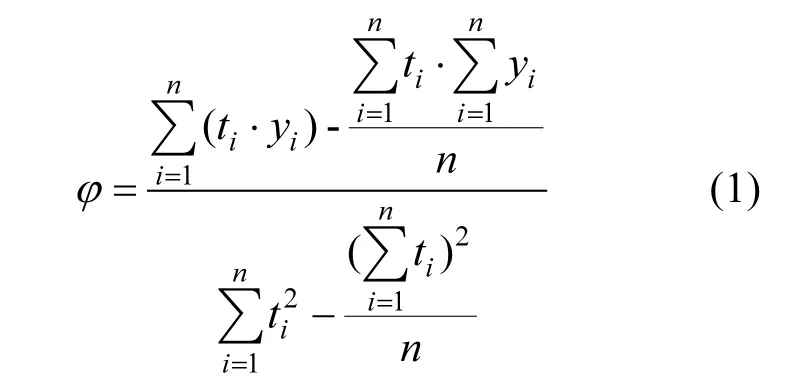
式中:ϕ为CO2浓度变化速率,10-6/s;ti为第i个监测数据对应的监测时间,s;yi为第i个监测数据的CO2浓度值,10-6;n为监测数据的数量.
为了衡量CO2浓度观测结果的数据质量,采用公式(2)计算监测数据的线性回归偏差.当偏差大于0.5,表示实验数据基本可靠,大于0.9表示实验数据质量较好,小于0.5表示实验数据质量较差.
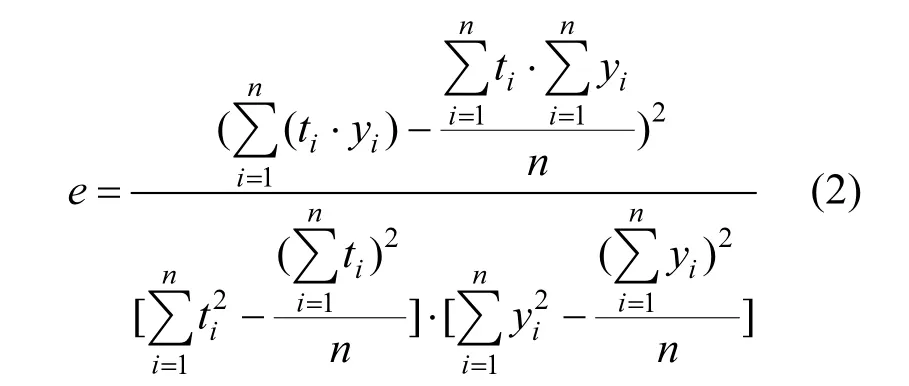
式中:e为线性回归偏差.
采用公式(3)可将CO2浓度变化速率转换成CO2通量,转化因子k采用公式(4)计算.
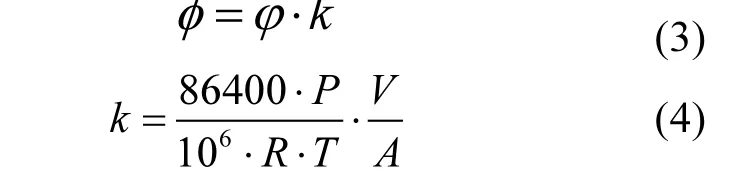
式中:Φ为CO2通量,mol/(m2·d);k为转化因子;P为空气气压,hPa;T为空气温度,K;V为气室体积,m3;A为气室入口处面积,m2;R为理想气体常数,J/(mol·k).
2 结果与讨论
2.1 土壤理化性质分析
2.1.1 颗粒级配 将监测点C1、C2、C3分层采集的样品,使用激光粒度仪进行分析,将各监测点的各层样品的平均粒径绘制柱形图,如图2所示.
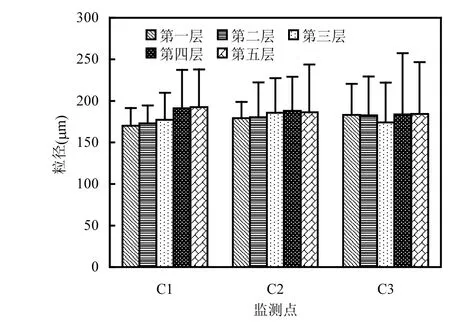
图2 土壤平均粒径垂向分布图Fig.2 The vertical distribution of average soil particle size
从图2可以看出,各监测点平均粒径水平相当,各点之间差距较小,整体粒径均值范围为170~190μm.C1、C2从表层至深层,粒径逐渐增大,C3点第三层粒径较小,其它层粒径水平相当.可见,越靠近岸边,潮流对泥沙的分选作用越明显.C1、C2和C3表层颗粒相对较为细小,这是由于潮汐作用,带来粒径较细的泥沙,表层颗粒粒径对沉积物有一定的影响[25],绘制各监测点表层粒径颗粒级配曲线图,如图3所示,横坐标为对数坐标,纵坐标小于某粒径的累积百分比.
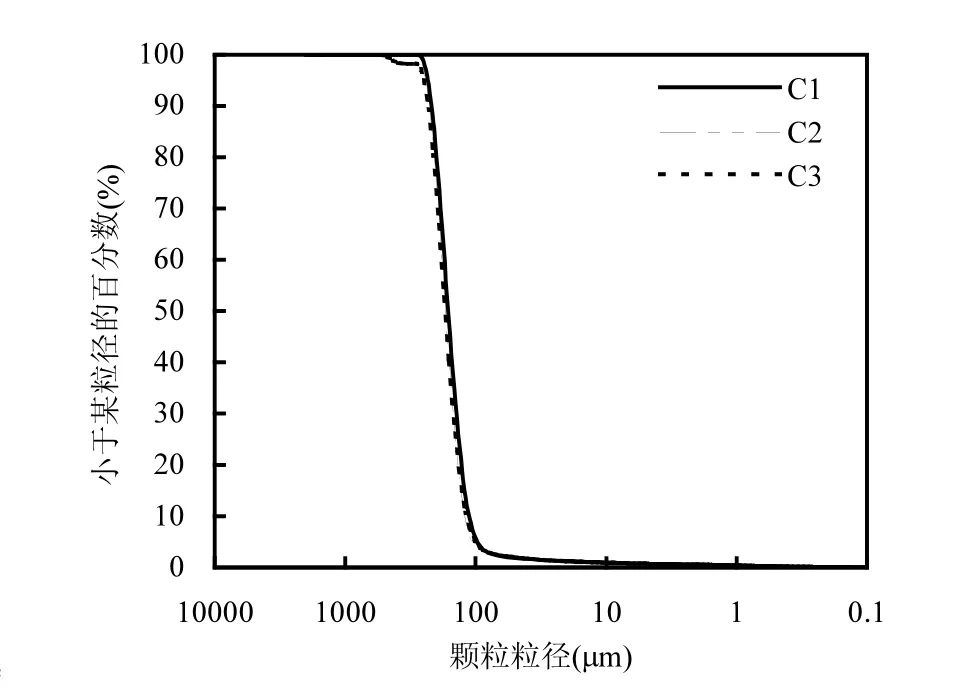
图3 表层土壤颗粒级配曲线Fig.3 The grading curves of the top soils
研究区域表层粒径主要分布在100~400μm之间,整体粒度在同一数量级水平,各监测点的粒度曲线几乎重合,相差较小,监测点位颗粒级配曲线较陡,粒径变化范围小,组成均匀.根据粒度分析手册对样品进行分类,见表1.

表1 样品理化特征分析Table 1 Physicochemical characteristics of soil samples
从表1可以看出该潮滩为粉砂质砂,监测点的沉积物中值粒径与平均粒径接近,说明粒径分布较为均匀[26].淤泥质潮滩沉积物通常为黏性细颗粒泥沙,潮滩沉积物的粒度分布可反映泥沙的沉积过程.从高潮滩到低潮滩,泥沙颗粒逐渐变粗,并且泥沙平均粒径夏季大于冬季,具有明显的季节性变化特征[27].这种变化会影响叶绿素的分布,当泥沙颗粒较为紧实时,使得光线无法透入深层,叶绿素a浓度显著下降[28].
2.1.2 TOC分析 各潮带土壤总有机碳含量的垂向分布特征如图4所示.由图4可见,高潮带表层土壤的总有机碳含量大于中潮带和低潮带,呈现由海岸向海域逐渐递减的趋势.从垂直方向上看,各监测点的第二层TOC有明显锐减的趋势.目测发现第二层样品颜色较深,这可能与人类养殖活动、水生生物活动、水动力扰动等因素有关.深层的TOC呈缓慢增加至稳定的规律,但规律性并不明显,可能是因为影响有机碳含量的原因多样、生物和物理搅动等因素影响[29].研究发现,随着土壤深度增加,深层土壤中可利用的有机物含量降低,而稳定有机碳比例较高[30].
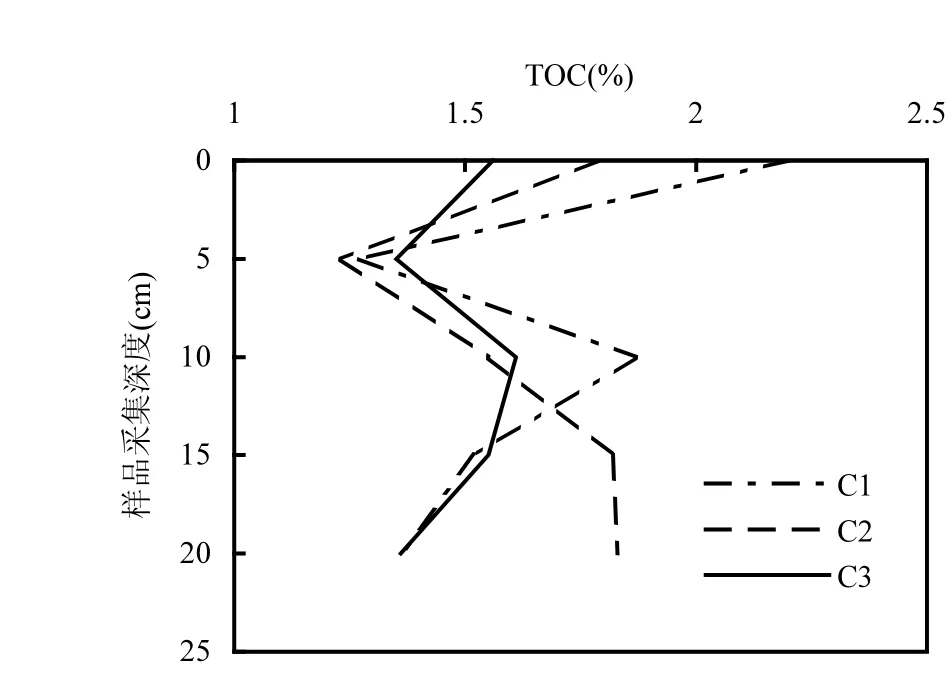
图4 各潮带土壤总有机碳含量Fig.4 Total organic carbon contents of soils in the different tidal zones
2.1.3 叶绿素a含量 研究区域为光滩,无植被覆盖.研究表明,该地区土壤中叶绿素a主要分布在表层1cm内[29],因此本研究仅测量表层叶绿素a含量,分析结果见表1.
该潮滩泥沙颗粒较细,具有较强吸附能力,为底栖微藻提供丰富的营养物质[31].由表1可见,潮滩各测点叶绿素a含量较高,介于7~8μg/g,这主要是因为春季潮滩属于快速淤积期,表层沉积物营养较丰富[32].此外,叶绿素a含量:高潮带<中潮带<低潮带.低潮带人为活动较少,受潮汐过程影响较大,涨潮过程中携带的大量营养盐,为表层沉积物中藻类的生长提供良好的环境[33].藻类生长与潮滩淤积之间存在相互促进的关系[28].观测期间潮汐作用携带大量泥沙,细颗粒物含量较高,有利于藻类的生长,因此叶绿素a含量较高.
2.2 潮间带CO2通量特征及影响因素分析
2.2.1 潮间带土-气界面CO2浓度及通量变化特征 统计潮间带各监测点各测次土-气界面CO2浓度的平均值和标准差,将重复监测每点计算所得CO2平均浓度绘制图形如图5所示.
从图中可以看出,C1点平均CO2浓度值在312×10-6~335×10-6之间,C2点的平均CO2浓度在300×10-6~310×10-6之间,C3点的平均CO2浓度在295×10-6~315×10-6.各监测点CO2浓度呈现下降趋势,表示潮滩进行CO2固定过程.C1点CO2浓度值大于C2和C3点,C1点为高潮带区,靠近陆地,更接近于人类活动区域,空气中CO2浓度本底值较高.C2和C3点浓度水平相当,中潮带和低潮带远离海岸,人类活动和干扰较小.对各监测点CO2浓度监测数据的标准差进行统计后发现,C1点监测数据最为稳定,CO2浓度随时间的变化幅度较小.
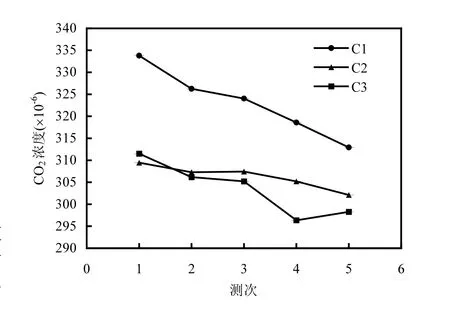
图5 潮间带土-气界面CO2浓度变化Fig.5 Variation in CO2 concentrations of intertidal zone at soil-air interface
将CO2浓度带入公式(1)和(3),计算土-气界面的CO2净通量,采用公式(2)计算CO2浓度的线性回归偏差.偏差计算结果表明,各测点回归偏差值介于0.72~0.99之间,说明碳通量观测结果的数据质量良好.为了便于对净通量进行分析,在本研究中定义土-气界面的CO2净通量等于固定通量减去释放通量.图6为土-气界面CO2净通量与TOC的关系图,其中各监测点CO2净通量用箱线图表示,折线图表示各监测点TOC含量.其中箱体上下边缘分别表示75%和25%的数值,箱体中间横线为中值,小正方形为平均值,箱线上下边缘代表最大值和最小值.
由图6可见,各测点各测次的CO2净通量均为正值,这意味着CO2固定通量大于释放通量,说明潮间带光滩湿地对CO2整体表现为固定作用.CO2净通量范围为0.004~0.035mol/(m2·d),各监测点通量值存在差异.高潮带(C1)和中潮带(C2)的CO2净通量整体上大于低潮带(C3).此外,各测点CO2净通量平均值均大于中值,说明通量呈现偏态分布,观测数据整体偏向于较大值. Heinsch等[34]对北美38个不同湿地类型4a CO2通量数据进行相关性分析发现,土壤含水率高、土壤盐度较低情况下,潮间带湿地表现为固定CO2,杨红霞等[33]在对崇明东滩潮间带沉积物-大气间碳通量进行研究,发现潮滩表现对中潮滩和低潮滩都表现为CO2的吸收汇,其中中潮滩在光照条件下CO2吸收量为-67.45mg/(m2·h),各潮滩CO2吸收量存在差异,与本研究的观测结果相符.
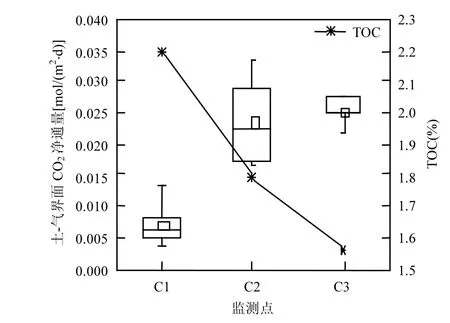
图6 土-气界面CO2净通量与TOC的关系Fig.6 The relationship between CO2 net fluxes at soil-air interface and TOC
影响碳通量的因素多样,研究表明,植被分布和土壤有机质含量是影响碳通量的重要原因,有机质含量高,营养丰富,因此藻类生物较为生长迅速,CO2通量有明显的变化规律.此外,还与土壤叶绿素含量、光照条件、地下水位的变化有关[35-36].
2.2.2 潮间带碳通量与土壤有机碳含量及叶绿素a的关系 土壤-大气之间的碳通量取决于土壤中有机碳的输入和输出速率[37],本研究区为潮间带裸露光滩,土壤CO2交换过程主要包括吸收和释放CO2,吸收部分主要来自自养微生物同化CO2,释放部分主要包括微生物的呼吸作用,另有少部分来自土壤动物的呼吸和化学氧化[38].本研究中土壤微生物的呼吸作用是CO2释放的主要途径,与微生物的代谢作用有关.图6中不同各监测点对应不同的有机碳含量,由图可见,不同TOC水平下,各监测点土-气界面CO2净通量存在显著差异.总体而言,净通量随土壤有机碳含量的增加而减小,并且CO2净通量随土壤TOC含量上升快速下降.这主要是因为潮间带土-气界面CO2通量由固定和释放两方面因素决定.
研究表明,土壤有机碳是微生物代谢的主要来源.在相同条件下,土壤有机碳含量越高,土壤呼吸速率越高,CO2释放通量越大[39].从TOC含量关系来看,C1>C2>C3,说明C1土壤呼吸速率较高,因此,CO2释放通量较大.
土壤对CO2固定作用主要是由土壤所含叶绿素a的含量和光照强度决定.由于各监测点距离较近,监测时间差别较小,故可认为监测时段内各监测点的光照强度近似相等.各监测点叶绿素a含量关系为:C1<C2<C3,由于光合作用速率与叶绿素a含量成正比,可以推断3个测点的CO2固定通量的大小关系为:高潮带<中潮带<低潮带.因此,CO2净通量随土壤TOC含量上升而下降.
2.2.3 落潮时间对潮间带碳通量的影响 由前述分析可以发现,同一监测点不同测次的碳通量存在明显差异,由于本研究监测时段内的土壤温度变化较小,因此,不同测次碳通量的差异性应该与落潮导致的地下水位变化有很大关系[1].
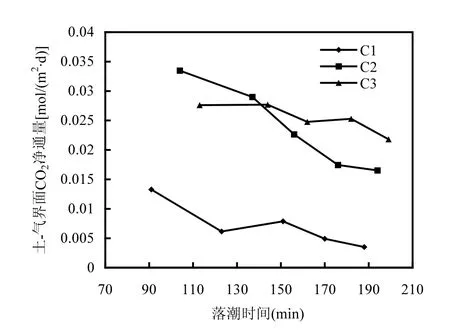
图7 土-气界面CO2净通量与落潮时间的关系Fig.7 The relationship between CO2 net fluxes at soil-air interface and the ebb durations
图7为各潮带土-气界面CO2净通量与落潮时间的关系.由图可见,随着落潮时间的增长,观测点地下水位逐渐降低,潮间带CO2净通量呈现明显下降趋势.这主要是随着落潮时间增长,地下水位不断降低,土壤微生物呼吸作用逐渐加强,呼吸速率提高,CO2释放量增大[34].由于监测时段较短,土壤中叶绿素a含量变化很小,潮间带土壤对CO2的固定通量几乎不变,因此,土-气界面CO2净通量呈现逐渐降低的趋势.
马安娜[40]对长江崇西湿地CO2交换的研究也发现,净光合作用固碳过程受潮汐的影响较小,潮汐对生态系统CO2交换过程主要制约土壤呼吸释放CO2过程,即潮汐淹没抑制了土壤呼吸及CO2传输释放.这主要是因为天然滨海湿地受潮汐影响的周期性淹水会使土壤处于饱和状态,土壤呼吸速率处于较低水平[41],提高水位可降低土壤呼吸速率[42].较高的水位会造成厌氧环境,不利于微生物的好氧呼吸,会造成CO2释放减少[43].
通过分析各潮带CO2净通量的关系发现,低潮带(C3)碳通量下降过程较平缓,而高潮带(C1)和中潮带(C2)的CO2通量下降趋势相对较快,这可能与观测点处地下水位的下降速率有关.高潮带(C1)和中潮带(C2)靠近岸边,地下水水位随落潮时间的下降速率较快,CO2净通量随时间呈显著变化.低潮带(C3)远离海岸,其地下水水位受落潮的影响相对滞后,水位下降速率相对较慢,因此,CO2净通量随时间变化不显著.此外,各潮带CO2净通量呈现前期快速降低,后期缓慢下降的趋势,这与其地下水水位变化也是密切相关的.落潮前期地下水水位迅速下降,各潮带对CO2的固定速率下降迅速,后期地下水水位变化逐渐放缓,CO2净通量也逐渐趋于稳定.
3 结论
3.1 研究区潮间带表层为细微颗粒.高潮带表层土壤TOC含量为2.2%,大于中潮带(1.78%)和低潮带(1.57%),呈现由海岸向近海逐渐递减的趋势.表层土壤叶绿素a含量空间分布与TOC呈相反变化趋势.
3.2 研究区潮间带光滩土-气界面CO2浓度随时间呈下降趋势,表现为CO2的吸收汇,净固定CO2量在0.004~0.035mol/(m2·d).高潮带和中潮带的CO2净通量大于低潮带,这与土壤有机碳和叶绿素a含量以及潮汐引起的地下水位变化密切相关.
3.3 CO2净通量随落潮时间呈现明显下降趋势.随着落潮时间增长,地下水位不断降低,土壤微生物呼吸作用加强,CO2释放量增大,净固定CO2能力减弱.
[1] Yamochi S, Tanaka T, Otani Y, et al. Effects of light, temperature and ground water level on the CO2flux of the sediment in the high water temperature seasons at the artificial north salt marsh of Osaka Nanko bird sanctuary [J]. Ecological Engineering, 2017,98:330-338.
[2] Chmura G L, Anisfeld S C, Cahoon D R, et al. Global carbon sequestration in tidal, saline wetland soils [J]. Global Biogeochemical Cycles, 2003,17(4):1-12.
[3] Moffett K B, Wolf A, Berry J A, et al. Salt marsh atmosphere exchange of energy, water vapor, and carbon dioxide: Effects of tidal flooding and biophysical controls [J]. Water Resources Research, 2010,46(10):5613-5618.
[4] Kirman M L, Megonigal J P. Tidal wetland stability in the face of human impact and sea-level rise [J]. Nature, 2013,504(7478):53-60.
[5] Hopkinson C S, Cai W J, Hu X, et al. Carbon sequestration in wetland dominated coastal systems a global sink of rapidly diminishing magnitude [J]. Current Opinion in Environmental Sustainability, 2012,4(2):186-194.
[6] Donato D C, Kauffman J B, Murdiyarso D, et al. Mangroves among the most carbon-rich forests in the tropics [J]. Nature Geoscience, 2011,4(5):292-297.
[7] Kurganova I N, Rozanova L N, Myakshina T N, et al. Monitoring of CO2emission from soils of different ecosystems in the southern Moscow region: Analysis of long-term field studies [J].Eurasian Soil Science, 2004,37:S74-S78.
[8] Morris J T, Whiting G J. Emission of gaseous carbon dioxide from salt-marsh sediments and its relation to other carbon losses[J]. Estuaries, 1986,9(1):9-19.
[9] Van Der Nat W A, Middelburg J J, Meteren D V, et al. Diel methane emission patterns from Scirpus lacustris and Phragmites australis [J]. Biogeochemistry, 1998,41(1):1-22.
[10] Wang Z A, Cai W J. Carbon dioxide degassing and inorganic carbon export from a marsh-dominated estuary (the Duplin River): A marsh CO2pump [J]. Limnology Oceanography, 2004,49(2):341-354.
[11] 王宗林,吴沿友,刑德科,等.泉州湾红树林湿地土壤CO2通量周期性变化特征 [J]. 应用生态学报, 2014,25(9):2563-2568.
[12] 刘绍辉,方精云.土壤呼吸的影响因素及全球尺度下温度的影响[J]. 生态学报, 1997,17(5):469-476.
[13] 曹 娜,何春光,罗文泊,等.冬季湿地二氧化碳通量研究进展[J]. 湿地科学, 2015,13(2):244-251.
[14] 宋长春,阎百兴,王跃思,等.三江平原沼泽湿地CO2和CH4通量及影响因子 [J]. 科学通报, 2003,48(23):2473-2477.
[15] Davidson E A, Trumbore S E, Amundson R. Soil warming and organic carbon content [J]. Nature, 2000,408(6814):789-790.
[16] Bertness M D, Ewanchuk P J, Silliman B R. Anthropogenic modification of New England salt marsh landscapes [J].Proceedings of the National Academy of Sciences of the United States of America, 2002,99(3):1395-1398.
[17] Charles H, Ducks J S. Effects of warming and altered precipitation on plant and nutrient dynamics of a New England salt marsh [J]. Ecological Applications, 2009,19(7):1758-1773.
[18] Ewanchuk P J, Bertness M D. The role of waterlogging in maintaining forb pannes in northern New England salt marshes[J]. Ecology, 2004,85(6):1568-1574.
[19] Barr J G, Engel V, Fuentes J D, et al. Controls on mangrove forest-atmosphere carbon dioxide exchanges in western Everglades National Park [J]. Journal of Geophysical Research,2010,115(G2):1-14.
[20] Freeman C, Lock M A, Reynolds B. Fluxes of CO2, CH4and N2O from a welsh peatland following simulation of water table drawdown: potential feedback to climatic change [J]. Biogeochemistry,1993,19(1):51-60.
[21] 杨朋金.裸露潮间带表观CO2通量变化规律 [D]. 青岛:中国海洋大学, 2009.
[22] 黄 辉,钱 宜,顾进伟.江苏沿海滩涂湿地景观变化及环境效应 [J]. 湖北农业科学, 2014,53(4):822-825.
[23] 王丹丹.江苏如东海岸互米花草盐沼沉积物叶绿素a分布特征[J]. 生态学杂志, 2012,31(9):2247-2254.
[24] GB12763-2007 海洋调查规范 [S].
[25] 杨世伦,时 钟,赵庆英.长江口潮沼植物对动力沉积过程的影响 [J]. 海洋学报, 2001,23(4):75-80.
[26] 邵学军,王兴奎.河流动力学概论 [M]. 北京:清华大学出版社,2005:23-25.
[27] 刘清玉,戴雪融,何小勤.崇明东滩沉积环境探讨 [J]. 海洋地质动态, 2003,19(12):1-4.
[28] 李万会.潮滩湿地沉积物中叶绿素a浓度的变化特征及其与沉积物特征性间的关系初探 [D]. 上海:华东师范大学, 2006.
[29] 尹胜乐,刘晓收,袁 超,等.胶州湾大沽河潮间带沉积物中叶绿素和有机质含量季节变化 [J]. 海洋湖沼通报, 2012,2:97-106.
[30] 周艳翔,吕茂奎,谢锦升,等.深层土壤有机碳的来源、特征与稳定性 [J]. 亚热带资源与环境学报, 2013,8(1):48-55.
[31] 李万会,丁平兴.滩涂沉积物中叶绿素a浓度与沉积特性的关系[J]. 华东师范大学学报(自然科学版), 2007,4:26-33.
[32] 杨世伦.长江三角洲潮滩季节性冲淤循环的多因子分析 [J].地理学报, 1997,52(2):123-130.
[33] 杨红霞,王东启,陈振楼,等.长江口潮滩湿地-大气界面碳通量特征 [J]. 环境科学学报, 2006,26(4):667-673.
[34] Heinsch F A, Zhao M, Running S W, et al. Evaluation of remote sensing based terrestrial productivity from MODIS using regional tower eddy flux network observations [J]. IEEE Transactions on Geoscience & Remote Sensing, 2006,44(7):1908-1925.
[35] 王文杰,于景华,毛子军,等.森林生态系统CO2通量的研究方法及研究进展 [J]. 生态学杂志, 2003,22(5):102-107.
[36] 马安娜,陆健健.湿地生态系统碳通量研究进展 [J]. 湿地科学,2008,6(2):116-123.
[37] Blanca B, Mitsch W J. A comparison of soil carbon pools and profiles in wetlands in Costa Rica and Ohio [J]. Ecological Engineering, 2008,34(4):311-323.
[38] 翟生强,史长光,杜乐山,等.若尔盖泥炭地地下水位和土壤温度对二氧化碳排放的影响 [J]. 湿地科学,2015,13(3):332-337.
[39] Ahn M Y, Zimmerman A R, Comerford N B, et al. Carbon mineralization and labile organic carbon pools in the sandy soils of a North Florida watershed [J]. Ecosystems, 2009,12(4):672-685.
[40] 马安娜,陆健健.长江口崇西湿地生态系统的二氧化碳交换及潮汐影响 [J]. 环境科学研究, 24(7):716-721.
[41] 仲启铖.温度和水位对滨海围垦湿地碳过程的影响-以崇明东滩为例 [D]. 上海:华东师范大学, 2013.
[42] Liveseley S J, Andrusiak S M. Temperate mangrove and salt marsh sediments are a small methane and nitrous oxide source but important carbon store [J]. Estuarine Coastal & Shelf Science,2012,97(2):19-27.
[43] Erwin K L. Wetlands and global climate change: the role of wetland restoration in a changing world [J]. Wetlands Ecology &Management, 2009,17(1):71-84.
