Effects of dietary forage to concentrate ratio and wildrye length on nutrient intake, digestibility, plasma metabolites, ruminal fermentation and fecal microflora of male Chinese Holstein calves
2018-02-05XlAChuanqiAzizUrRahmanMuhammadNlUWenjingSHAOTaoqiQlUQinghuaSUHuaweiCAOBinghai
XlA Chuan-qi, Aziz-Ur-Rahman Muhammad, NlU Wen-jing SHAO Tao-qi QlU Qing-hua SU Huawei CAO Bing-hai
1 State Key Laboratory of Animal Nutrition, College of Animal Science and Technology, China Agricultural University, Beijing 100193, P.R.China
2 Institute of Animal and Dairy Sciences, University of Agriculture Faisalabad, Faisalabad 3800, Pakistan
1. lntroduction
Increasing demand for beef in recent years is motivating Chinese farmers to introduce dairy bull calves into the beef industry to fulfill the beef requirements of the large Chinese population (Muhammadet al. 2016). To obtain maximum growth and higher profitability, high concentrate diets along with a forage source are usually used.
The forage to concentrate ratio (F:C) depends on dietary ingredients and nutritional requirements of ruminant animals.Small-scale farmers may alter the forage to concentrate ratio without knowing that it has negative consequences on the physiological system of the animal. It has been reported that forage level affects nutrient intake, utilization,and digestibility (Zebeliet al. 2006; Yang and Beauchemin 2007; Granja-Salcedoet al. 2016). Studies have also determined the influence of forage length (FL) on voluntary nutrient intake and apparent total tract nutrient digestibilities(Mooney and Allen 1997; Kononoff and Heinrichs 2003;Al-Saiadyet al. 2010). It is also well established that the physical form of dietary forage plays important roles in the development and performance of the rumen in dairy calves(Montoroet al. 2013); inadequate dietary fiber or lack of fiber effectiveness reduces chewing time of ruminants, which may lead to reduced saliva secretion and lower rumen pH(Erdman 1988). However, the literature on growing calves,especially Chinese Holstein calves, is limited.
Previous research has shown that normal microflora plays an important protective and nutritional role in the gastrointestinal tract (Conway 1994). The gastrointestinal tract of young animals is vulnerable and can be easily invaded by pathogenic microorganisms, which can ultimately cause microbial flora imbalance (Ofeket al.1975). Regulation of the numbers and species of beneficial microflora in the intestinal tract can be controlled by diet(Reid and Hillman 1999; Snelet al. 2002). Thomlinson and Lawrence (1981) demonstrated that dietary fiber could inhibit the proliferation ofEscherichia coli, and effectively improve intestinal health. Results of another study indicated that a large amount of unfermented carbohydrate in the back end of the digestive tract might increase osmotic pressure and lead to excessive water secretion, ultimately causing diarrhea (Radecki and Yokoyama 1991). However, there is less research focus on the effects of dietary FL and F:C on intestinal microorganisms in calves.
Therefore, the objective of this trial was to determine the effects of F:C and FL on nutrient intake, digestion, plasma metabolites, ruminal fermentation, and fecal microflora of male Chinese Holstein calves.
2. Materials and methods
The current study was performed in Zhuozhou City in Hebei Province, China. All procedures were approved for animal welfare following instructions of the China Council on Animal Care. The experiment was authorized by the Animal Care and Use Committee of the China Agricultural University.
2.1. Animals, experimental design, and diets
Twenty-eight male Holstein calves (approximately 4–5 mon old) weighing (156.8±33.4) kg (mean±SD) were used in a completely randomized design with four treatments in 2×2 factorial design. Factors were FL and F:C. Each treatment contained seven replicates of one calve per replicate.Dietary FL levels were designated as whole-length (WL)and short-length (SL). At each FL level the calves were fed diets with one of two F:C levels: low F:C (50:50) and high F:C (65:35). Chinese wildrye was used as the only forage source in the present study. Chopped hay was obtained using a machine (STORTI HUSKY, 9SJW-500, Italy) set at 1 900 r min–1to achieve short particle sizes (~50% particles>19 mm). Ingredients and composition of experimental diets are given in Table 1. Animals were fed the total mixed ration twice daily at 0800 h and 1700 h and the duration of the experiment was 2 mon. Experimental calves were individually reared, and offered feedad libitum(5% residual was permitted).
2.2. Nutrient intake and apparent total tract nutrient digestibility
Feed intake data were collected daily. Feed and residual samples were collected for further analysis in the laboratory.At the end of the experiment, a digestion trial was conducted for 3 d. Feces were mixed, weighed, and subsampled twice daily. Approximately 200 g samples were collected,and tartaric acid (10%) was added at 5% fresh weight, and then the samples were dried at 65°C for 48 h. Total tract digestibility was calculated based on the differences in nutrient concentration between feces and diets.
Before chemical analyses, feed and feces samples were ground using a 1-mm sieve. Dry matter (DM) and crude ash content of all samples were determined according to the Association of Official Analytical Chemistry (AOAC 2000). Organic matter (OM) concentration was calculated as the difference between DM and crude ash content.Crude protein (CP) was estimated by N×6.25, which was determined according to AOAC (2000) (Leco FP-528 Nitrogen Analyzer; Leco Corp., St. Joseph, MI, USA).Neutral detergent fiber (NDF) and acid detergent fiber (ADF)were measured using heat-stable α-amylase (Van Soestet al. 1991) and Na2SO3(Hintzet al. 1996).
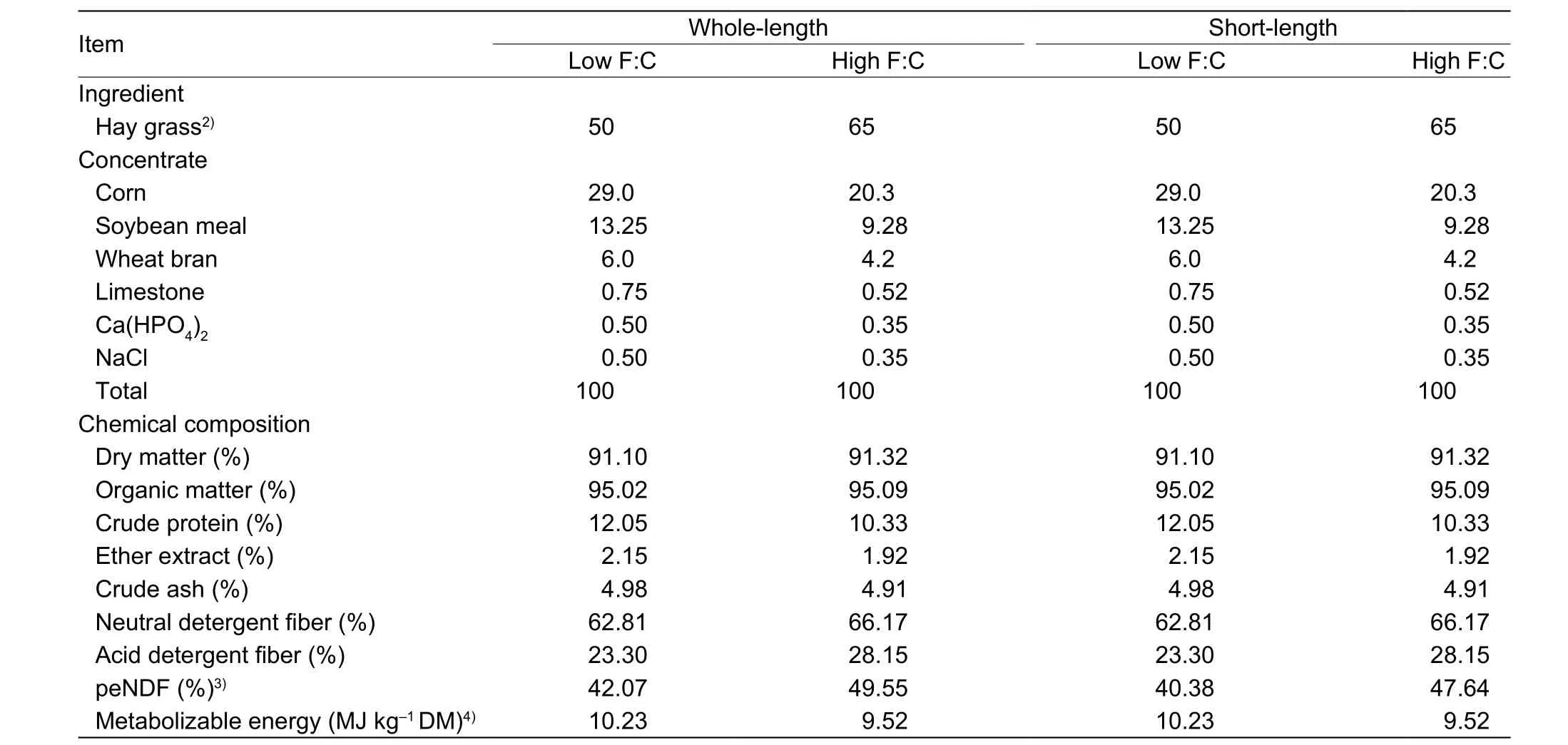
Table 1 Ingredient and chemical composition of the basal diet (DM basis)1)
2.3. Plasma sample collection
Blood samples (approximately 20 mL) were obtained from the jugular vein before morning feeding on the last day of the experiment, and put into tubes containing sodium heparin.Then, the samples were centrifuged at 1 000×g for 20 min(4°C). The supernatant was stored at –20°C prior to analysis.Plasma samples were analyzed for biochemical parameters including plasma urea nitrogen (PUN), cholesterol (CHO),glucose (GLU), total protein (TP), and triglyceride (TG)using an Automated Biochemical Analyzer (HITACHI 7020,Tokyo, Japan). Plasma samples were also analyzed for hormones including leptin (LP), testosterone (T), thyroxin(T3), growth hormone (GH), and insulin (INS) using enzymelinked immune-sorbent assay (ELISA) Kit (Jiancheng Bioengineering Institute, Nanjing, China).
2.4. Ruminal sampling
Ruminal samples were collected from the rumen 2 h after morning feeding on the last day of each finishing phase. The samples were collected using a stomach tube. The tube was fed gently but firmly over the base of the tongue and passed into the esophagus until it reached the rumen. A stirrup pump was used to withdraw a sample. Samples were filtered through four layers of cheesecloth. Immediately after samples were obtained, ruminal pH was measured with a pH meter(PHS-3C, Leici Scientific Instrument Co., Ltd., Shanghai,China). Then the filtrate was centrifuged at 4 000 r min–1for 15 min. Approximately 10 mL of the supernatant liquid was acidified with 3 mL of 25% HPO3(H2O:HPO3=4:1) and frozen at –20°C until analysis of NH3-N using a spectrophotometer(UV-1700, Shimazu, Kyoto, Japan) following the method of Broderick and Kang (1980). The supernatant was also analyzed for volatile fatty acid (VFA) concentration using a gas chromatography (GC-2014, Shimazu, Kyoto, Japan)according to the method of Kimet al. (2013).
2.5. Microbial population determination in rectal feces
Fecal score evaluationDefecation was continuously observed, starting approximately 60 min after morning feeding throughout the experimental period. Fresh feces were evaluated and given scores based on the fecal score criteria as described by Ireland-Perry and Stallings (1993).Pictures of the different fecal scores based on these criteria are shown in Fig. 1.
At the end of the experimental period, three animals in each group were selected for collection of fecal samples.Each sample (>100 g) was collected aseptically, about 10 cm into the rectum, and immediately preserved at –40°C (Gowet al. 2008; Maoet al. 2012; Mealeet al. 2016). On the same day (less than 6 h after sampling), samples were sent to the laboratory to quantify and identify species in the microbial population. Each sample was divided into two parts (each part was >50 g); one part was used for main microbial quantity determination using the dilution inocula method, and the other samples were used for microbialflora analysis by the polymerase chain reaction (PCR) and denatured gradient gel electrophoresis(DGGE) technique.E. coli, Salmonella, and Lactobacillus quantity determinationFecal samples were weighed and diluted with sterile saline solution (1:9, w/w); 10 further dilutions were made to 10–2, 10–3, 10–4, 10–5, 10–6, 10–7, and 10–8gradients. Appropriate dilution rates were selected, and 0.1 mL liquid was inoculated in selective medium:E. coliin EMB medium,Salmonellain SS medium, andLactobacillusin MRS medium (Luqiao Technology Co., Ltd., Beijing, China).Each gradient was inoculated three times, and grown at 37°C in a constant temperature incubator, for 24 h forE.coliandSalmonella, and 72 h forLactobacillus. After that, the bacterial colonies were calculated and expressed as lgCFU g–1(lg(Colony-forming units per gram of rectum content)).Microbial diversity analysis in the rectal feces(i) Total DNA extraction of contents in the rectal feces. Bacterial genomic DNA in feces was extracted, using a Fecal DNA Purification Kit (Simgen Biotechnology Co., Ltd., Hangzhou,China). Samples were thawed before extraction, and 0.3–0.4 g sample was poured into 2-mL centrifuge tubes.The procedure was followed according to the manufacturer’s instructions. Extracted DNA was detected using 0.8%agarose gel electrophoresis, and then preserved at –20°C before being used as template DNA for PCR.
(ii) PCR system. PCR primers GC-338F (5´-CG CCCGGGGCGCGCCCCGGGGCGGGGCGGGGGCG CGGGGGCCTACGGGAGGCAGCAG-3´) and 518R(5´-ATTACCGCGGCTGCTGG-3´) were combined to amplify variable 16S rDNA regions in the bacterial species.
PCR reactions of 50 μL contained: 5 μL 10× PCR buffer,3.2 μL dNTP (2.5 mmol L–1), 0.4 μLrTaq(5 U μL–1), 1 μL GC-338F (20 μmol L–1), 1 μL 518R (20 μmol L–1), 2 μL template DNA (50 ng), and sterile double evaporated water was added to 50 μL.
The PCR reaction conditions were as follows: first an incubation step of 5 min at 94°C was run to pre-denature template DNA; subsequently, 30 circles were executed(each cycle consisted of denaturation for 1 min at 94°C,renaturation for 45 s at 55°C, and extension for 1 min at 72°C); finally the amplification products were subjected to a final extension step of 10 min at 72°C. After the reaction,electrophoresis using 1.0% (w/v) agarose gels was used to analyze the amplification products.
Denatured gradient gel electrophoresis (DGGE)PCR samples were loaded directly into 8% (w/v) polyacrylamide gels (denaturant is 7 mol L–1urea and 40% (v/v)propionamide) in 1× TAE with 35–55% denature gradients.Electrophoresis was performed at a constant voltage of 150 V and a temperature of 60°C for 5 h. After electrophoresis,the gels were stained with silver nitrate, and were then colorfixed and scanned.
DGGE image processing and data analysisBacterial diversity indexes were analyzed according to band numbers and the intensity (grayscale) of each stripe in the sample electrophoresis. DGGE Gel Quantity One 4.31 (Bio-Rad Laboratories Inc., California, USA) was used to digitally analyze electrophoresis band numbers and the density of each band in the DGGE profiles. The Shannon-Wiener index (H),richness index (S) and evenness index (E), among other indicators were used to compare the diversity in different samples. The algorithm is shown below:

Fig. 1 Fecal scores 1.0–4.0. This fecal score criteria were derived from Ireland-Perry and Stallings (1993). Scores and visual observation of feces: score 1.0, liquid consistency,splatters on impact, spreads readily, amorphous shape; score 2.0, may pile slightly and spreads and splatters moderately on impact and setting; score 3.0, firm but not hard, piles but spreads slightly on impact and setting; score 4.0, hard, dry appearance, original form not distorted on impact and setting.1.0<score<3.0 was considered slight diarrhea; score<1.0 was considered diarrhea.

Where,Piis the ratio of single band intensity in the sample to the total intensity of all bands in the samples;Nis the richness of all bands from a single lane in the DGGE profiles;Niis the richness of the numberiband; andSis the sum of all band numbers in the sample.
2.6. Statistical analysis
All data were analyzed as a 2×2 factorial arrangement of treatments, using the General Liner Model (GLM) multivariate procedure (SPSS v.15, SPSS, Chicago, IL, USA). The linear model used for each dependent variable accounted for the effects of F:C, FL, and F:C×FL interaction fixed effects. Individual animal was the experimental unit in this study. An extremely significant difference was declared atP<0.01, significant difference was declared atP<0.05, andP<0.10 was considered a tendency.DGGE Gel Quantity One 4.31 (Bio-Rad Laboratories Inc.,California, USA) analysis software package was used to map the number of bands and analyze similarity.
3. Results
3.1. Nutrient intake
Dry matter intake (DMI) and OM intake increased with increasing F:C (P<0.01) and FL (P<0.05) (Table 2). CP intake was not affected by FL, F:C and their interaction (P>0.05). Increasing the F:C enhanced NDF and ADF intakes (P<0.01), but reducing dietary FL did not have a significant effect on NDF and ADF intakes (P>0.05).
3.2. Apparent total tract nutrient digestibilities
The NDF digestibility was affected by FL (P<0.01), and interaction between F:C and FL (P<0.05, Table 2). Under both low F:C and high F:C, NDF digestibility was increased as the FL increased;however, compared with low F:C, high F:C increased NDF digestibility in the SL group (P<0.05) but did not affect it in the WL group (P>0.05). All other nutrient digestibilities were not affected by FL and F:C (P>0.05).
3.3. Plasma biochemical parameters

?
Increasing the F:C enhanced CHO (P<0.01) and LP (P<0.05)levels in the plasma. Decreasing the FL enhanced the plasma TG (P<0.01) and INS levels (P<0.05) (Table 3). GH concentration was affected by both FL and F:C, it increased as the dietary F:C increased and as the FL decreased in calves (P<0.01). Plasma GLU, PUN, TP, T, and T3 concentrations were not affected by FL,F:C and their interaction (P>0.05).
3.4. Ruminal fermentation
The results suggested that the ruminal pH of calves fed with low F:C diets were significantly lower than those in the high F:C treatment (P<0.05) (Table 4). Ruminal NH3-N concentration was not affected by FL and F:C (P>0.05). Increasing the F:C increased ruminal acetic acid (P<0.05) and acetic acid/propionic acid (P<0.01). Butyrate concentration was affected by both FL and F:C, it increased as the dietary F:C decreased and as the FL increased in calves (P<0.01) but was not affected by interaction between FL and F:C (P>0.05). Furthermore,iso-valeric acid concentration was significantly increased with increasing dietary FL (P<0.05). Other ruminal VFAs and total VFA (TVFA) were not affected by FL, F:C and their interaction (P>0.05).
3.5. Fecal scores, and E. coli, Salmonella, and Lactobacillus in rectal feces
Effects of dietary FL and F:C on microorganism quantities in the rectum of calves are shown in Table 5. Results showed that fecal score in the SL group was significantly lower than in the WL group (2.79vs. 3.21,P<0.05), it indicated that slightly more diarrhea occurred in calves offered chopped hay diets. However,no significant effect of F:C on the fecal score was observed in the present study (P>0.05).E. coliin rectal feces was significantly reduced as FL and F:C increased (P<0.05 andP<0.01, respectively).Compared with low dietary FL and F:C groups,Salmonellain rectal feces were significantly reduced in high FL and F:C diet treatments(P<0.05). TheLactobacillusamount in high FL and F:C groups were significantly higher than the low FL and F:C groups (P<0.05).Furthermore, no significant differences were observed for the interaction between FL and F:C on the fecal score, or on the content ofE.coli,Salmonella, orLactobacillus(P>0.05).
3.6. Microbial diversity in the rectal feces
The effects of F:C and FL on the DGGE band numbers and diversity indexes (H, S, E) of microflora in the rectum are shown in Table 5 and Fig. 3. Bands were affected by the interaction between F:C and FL (P<0.05). Under high F:C, band numbers from the WL group was higher than that from the SL group (P<0.05), whereas there was no difference between the WL and SL groups under low F:C (P>0.05). The Shannon-Wiener value had a tendency to increase as FL increased (P=0.06). Richness (S) was significantly affected by the interaction between F:C and FL (P<0.05), and showed the same results with band numbers.

?

?

?
The similarity index of DGGE profiles of microflora in rectum contents between each treatment is shown in Fig. 4. The DGGE similarity profile could be divided into two clusters: SL with low F:C treatment and all other treatments. Microflora similarity in different treatments was 50–73.5%.
4. Discussion
4.1. Nutrient intake

Fig. 2 PCR amplification of 16S rDNA (V3) of bacteria in rectum of calves (M, DL 2000 marker). Primers GC-338F and 518R were used to amplify the 16S rDNA sequence, leading to DNA fragments of about 200 bp. Total microbial DNA concentration in rectum contents is high, with good purity, and the OD260/OD280 value is between 1.8 to 2.0. PCR amplification of the DNA fragments was successful, and PCR products could be used for subsequent denatured gradient gel electrophoresis (DGGE) analysis.
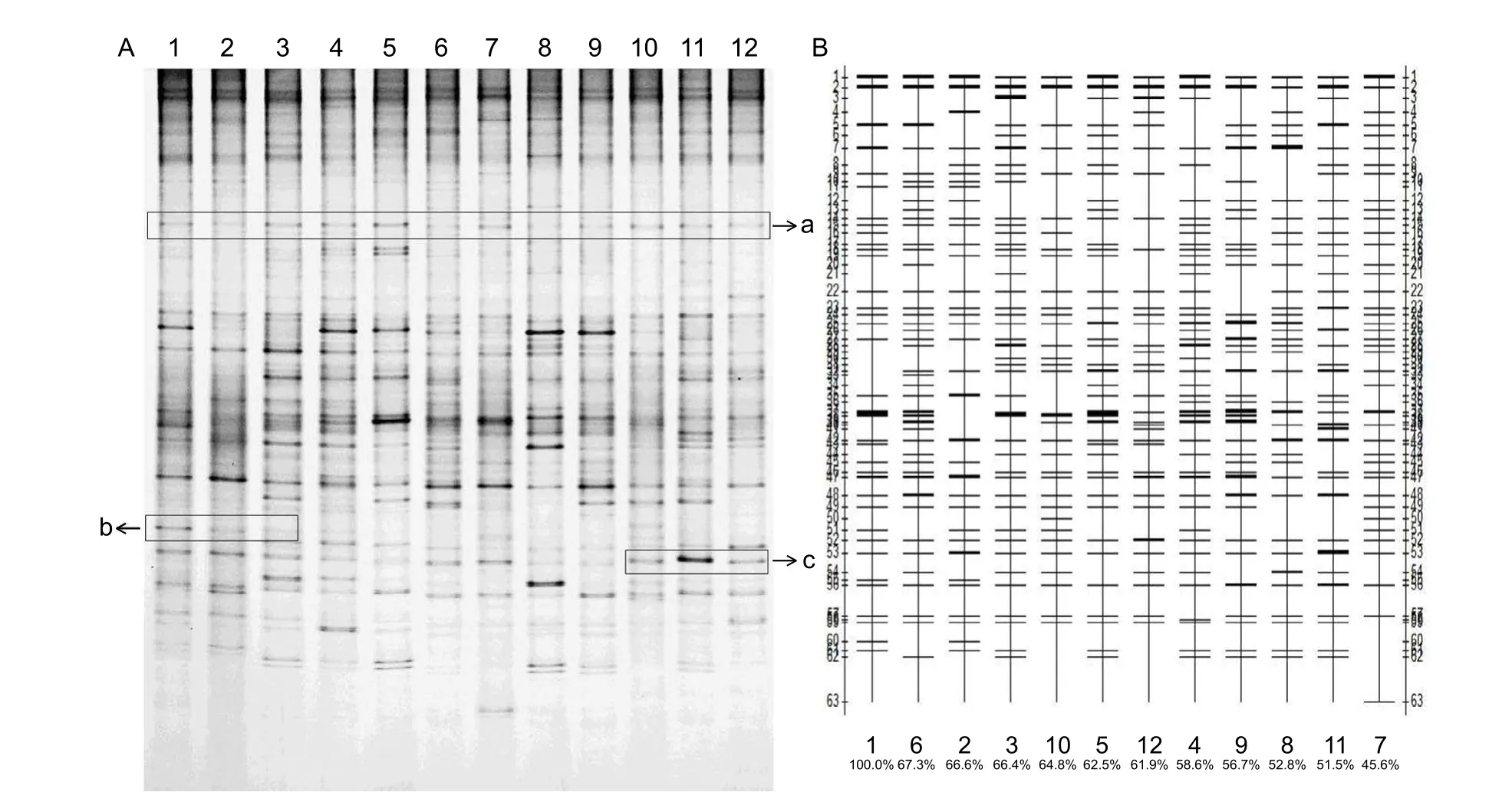
Fig. 3 Photograph of denatured gradient gel electrophoresis (DGGE) bands and the quantitative analysis photograph. A, denatured gradient gel electrophoresis (DGGE) bands. lanes 1, 2, and 3 belong to whole-length with low F:C (low forage to concentrate ratio, 50:50) treatment; lanes 4, 5, and 6 belong to whole-length with high F:C (high forage to concentrate ratio, 65:35) treatment;lanes 7, 8, and 9 belong to short-length with low F:C (50:50) treatment; and lanes 10, 11, and 12 belong to short-length with high F:C (65:35) treatment. a, the common bands; b, the special bands in some treatments; c, the intensity of bands is different among individuals from the same treatment. B, the quantitative analysis version of the DGGE photograph. The number on the horizontal axis of Fig. 3-B means the similarity index of each DGGE lane compared to lane 1, which was defined as the control group.
Forage proportion and its physical characteristics in ruminant diets could affect ruminal buffering, digesta turnover, feed intake, and nutrient utilization (Zebeliet al.2006). A previous study (Sterket al. 2011) reported that linear DMI increased as the dietary concentrate increased.In the current study, we found that DMI and OM intake increased for calves fed diets with increasing forage content.Animals eat feedstuff mainly to satisfy their physiological requirement for energy (Van Soestet al. 1984). Therefore,DMI and OM intake would be inhibited by feeding cattle with low F:C diets, as higher energy density in the concentrate can suppress voluntary forage intake. Krause and Combs(2003) indicated that DMI and OM intake decreased with increasing fermentable carbohydrate in the diet, which was similar to the current study. As described in the current study and in an accompanying study (Muhammadet al. 2016),reduction of FL resulted in a decrease in DMI and OM intake.Consistent with our results, Krause and Combs (2003) also observed a reduction in DMI and OM intake when the forage particle length was decreased. This reduction could be explained by a greater forage length favorably stimulating mastication, rumination activity, and salivary secretions,resulting in an improved rumen condition with preferable buffering capacity, which contributed to the increased feed intake (Zebeliet al. 2006; Castellset al. 2012; Montoroet al.2013). Furthermore, as is shown in our rumen fermentation results, rumen pH values were significantly decreased in the low F:C diet and numerically decreased in the SL diet, which might negatively affect the DMI and OM intake. A study conducted by Stone (2004) demonstrated that the low rumen pH in cattle fed the short forage length diet reduced feed intake. It is possible that the animals were mainly sorting against the long hay particles and for the concentrate, which made up the majority of the short and fine particle fraction and had higher CP content. Sorting for higher CP content might be the reason for the similar intake of all treatments.Animals easily discriminated between long particles and short particles when fed a diet with low F:C (Muhammadet al. 2016). However, regardless of low NDF and ADF concentrations in the low forage diet, increased sorting by calves in the low F:C diet treatment was another important reason for decreased NDF and ADF intakes. Decreased intake of NDF for low F:C diet treatment was also reported by Yanget al. (2001) and Yang and Beauchemin (2007).Another reason for this lower NDF and ADF intakes might be lower organic matter intake for the low F:C diet.
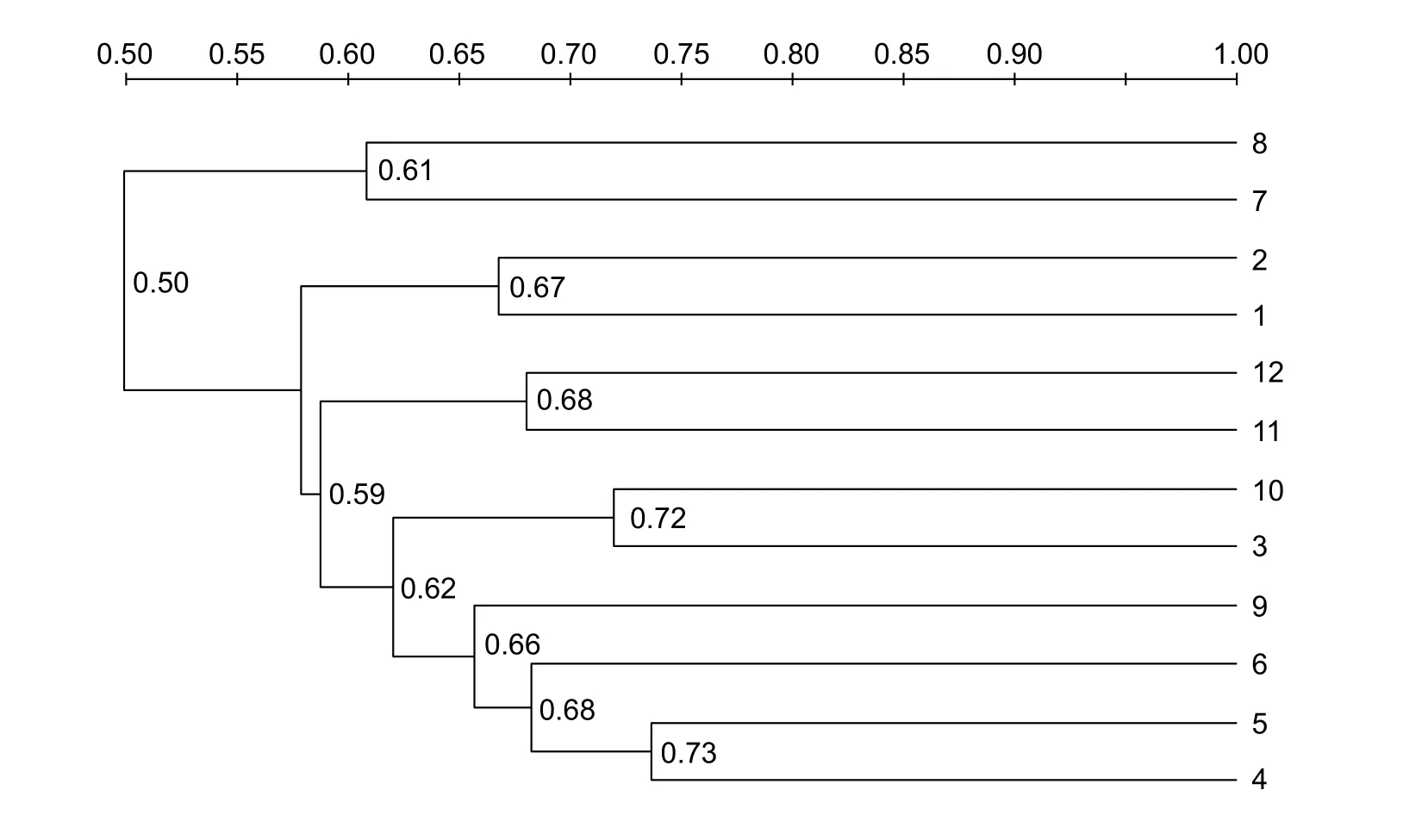
Fig. 4 Similarity index of denatured gradient gel electrophoresis (DGGE) profiles of rectal microflora. Lanes 1, 2, and 3 belong to whole-length with low F:C (low forage to concentrate ratio, 50:50) treatment; lanes 4, 5, and 6 belong to whole-length with high F:C (high forage to concentrate ratio, 65:35) treatment; lanes 7, 8, and 9 belong to short-length with low F:C (50:50) treatment;lanes 10, 11, and 12 belong to short-length with high F:C (65:35) treatments.
4.2. Apparent total tract digestibility of nutrients
In the present study, despite the fact that calves receiving the WL diet consumed similar NDF compared with the SL diet treatment, the apparent NDF digestibility in calves fed the WL diet was higher than those given the SL diet.Kennedyet al. (1984) found that nutrient digestibilities of ruminant animals were determined by the comprehensive impact of the competitive relationship between passage rate and digestion. Increasing the length of roughage could improve animals’ chewing activity but prolong their chewing time (Zebeliet al. 2007), which would increase saliva secretions and enhance rumen-buffering capacity.Therefore, a suitable rumen environment is beneficial forfiber degradation. We speculate that the existence of WL forage in the diet is associated with improving the rumen environment, which would eventually cause positive variation in nutrient digestibility. Nevertheless, it is also possible that long forage in the diet increased digesta retention time in the digestive tract, therefore improving apparent digestibility. Similar results were reported by Yang and Beauchemin (2005) and Al-Saiadyet al. (2010).
4.3. Plasma biochemical parameters
Decreasing the FL enhanced the plasma TG and INS levels in the present study, which is similar to the findings observed by Norouzian and Valizadeh (2014). Delavaudet al. (2002)reported that the plasma LP concentration is higher when sheep and cows have higher DMI. We also found lower DM intake in the low F:C diet treatment, which may be the reason for lower LP concentration (Muhammadet al. 2016).Interestingly, in the present study, the GH level in plasma increased as dietary F:C increased and FL decreased, but there are very few studies on this issue. Moreover, the mechanism by which F:C and FL influence GH is unclear.We suspect that degradation products in the digestive tract could affect plasma parameters such as INS (which has an antagonistic role on GH), which could further influence GH secretion glands.
4.4. Ruminal fermentation
The present study showed that the ruminal pH values of calves fed with low F:C diets were significantly lower than those in the high F:C treatment. Inadequate dietary fiber or lack of fiber effectiveness can reduce chewing time of ruminants, which may lead to reduced saliva secretions and rumen pH (Erdman 1988). Llamas-Lamas and Combs (1991) also observed that ruminal pH in dairy cattle significantly increased when the alfalfa hay proportion increased from 56 to 86% in the diet. This might be because high forage diet content means lower amounts of easily fermentable carbohydrate, less VFA produced, and higher ruminal pH.
VFA is the main fermentation product in the rumen, and could provide approximately 70–80% of the energy needed by ruminants (Bergman 1975). There are many studies regarding the influence of dietary FL and F:C on rumen fermentation in ruminants. Krauseet al. (2002) observed that the TVFA concentration in the rumen fluid increased,whereas the ruminal acetic acid/propionic acid ratio was increased as diet particle size decreased. However, Pittet al. (1996) demonstrated that the TVFA concentration of the rumen fluid was not affected by dietary physically effective NDF (peNDF) levels, while Beaucheminet al.(2003) found that TVFA and the acetate:propionate ratio were not affected by particle size in the diet; these studies were consistent with our results. Different forage to concentrate ratios in ruminant diets could affect energy and non-structural carbohydrate intake, further influencing rumen fermentation (Sutton 1989). Giger-Reverdinet al.(2014) reported that ruminal propionate, butyrate, valerate,and TVFA concentrations in dairy goats were increased by increasing readily-fermentable carbohydrate in the diet.In the present study, we found that higher concentrate treatments decreased the proportion of acetic acid A,possible reason for this is that NDF intake used for acetic acid fermentation was lower in the low F:C treatment.
4.5. Fecal scores, E. coli, Salmonella, and Lactobacillus in the rectum
Stable flora in the gastrointestinal tract plays a very important role in nutrient digestion, absorption, and utilization in animals (Chadwicket al. 1992; Singhet al. 2014). The diet is the source of metabolic substrates for microbes living in the digestive tract, and nutritional changes can significantly affect microbial composition, in terms of the amount and type of flora (Reid and Hillman 1999). Intestinal microflora of young animals is unstable, and feed and environmental changes could cause microbial flora imbalance, ultimately increasing pathogenic microorganisms (Ofeket al. 1975).Present results showed that fecalLactobacilluscontent was significantly higher, whileE.coliandSalmonellacontents were significantly lower in the high FL and F:C group compared with other groups. A possible reason is that insoluble non-starch polysaccharide content is higher in the high F:C diet treatment. Rapid-fermentable carbohydrate could be completely fermented in the front of the digestive tract, and thus have little influence on microflora in the cecum and colon, however, low-speed fermentable carbohydrate could promote the proliferation of beneficial bacteria, thus inhibiting pathogenic microbes (Snelet al.2002). Williamset al. (2001) revealed that dietary nonstarch polysaccharides promote carbohydrate fermentation,thus enhancing the microbial activity in the back end of the intestinal tract. Canibeet al. (2001) reported that acidic conditions (lower pH in the gastrointestinal tract) could inhibit the proliferation of pathogenic bacteria, and be conducive to increasing the population ofLactobacillus. In addition, the high FL diet had a longer retention time for fermentation in the digestive tract than the SL diet, which could ultimately decrease the pH in the digestive tract. This result indicated that nutrient intake and the intestinal environment have large impacts on the composition of microbial flora (Duncanet al. 2009; Xuet al. 2012). Furthermore, Thomlinson and Lawrence (1981) found that the fibrous component in the diet could inhibit the proliferation ofE.coli, effectively improving intestinal health.
Calves in the SL diet treatments had lower fecal scores(higher diarrhea rate) in this study. A possible reason is the increased passage rates of digesta through the digestive tract in the SL wildrye diet treatment (Tafajet al. 2001). This diet may cause a large amount of unfermented carbohydrates to flow into the back end of the digestive tract, which may increase the osmotic pressure and cause excessive water secretion, finally causing diarrhea (Radecki and Yokoyama 1991). Furthermore, unstable microflora and pathogenic bacteria in the gastrointestinal tract could produce toxins that reduce digestion and absorption ability, leading to diarrhea(Williamset al. 2001), which is consistent with the results of our study. Lower fecal scores of calves in chopped hay diet treatments were accompanied by higherE.coliand lowerLactobacilluscontent in rectal feces. This result suggests a correlation among forage length, microflora, and fecal score(diarrhea rate). Gutzwiller and Jost (1998) reported that diarrhea could change native microflora, while total bacteria,LactobacillusandBifidobacteriumquantity in fecal samples were reduced as theE.colinumber increased. Therefore,to a certain extent, feeding the calves with whole-length forage diets could improve fecal scores, and reduce the diarrhea rate.
4.6. Microbial diversity in the rectum
Diet composition affects microflora composition, vitality,and the composition and quantity of fermentable material(Conway 1994). In this study, there were differences in band numbers and intensity based on DGGE, which could reflect the diversity of microbial flora in fecal samples among dietary treatments. Electrophoretic bands showed that fecal microorganism content of calves in WL with high F:C treatment was significantly higher than other groups. The Shannon-Wiener index is usually used to evaluate microflora diversity, and could reflect the microbial population number and its distribution evenness among individuals (Greenberg 1956). Decreasing the FL had a tendency to reduce Shannon-Wiener and richness in the present study (P=0.06).The reduced Shannon-Wiener and richness indexes in SL with high F:C and SL with low F:C treatments might be due to the lower retention time of digesta in the digestive tract. This inference was supported by a higher digestion rate of NDF in the WL hay diet treatment, which might be attributed to higher digesta retention time. With higher digesta retention time, microorganisms have more time to adhere to the indigested feed for proliferation, ultimately increasing the amount and type of fecal microbes. A higher Shannon-Wiener index indicates better dynamic balance among microorganisms, which increases the ability to cope with adverse factors occurring in animals (Hooper and Macpherson 2010; Shanahan 2010). DGGE similarity can clearly reflect the similarities among flora in different samples(Muyzer and Smalla 1998). We acknowledge that individual microflora differences existed within the same treatment,which is consistent with Simpsonet al. (2000). However, our results showed that similarity reached the maximum of 73%intra-treatment but averaged about 55% among different treatments. These results suggested higher microflora similarity among calves in the same treatment than those in different treatments, which indicated the dietary effects on the fecal microflora of calves.
5. Conclusion
Our results demonstrated that dietary F:C and FL must be given sufficient consideration when considering appropriate diets for calves. Dairy calves fed the WL diet had improved NDF digestibility compared to other groups. However,increased ruminal fermentation was observed in calves given diets with an increased F:C ratio. In addition, our results indicated that the rectum microbial composition could be regulated and optimized by adjusting diets, because as theLactobacilluscontent increased, pathogenic microbes decreased in the WL and high F:C feeding groups. Elevating forage length might be an effective feeding practice for lowering the diarrhea rate, an indication of gut health.Overall, we recommend the whole-length wildrye with high F:C diet for weaned calves to optimize the growth and health of male dairy calves.
Acknowledgements
This research was supported by the earmarked fund for China Agriculture Research System (CARS-37) and Special Fund for Agro-scientific Research in the Public Interest(201303144).
Al-Saiady M Y, Abouheif M A, Makkawi A A, Ibrahim H A,Al-Owaimer A N. 2010. Impact of particle length of alfalfa hay in the diet of growing lambs on performance, digestion and carcass characteristics.Asian-Australasian Journal of Animal Sciences, 23, 475–482.
AOAC (Association of Official Analytical Chemistry). 2000.Official Methods of Analysis. 17th ed. Association of Official Analytical Chemistry, Arlington.
Beauchemin K A, Yang W Z, Rode L M. 2003. Effects of particle size of alfalfa based dairy cow diets on chewing activity,ruminal fermentation, and milk production.Journal of Dairy Science, 86, 630–643.
Bergman E N. 1975. Production and utilization of metabolites by the alimentary tract as measured in portal and hepatic blood. In: McDonald I W, Warner A C I, eds.,Digestion and Metabolism in the Ruminant. University of New England Publishing Unit, Armidale, Australia. pp. 292–305.
Broderick G A, Kang J H. 1980. Automated simultaneous determination of ammonia and total amino acids in ruminalfluid andin vitromedia.Journal of Dairy Science, 63, 64–75.
Canibe N, Steien S H, Øverland M, Jensen B B. 2001. Effect of K-diformate in starter diets on acidity, microbiota, and the amount of organic acids in the digestive tract of piglets,and on gastric alterations.Journal of Animal Science, 79,2123–2133.
Castells L I, Bach A, Araujo G, Montoro C, Terré M. 2012. Effect of different forage sources on performance and feeding behavior of Holstein calves.Journal of Dairy Science, 95,286–293.
Chadwick R W, George S E, Claxton L D. 1992. Role of the gastrointestinal mucosa and microflora in the bioactivation of dietary and environmental mutagens or carcinogens.Drug Metabolism Reviews, 24, 425–492.
Conway P L. 1994. Function and regulation of the gastrointestinal microbiota of the pig.European Association for Animal Production Publication, 2, 231–240.
Delavaud C, Ferlay A, Faulconnier Y, Bocquier F, Kann G,Chilliard Y. 2002. Plasma leptin concentration in adult cattle:Effects of breed, adiposity, feeding level, and meal intake.Journal of Animal Science, 80, 1317–1328.
Duncan S H, Louis P, Thomson J M, Flint H J. 2009. The role of pH in determining the species composition of the human colonic microbiota.Environmental Microbiology,11, 2112–2122.
Erdman R A. 1988. Dietary buffering requirements of the lactating dairy cow: A review.Journal of Dairy Science,71, 3246–3266.
Giger-Reverdin S, Rigalma K, Desnoyers M, Sauvant D,Duvaux-Ponter C. 2014. Effect of concentrate level on feeding behavior and rumen and blood parameters in dairy goats: relationships between behavioral and physiological parameters and effect of between-animal variability.Journal of Dairy Science, 97, 4367–4378.
Gow S P, Waldner C L, Harel J, Boerlin P. 2008. Associations between antimicrobial resistance genes in fecal genericEscherichia coliisolates from cow-calf herds in western Canada.Applied and Environmental Microbiology, 74,3658–3666.
Granja-Salcedo Y T, Ribeiro Júnior C S, De Jesus R B, Gomez-Insuasti A S, Rivera A R, Messana J D, Canesin R C,Berchielli T T. 2016. Effect of different levels of concentrate on ruminal microorganisms and rumen fermentation in Nellore steers.Archives of Animal Nutrition, 70, 17–32.
Greenberg J H. 1956. The measurement of linguistic diversity.Language, 32, 109–115.
Gutzwiller A, Jost M. 1998. Piglet diarrhea and oedema disease:Prevention is better.Agrarforschung Schweiz, 5, 459–462.
Hintz R W, Mertens D R, Albrecht K A. 1996. Effects of sodium sulfite on recovery and composition of detergent fiber and lignin.Journal of Association of Official Analytical Chemistry,79, 16–22.
Hooper L V, Macpherson A J. 2010. Immune adaptations that maintain homeostasis with the intestinal microbiota.Nature Reviews Immunology, 10, 159–169.
Ireland-Perry R L, Stallings C C. 1993. Fecal consistency as related to dietary composition in lactating Holstein cows.Journal of Dairy Science, 76, 1074–1082.
Kennedy P M, Hazlewood G P, Milligan L P. 1984. A comparison of methods for the estimation of the proportion of microbial nitrogen in duodenal digesta, and of correction for microbial contamination in nylon bags incubated in the rumen of sheep.British Journal of Nutrition, 52, 403–417.
Kim E T, Min K S, Kim C H, Moon Y H, Kim S C, Lee S S. 2013.The effect of plant extracts on in-vitro ruminal fermentation,methanogenesis and methane-related microbes in the rumen.Asian-Australasian Journal of Animal Sciences,26, 517–522.
Kononoff P J, Heinrichs A J. 2003. The effect of corn silage particle size and cottonseed hulls on cows in early lactation.Journal of Dairy Science, 86, 2438–2451.
Krause K M, Combs D K. 2003. Effects of forage particle size,forage source, and grain fermentability on performance and ruminal pH in midlactation cows.Journal of Dairy Science,86, 1382–1397.
Krause K M, Combs D K, Beauchemin K A. 2002. Effects of forage particle size and grain fermentability in midlactation cows. II. Ruminal pH and chewing activity.Journal of Dairy Science, 85, 1947–1957.
Llamas-Lamas G, Combs D K. 1991. Effect of forage to concentrate ratio and intake level on utilization of early vegetative alfalfa silage by dairy cows.Journal of Dairy Science, 74, 526–536.
MAFF (Ministry of Agriculture, Fisheries and Food, United Kingdom). 1984.Energy Allowances and Feeding Systems for Ruminants.Reference Book 433. Her Majesty’s Stationery Office, London. p. 85.
Mao S, Zhang R, Wang D, Zhu W. 2012. The diversity of the fecal bacterial community and its relationship with the concentration of volatile fatty acids in the feces during subacute rumen acidosis in dairy cows.BMC Veterinary Research, 8, 237.
Meale S J, Li S C, Paula A, Hooman D, Plaizier J C, Ehsan K, Steele M A. 2016. Development of ruminal and fecal microbiomes are affected by weaning but not weaning strategy in dairy calves.Frontiers in Microbiology, 7, 582.Montoro C, Miller-Cushon E K, DeVries T J, Bach A. 2013.Effect of physical form of forage on performance, feeding behavior, and digestibility of Holstein calves.Journal of Dairy Science, 96, 1117–1124.
Mooney C S, Allen M S. 1997. Physical effectiveness of the neutral detergent fiber of whole linted cottonseed relative to that of alfalfa silage at two lengths of cut.Journal of Dairy Science, 80, 2052–2061.
Muhammad A U R, Xia C Q, Cao B H. 2016. Dietary forage concentration and particle size affect sorting, feeding behaviour, intake and growth of Chinese Holstein male calves.Journal of Animal Physiology and Animal Nutrition,100, 217–223.
Muyzer G, Smalla K. 1998. Application of denaturing gradient gel electrophoresis (DGGE) and temperature gradient gel electrophoresis (TGGE) in microbial ecology.Antonie van Leeuwenhoek, 73, 127–141.
Norouzian M A, Valizadeh R. 2014. Effect of forage inclusion and particle size in diets of neonatal lambs on performance and rumen development.Journal of Animal Physiology and Animal Nutrition, 98, 1095–1101.
Ofek I, Beachey E H, Jefferson W, Campbell G L. 1975. Cell membrane-binding properties of group A streptococcal lipoteichoic acid.Journal of Experimental Medicine, 141,990–1003.
Pitt R E, Van Kessel J S, Fox D G, Pell A N, Berry M C, Van Soest P J. 1996. Prediction of ruminal volatile fatty acids and pH within the net carbohydrate and protein system.Journal of Animal Science, 74, 226–244.
Radecki S V, Yokoyama M T. 1991. Intestinal bacteria and their influence on swine nutrition. In: Miller E R, Ullrey D E,Lewis A J, eds.,Swine Nutrition. Butterworth-Heinemann,Stoneham, MA, USA. pp. 439–447.
Reid C A, Hillman K. 1999. The effects of retrogradation and amylose/amylopectin ratio of starches on carbohydrate fermentation and microbial populations in the porcine colon.Animal Science(Penicuik, Scotland), 68, 503–510.
Shanahan F. 2010. Probiotics in perspective.Gastroenterology,139, 1808–1812.
Simpson J M, McCracken V J, Gaskins H R, Mackie R I. 2000.Denaturing gradient gel electrophoresis analysis of 16S ribosomal DNA amplicons to monitor changes in fecal bacterial populations of weaning pigs after introduction ofLactobacillus reuteristrain MM53.Applied and Environmental Microbiology, 66, 4705–4714.
Singh K M, Shah T M, Reddy B, Deshpande S, Rank D N, Joshi C G. 2014. Taxonomic and gene-centric metagenomics of the fecal microbiome of low and high feed conversion ratio(FCR) broilers.Journal of Applied Genetics, 55, 145–154.
Snel H, Hoolwerf J, Wissink-Lettink M, Bovee-Oudenhoven I, Van Der Meer R, Herrewegh A. 2002. Use of real-time polymerase chain reaction for microflora analysis.British Journal of Nutrition, 88, 117–118.
Van Soest P J, Ferreira A M, Hartley R D. 1984. Chemical properties of fiber in relation to nutritive quality of ammonia treated forages.Animal Feed Science and Technology,10, 155–164.
Van Soest P J, Robertson J B, Lewis B A. 1991. Methods for dietary fiber, neutral detergent fiber, and non-starch polysaccharides in relation to animal nutrition.Journal of Dairy Science, 74, 3583–3597.
Sterk A, Johansson B E, Taweel H Z, Murphy M, van Vuuren A M, Hendriks W H, Dijkstra J. 2011. Effects of forage type, forage to concentrate ratio, and crushed linseed supplementation on milk fatty acid profile in lactating dairy cows.Journal of Dairy Science, 94, 6078–6091.
Stone W C. 2004. Nutritional approaches to minimize subacute ruminal acidosis and laminitis in dairy cattle.Journal of Dairy Science, 87, E13–E26.
Sutton J D. 1989. Altering milk composition by feeding.Journal of Dairy Science, 72, 2801–2814.
Tafaj M, Steingass H, Drochner W. 2001. Influence of hay particle size at different concentrate and feeding levels on digestive processes and feed intake in ruminants. 2.Passage, digestibility and feed intake.Archives of Animal Nutrition, 54, 243–259.
Thomlinson J R, Lawrence T L. 1981. Dietary manipulation of gastric pH in the prophylaxis of enteric disease in weaned pigs: Some field observations.Veterinary Record, 109,120–122.
Williams B A, Verstegen M W A, Tamminga S. 2001.Fermentation in the large intestine of single-stomached animals and its relationship to animal health.Nutrition Research Reviews, 14, 207–227.
Xu X, Xu P, Ma C, Tang J, Zhang X. 2012. Gut microbiota, host health, and polysaccharides.Biotechnology Advances, 31,318–337.
Yang W Z, Beauchemin K A. 2005. Effects of physically effectivefiber on digestion and milk production by dairy cows fed diets based on corn silage.Journal of Dairy Science, 88,1090–1098.
Yang W Z, Beauchemin K A. 2007. Altering physically effectivefiber intake through forage proportion and particle length:Digestion and milk production.Journal of Dairy Science,90, 3410–3421.
Yang W Z, Beauchemin K A, Rode L M. 2001. Effects of grain processing, forage to concentrate ratio, and forage particle size on rumen pH and digestion by dairy cows.Journal of Dairy Science, 84, 2203–2216.
Zebeli Q, Tafaj M, Steingass H, Metzler B, Drochner W. 2006.Effects of physically effective fiber on digestive processes and milk fat content in early lactating dairy cows fed total mixed rations.Journal of Dairy Science, 89, 651–668.
Zebeli Q, Tafaj M, Weber I, Dijkstra J, Steingass H, Drochner W. 2007. Effects of varying dietary forage particle size in two concentrate levels on chewing activity, ruminal mat characteristics, and passage in dairy cows.Journal of Dairy Science, 90, 1929–1942.
杂志排行

Journal of Integrative Agriculture的其它文章
- Rapid mapping of candidate genes for cold tolerance in Oryza rufipogon Griff. by QTL-seq of seedlings
- A dCAPS marker developed from a stress associated protein gene TaSAP7-B governing grain size and plant height in wheat
- A major quantitative trait locus controlling phosphorus utilization efficiency under different phytate-P conditions at vegetative stage in barley
- Overexpression of IbSnRK1 enhances nitrogen uptake and carbon assimilation in transgenic sweetpotato
- Collision detection of virtual plant based on bounding volume hierarchy: A case study on virtual wheat
- lntegrated management strategy for improving the grain yield and nitrogen-use efficiency of winter wheat