Multi-functional roles of TaSSI2 involved in Fusarium head blight and powdery mildew resistance and drought tolerance
2018-02-05HULiqinMUJingjingSUPeisenWUHongyanYUGuanghuiWANGGuipingWANGLiangMAXinLlAnfeiWANGHongweiZHAOLanfeiKONGLingrang
HU Li-qin, MU Jing-jing, SU Pei-sen, WU Hong-yan, YU Guang-hui, WANG Gui-ping, WANG Liang, MA Xin, Ll An-fei, WANG Hong-wei, ZHAO Lan-fei, KONG Ling-rang
1 State Key Laboratory of Crop Biology/Shandong Key Laboratory of Crop Biology/College of Agronomy, Shandong Agricultural University, Tai’an 271018, P.R.China
2 Agriculture Bureau of Rushan, Rushan 264500, P.R.China
3 Shandong Agricultural University Fertilizer Science Technology Co., Ltd., Feicheng 271600, P.R.China
4 College of Resources and Environmental Sciences, China Agricultural University, Beijing 100193, P.R.China
1. lntroduction
Fusarium head blight (FHB) or scab is a devastating disease of wheat (Triticum aestivum) and barley (Hordeum vulgare)in humid and warm regions, which is mainly caused byFusarium graminearumandFusarium avenaceum(Bai and Shaner 1994). The explode of FHB not only reduces yield, but also deteriorates grain quality, due to the grains contaminated with trichothecene mycotoxins mainly as deoxynivalenol (DON) and other trichothecenes produced by pathogen (McMullenet al. 1997). DON is a major virulence factor known as a protein synthesis inhibitor, which can inhibit peptidyl transferase binding to a special site on the 60S ribosomal subunit. Thus, the health of humans and animals could be threaded once ingested DON contaminated grains(Pestkaet al. 2004; Rochaet al. 2005).
Although more than 100 resistance quantitative trait loci(QTLs) for FHB resistance have been identified in different sources (Bollinaet al. 2010), the molecular mechanism to control resistance to the disease in wheat is poorly characterized. Recently, the major resistance gene ofFhb1was successfully cloned as pore-forming toxin-like (PFT) gene encoding a chimeric lectin with two agglutinin domains and an ETX/MTX2 toxin domain, while the underlying resistance mechanism is still unraveled (Rawatet al. 2016). The hemibiotrophic pathogen ofF.graminearumandF.avenaceumare thought to more easily germinate in floweret, especially on anther, exhibiting biotrophic phase during the early stage of infection. Once the hyphae penetrated cuticle layer into the cell, the DON would be secreted to destroy plant cell and the pathogen transfer to the necrotrophic phase (Jansenet al. 2005). In model plants, defense signaling had been well studied and believed to play important roles in disease resistance, with a general consensus that salicylic acid(SA) and jasmonic acid (JA) signalings are antagonistic but confer resistance to biotrophic and necrotrophic pathogen,respectively. However, their roles in FHB resistance is still in debate even dozen studies had been carried out (Makandaret al. 2010, 2012; Sorahinobaret al. 2015; Qiet al. 2016).Several transcriptomic studies using near-isogenic lines(NILs) have revealed the importance of JA in FHB defense(Xiaoet al. 2013). Silencing ofAOSin wheat, a putative marker gene in JA synthesis, leaded to increased susceptibility of FHB, whilein vitrotreatment of MeJA reduced the number of diseased spikelet (Qiet al. 2016). After silencing a JA-dependent signaling geneTaJRLL1, the infection ofF.graminearumwas more serious than the negative control in detached leaves (Xianget al. 2011). A combination of transcriptomic and proteomic study demonstrated that SA,ethylene (ET) and JA might play important roles in the defense ofF.graminearuminfection (Li and Yen 2008; Dinget al. 2011; Xiaoet al. 2013). Furthermore,AtPGIPwhich translated polygalacturonase-inhibiting proteins to restrict pectin degradation during fungal infection could restrict the development of FHB symptoms when over-expressing in wheat (Ferrariet al. 2012). Recently,AtNPR1, a key plant defence regulator inArabidopsisSA pathway, was reported to effectively suppress FHB symptom expanded to flanking spikelet after over-expressing it in wheat (Makandaret al.2006; Gaoet al. 2013). This is relatively amazing as more JA response was found at late stage ofF.graminearuminfection while rare SA responsive genes were observed at the very early stage of infection (Dinget al. 2011). As the disease developmental process of FHB is rather complex,more resistant genes need to be cloned to unravel the resistance mechanism of wheat in detail.
Fatty acids and lipids are also implicated in plant defense signaling pathways (Maldonadoet al. 2002; Kachroo and Kachroo 2009; Okazaki and Saito 2014). For instance, theSSI2(suppressor of SA-insensitivity ofnpr1-5) encoded stearoyl-acyl carrier protein-desaturase (SACPD) was involved in pathogen resistance. A mutation in theArabidopsis SSI2gene resulted in reduced oleic acid (18:1) level, activating SA-mediated defense responses with high levels of SA and constitutive expression of pathogenesis-related (PR)genes (Kachrooet al. 2001). Consequently, these plants exhibit enhanced resistance toPeronospora parasitica,Pseudomonas syringae(Shahet al. 2001), andCucumber mosaicvirus(Sekineet al. 2004). Furthermore, the mutant plants are stunted in size and exhibit spontaneous lesion formation on their leaves. The SACPD catalyzes conversion of stearic acid (18:0) to oleic acid (18:1) in plants(Shanklin and Somerville 1991; Cahoon and Shanklin 1997). TheArabidopsisSACPD family consists of seven highly conserved members, among themSSI2makes great contribution to the total 18:1 pool in the plant (Kachrooet al.2007). An increasing body of evidence has demonstrated the importance of 18:1 in plant defense pathway. Mutations in the glycerol-3-phosphate (G3P)-acyl transferase (ACT1)or the G3P dehydrogenase (GLY1) gene which can restore wild-type levels of 18:1 are able to restore all the altered phenotypes inssi2plants (Kachrooet al. 2003, 2004). Exogenous application of glycerol also lowered the 18:1 content inSSI2plants and converted these plants intossi2-mimics(Kachrooet al. 2004). As to the low 18:1-drived pathway,Mandalet al. (2012) thought oleic acid could modulate levels of nitric oxide-associated protein (NOA1), thereby regulating nitric oxide-mediated defense signaling inArabidopsis.
In recent years, a number ofSACPDgenes have been isolated from crops. In soybean, silencing ofGmSACPDsinduces a variety of defense-related phenotypes and confers resistance to multiple pathogens, such asPseudomonas syringaepv. glycinea (Kachrooet al. 2008). RNAi-mediated knockdown ofOsSSI2(OsSSI2-kd) in rice enhanced resistance to the blast fungusMagnaporthe griseaand leaf blight bacteriaXanthomonas oryzaepv. oryzae (Jianget al. 2009). Over-expression of a wheatSACPDgene inArabidopsisssi2mutant compromise its resistance to powdery mildew (Golovinomyces cichoracearum) (Songet al. 2013).
In this paper, we cloned aTaSSI2gene from wheat. For functional characterization of this gene, barley stripe mo-saic virus (BSMV)-induced gene silencing (VIGS) and the transgenicBrachypodiumwere used to analyze its function involved in powdery mildew, FHB resistance and abiotic stress tolerance. The underlying molecular mechanism was also primarily characterized.
2. Materials and methods
2.1. Plant materials and growth conditions
The FHB-resistant wheat cultivar Sumai 3 was used for gene cloning. The wild-typeNicotiana benthamianaplants were used as mediation of virus-assembly for BSMV-VIGS experiment. Powdery mildew-susceptible cultivar Huixianhong was inoculated with infectedN.benthamianasap and grown in a chamber with the photoperiod of 16/8 h and temperature of 24/20°C (day/night). The wheat of KN199 for FHB resistance evaluation andB.distachyon(Bd21) and transgenicBrachypodiumplants were grown in a greenhouse. The tetraploid wheat mutants of “Kronos” were kindly provided by Prof. Fu Daolin (Shandong Agricultural University, China).
2.2. In silico analysis
2.3. Virus-induced gene silencing of TaSSI2
BSMV-VIGS was performed as described previously (Yuanet al. 2011; Houet al. 2015). Briefly, the fragment ofTaSSI2was obtained by RT-PCR using the primer pair TaSSI2F-lic/TaSSI2R-lic (Appendix A) and then was inserted into the vector of pCa-γbLIC. Each of the pCaBS-α, pCaBS-β and pCa-γbLIC plasmid was independently transformed intoAgrobacterium tumefaciensstrain EHA105 and cultured at 28°C, 200 r min–1in LB medium containing rifampicin(25 μg mL–1) and kanamycin (50 μg mL–1). The obtained bacterial cells were pelleted at 2 200×g for 10 min, resuspended in infiltration buffer (10 mmol L–1MgCl2, 10 mmol L–12-(N-morpholino) ethanesulfonic acid (MES), pH 5.2, and 0.1 mmol L–1acetosyringone) to 0.7 OD600and incubated at room temperature for 3 to 5 h with equal amounts of three different bacteria. Then the mixture was infiltrated into leaves ofN.benthamianawith a 1-mL needleless syringe.After maintenance in a growth chamber for 8 to 12 days,the infiltrated leaves were harvested, ground in 20 mmol L–1Na-phosphate buffer (pH 7.2) containing 1% celite, and the sap was inoculated by hand rubbing method onto the two-leaf stages of wheat. The procedure of VIGS in wheat spikes was similar to the above. The infected wheat leaves were harvested and ground in 20 mmol L–1Na-phosphate buffer (pH 7.2) containing 1% celite. Then, the entire leaves or spikes surface were then coated with this mixture by sliding the gently pinched fingers from base to tip three times.
2.4. Powdery mildew inoculation assays
At 8–12 days after inoculation withN.benthamianaleaf sap,wheat developing visible BSMV symptoms was challenged withB.graminisf. sp. triticum isolate E09, by dusting test plants with conidia from the infected wheat. Plants were grown in a chamber as described above until mildew observations were recorded. For the powdery mildew test, we used a 1–4 scale, with 1 representing necrotic flecks with low sporulation (highly resistant), 2 for necrosis with medium sporulation (resistant), 3 as no necrosis with medium to high sporulation (susceptible) and 4 as no necrosis with full sporulation (highly susceptible) (Liuet al. 2002).
2.5. FHB resistance evaluation of plants
Thein vitro F.graminearuminfection assay on detached wheat leaves was performed as described previously with minor modifications (Chenet al. 2009; Xianget al. 2011;Houet al. 2015). Briefly, 5-cm long leaf sections were cut from the central portion of the third leaves detached from seedlings and wounded on the adaxial surface, and then placed on a 1% water-agar medium. A conidial suspension ofF.graminearum(3 μL of 1×106conidia mL−1) was applied to the fresh wound on the adaxial surface. After inoculation,the plates were sealed and placed in a growth chamber at 20–24°C with a 16/8 h (light/dark) cycle. The saprophytic spot length was measured 4 days post inoculation (DPI)after trypan blue staining.
Type II FHB resistance evaluation was performed as described previously with minor modifications (Shenet al.2004; Zhanget al. 2011). Briefly, single-floret inoculation was conducted for FHB type II disease evaluation after VIGS. At anthesis, the basal florets of the third or fourth spikelet from the tip of virus infected spikes were each inoculated with 10 μL ofF.graminearumspore suspension(1×106conidia mL−1), and the inoculated spikes were covered with transparent plastic bags for 72 h to maintain high humidity. Five days after inoculation, 30 plants of each group were evaluated for FHB response. The extending rate was calculated and recorded as disease severity measured as the ratio of the number of diseased spikelet in all of the inoculated spikes.
2.6. Over-expression of TaSSI2 in B. distachyon and drought tolerance evaluation
To over-expressTaSSI2gene inB.distachyon(Bd21), the coding region ofTaSSI2-2ALwas sub-cloned into the vector of pCambia1300-35s-sGFP, and then transformed into theA. tumefaciensAGL1 (Yuet al. 2014). Tissue culture andAgrobacterium-mediated transformation ofBrachypodiumwas optimized from the protocol described previously(Bragget al. 2012). Transgenic lines were selected on callus induction medium (CIM) plates containing 150 mg L–1timentin and putative transgenic plants were confirmed by PCR analysis. The homozygous T3lines were used for further phenotypic analysis. It has been demonstrated thatBrachypodiumcan be used as a new host to study FHB(Peraldiet al. 2011). For drought tolerance evaluation,25% PEG10000 was employed for treatments as described previously (Qinet al. 2014). The contents of stearic acid(18:0), oleic acid (18:1), linoleic (18:2) and linolenic acid(18:3) were measured.
2.7. Subcellular localization
TaSSI2without stop codon was amplified using specific primer pair S-TaSSI2F/R (Appendix A) and integrated into the pBIN35S:EGFP vector that was then transformed into theA. tumefaciensstrain EHA105 (Qinet al. 2014). The transformed bin vector of 35S:EGFP was used as a control.TheAgrobacteriumculture was resuspended in infiltration medium (10 mmol L–1MES, 10 mmol L–1MgCl2, pH 5.2, and 200 mmol L–1acetosyringone) to 0.7 OD600and injected into 4-week-oldN.benthamianaleaves. For co-localization, theAgrobacteriumharboring TaSSI2:EGFP and theAgrobacteriumharboring G-rbCD3-968 (Golgi apparatus)or pt-rbCD3-1000 (plastids) were mixed with a 1:1 ratio and co-injected in tobacco leaves as described above(Nelsonet al. 2007). After infiltration, plants were kept in an incubator for 2–3 days until used for microscopic observation. In addition, TaSSI2:EGFP and G-rbCD3-968 were transformed into the onion (Allium cepa) epidermis with a Bio-Rad PDS-1000/He Biolistic Particle Delivery System(Bio-Rad, USA) (Daset al. 2009). The fluorescent images were captured by the Zeiss LSM510 Confocal Laser Microscope (Carl Zeiss Vision, Oberkochen, Germany).GFP-mCherry double-labeled plant materials were captured alternately using channel switching with the multitrack function (488 nm for GFP and 543 nm form Cherry) and detected using a 530- to 600-nm band pass filter for GFP and a 575- to 650-nm band pass filter for mCherry.
2.8. RNA extraction, RT-PCR and qPCR
For exogenous hormone treatments, the seedlings were treated with 1.5 mmol L–1SA, 200 μmol L–1methyl jasmonate (MeJA), 50 μmol L–11-amino cyclopropane-1-carboxy(ACC), 50 μmol L–1indole acetic acid (IAA), 50 μmol L–1abscisic acid (ABA), respectively. The seedling leaves were collected respectively at 0, 6, 12, 24 and 48 h post treatment and frozen immediately in liquid nitrogen, and stored at –80°C. Total RNA was extracted using RNA Prep Pure Plant Kit (Tiangen, Germany) according to the manufacturer’s instructions. The cDNA was synthesized using PrimeScriptTMII 1st Strand cDNA Synthesis Kit (TaKaRa,Japan) with oligo(dT) primers according to manufacturer’s protocol. Quantitative real-time PCR analysis was performed using a Roche LightCycler®480 (Roche Diagnostics GmbH, Mannheim, Germany) as described previously (Houet al. 2015). The housekeeping gene β-actin was used as an internal standard (Maet al. 2015). The threshold cycle(CT) values were used to calculate the fold change in transcript accumulation with the formula 2–ΔΔCT. All quantitative real-time (qRT)-PCR reactions were performed on three biological replicates, and three technical replicates were conducted for each sample. All primers used in this study are listed in Appendix A.
根据胡广军关于汽车座椅研究[15]可知:在坐姿状态下,人体的共振频率在4~6Hz。其中,胸、腹部的共振频率在4~8Hz,头、颈部的共振频率在20~30Hz。结合图6可以得出以下结论:
3. Results
3.1. Cloning and phylogenetic analysis of TaSSI2
According to theAtSSI2(At2g43710), the homologous copies in wheat genome were searched (http://plants.ensembl.org/Triticum_aestivum/Info/Index), and a total of three gene copies on A, B and D genomes were found (AA0283540,AA0388780 and AK332689), named asTaSSI2-2AL,TaSSI2-2BLandTaSSI2-2DL, respectively. Based on the reference sequence information ofTaSSI2-2AL, a 1 179-bp sequence was cloned from Sumai3 cDNA using the primer pair TaSSI2F/R (Appendix A) and designated asTaSSI2-2A.The open reading frame (ORF) ofTaSSI2-2Aencoded a 392-amino acids protein with a predicted molecular weight of 44.57 kDa and an isoelectric point of 6.18. In addition toTaSSI2-2A, we also clonedTaSSI2-2BandTaSSI2-2Dfrom Sumai 3. In order to further investigate the relationship betweenTaSSI2and otherSACPDS, we performed a maximum likelihood phylogenetic tree using MEGA 7. As shown in Fig. 1, threeTaSSI2members (AA0283540, AA0388780 and AK332689) and their closest homolog of Os04g037990 were clustered in a same branch with 100% bootstrap value. The putativeOsSSI2sequence published before were clustered with another wheat gene of AA0667810.1, while the putativeAtSSI2sequence fell in a relative far position together with severalArabidopsismembers (Kachrooet al.2003; Jianget al. 2009).
3.2. Functional analysis of TaSSI2 involved in powdery mildew resistance
To further confirm the function ofTaSSI2, we used the BSMV-VIGS system to transitionally silence the gene. A 208-bp conserved fragment of three copies ofTaSSI2were cloned into the vector of pCa-γbLIC for gene silencing. This fragment was blasted against the Chinese Spring genome(http://plants.ensembl.org/Triticum_aestivum/Info/Index)and confirmed the specificity toTaSSI2that no more than 20 bp homology could be found in other positions of wheat genome. Photo-bleaching caused by silencing of phytoene desaturase (PDS) was used as the reporter in BSMVVIGS studies because it produced an obvious phenotype(Yuanet al. 2011). After 12 days of virus infection, the photo-bleaching phenotype was observed in new emerging leaves of BSMV:TaPDS as expected (Fig. 2-A). These results demonstrated that the BSMV-VIGS system in our study could be successfully applied for gene silencing in wheat. By using this system, the expression ofTaSSI2in leaves was verified by qRT-PCR that reduction ofTaSSI2expression by nearly 80% was evident in our experiments at 8 DPI (Fig. 2-B). The potential effects ofTaSSI2silencing on powdery mildew infections were assessed at 10 days after BSMV:00 (empty vector) or BSMV:TaSSI2inoculation of Huixianhong plants. At 8 days after conidia applications,most leaves infected with the BSMV:00 control virus had wilted and displayed extensive mildew colonization and growth, whereas the BSMV:TaSSI2-treated plants were more robust and showed much lower levels of mildew development (Fig. 2-C). For statistical evaluations of the powdery mildew infections, phenotype identification was performed as described above (Liuet al. 2002). BSMV:TaSSI2showed higher ratio of highly resistant (1), resistant (2) plants but lower ratio of highly susceptible (4) plants than BSMV:00(Fig. 2-D). Thus, down-regulation ofTaSSI2resulted in decreased susceptibility to powdery mildew infection, as was the case with the previous study inA.thaliana(Songet al. 2013).
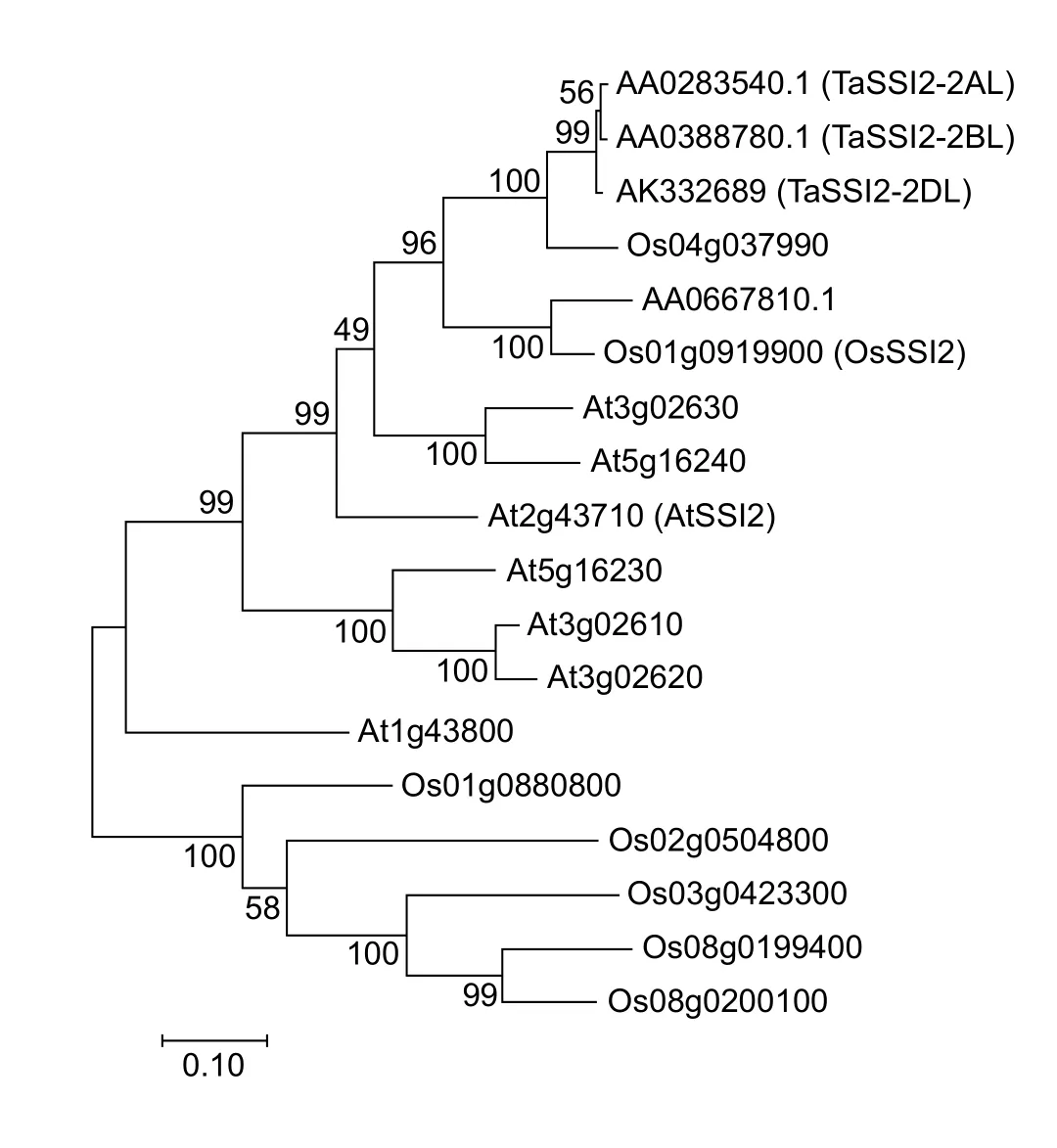
Fig. 1 Phylogenetic relationship of TaSSI2. The TaSSI2 and the putative homologs from Oryza sativa and Arabidopsis were used for constructing phylogenetic tree by using the maximum likelihood method based on the Tamura-Nei model, with 1 000 bootstrap replicates.
3.3. Functional analysis of TaSSI2 involved in FHB resistance
To determine whetherTaSSI2is involved in resistance toF.graminearum, the BSMV-VIGS system was employed on wheat spikes. The recombinant virus harboring bin vector of BSMV:00 and BSMV:TaSSI2were inoculated on flag leaf of wheat cultivar KN199. Before anthesis, the gene silencing in glumes were confirmed and then the basalfloweret of spikes were inoculated byF.graminearum, the disease development was then speculated in the next two weeks (Fig. 3). It was shown that the diseased spikelets with BSMV:TaSSI2were significantly reduced compared with the plants with control vector of BSMV:00, supporting the negatively regulation ofTaSSI2in type II FHB resistance(Fig. 3-A and D). Moreover, the detached leaves from seedlings were also challenged withF.graminearumat 10 DPI with BSMV:00 as the negative control. The pathogen infection on leaves was observed 4 days after inoculation ofF.graminearum. Significantly more and faster hyphae growth was observed in the control than theTaSSI2silenced leaves (Fig. 3-B), which was consistent with the results on spike, further implying that silencing ofTaSSI2enhanced the resistance of plant cell to pathogen infection.
To further investigate the regulation in FHB, the tetraploid wheat tilling mutants ofTaSSI2were employed to evaluate the FHB resistance. A total of 3 mutants were found harboring stop mutations onTaSSI2-2ALand 1 mutant onTaSSI2-2BL(Fig. 3-F, G and H). ForTaSSI2-2AL, the mutant of Kronos 4618 showed a slightly enhanced FHB resistance while Kronos 2655 and Kronos 2143 were apparently more resistant than the wild-type plants (Fig. 3-G). The mutant of Kronos 655 harbors a same mutation with Kronos 2143 on position of 619 bp, which also exhibited enhanced FHB resistance (Fig. 3-G and H). These results suggested thatTaSSI2negatively regulated FHB resistance and this effect might be correlated with gene dosage in tetraploid wheat.
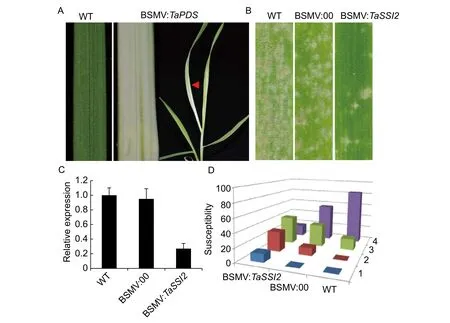
Fig. 2 Powdery mildew resistance of TaSSI2 by barley stripe mosaic virus (BSMV)-induced gene silencing (VIGS). A, symptoms of BSMV-VIGS using PDS gene. B, phenotype of TaSSI2-silenced leaves against powdery mildew strain E09. BSMV:00, BSMV containing empty vectors; BSMV:TaSSI2, BSMV containing TaSSI2. C, verification of silence of TaSSI2 in BSMV-infected leaves by quantitative real-time (qRT)-PCR. The cDNA from 7 days post inoculation (DPI) leaves was amplified by primers amplifying the total amount of TaSSI2 transcripts for A, B and D copies. D, the statistics of powdery mildew resistance of TaSSI2-silenced leaves.The disease symptoms were noted from 1 to 4 scale, representing highly resistant, resistant, susceptible and highly susceptible,respectively. WT, wild type. Data are means±SD.
3.4. Transcriptional regulation of TaSSI2
To obtain a clue on the possible involvement of a regulatory pathway, the transcriptional response ofTaSSI2against various hormones was carried out by qRT-PCR. Due to the extreme sequence similarity of three copies ofTaSSI2, a pair of specific primers set was employed to amplify the total transcripts ofTaSSI2on A, B and D genomes (Appendix A).After treatment with MeJA, the expression level ofTaSSI2was gradually decreased at first 12 h and then became stable until 24 h (Fig. 4-A). However, the expression ofTaSSI2was up-regulated by SA and reached the peak at 12 h, with about 2.3 fold increase (Fig. 4-A). With ABA treatment, the expression ofTaSSI2increased at 6 h and then gradually recovered. No apparent changes of expression were found under IAA and ACC treatment (Fig. 4-A).
To unravel the regulation ofTaSSI2in FHB resistance,we examined thePRgenes expression in leaves 7 DPI of BSMV-VIGS. It was found thatTaPR4andTaPR5were up-regulated inTaSSI2-silenced leaves compared with the mock of BSMV:00 (Fig. 4-B).
3.5. Drought tolerance of TaSSI2 over-expressed Brachypodium
Lipids composition in cell membrane and wax is crucial for environmental stresses for plants. The drought tolerance is dependent on high levels of polyunsaturated fatty acids (PUFAs) and the ability to maintain fatty acid (FA)desaturation activity (Torres-Franklinet al. 2009). In order to investigate whetherTaSSI2is involved in drought tolerance, theTaSSI2-2ALwas over-expressed inB.distachyon(Bd21) and the transgenic lines were exposed in the 25%PEG10000 for drought tolerance evaluation. A total of three transgenic lines were confirmed to highly expressTaSSI2by semi-quantitative RT-PCR, withBdUBC18(Bradi4g00660)as internal control (Appendix B). In mock condition of 1/2MS medium, the wild-type and transgenic lines grew similarly proving the over-expressing ofTaSSI2did not affect the plant growth (Fig. 5-B). Under drought treatment, the wild-type plants became withered after 10 days after treatment and about 80% died at 15 days after planting (DAP) (Fig. 5-B),while all of the three transgenic lines showed enhanced tolerance compared with control at 15 DAP (Appendix A).The detail examination of line 2 (TaSSI2-2) showed that no significant growth difference was found between wild-type and transgenic plants, while the latter exhibited about 32 and 36% higher survival rate than wild-type plants at 10 and 15 DAP (Fig. 5-A and C). This phenotype is consistent with its expression in responsive to drought stimulation thatTaSSI2was induced as early as 6 h after treatment and reached the peak at 12 h with about 8-fold increase of transcript(Fig. 5-D). Due to the possible role ofTaSSI2in fatty acids unsaturation, the contents of stearic acid (18:0), oleic acid(18:1), linoleic (18:2) and linolenic acid (18:3) were then measured in the transgenic plants. The results showed that the relative ratio of linolenic acid (18:3) in total fatty acids was apparently increased in transgenic plants rather in wild-type plants, while the oleic acid (18:1) in wild-type plants was a bit higher (Fig. 5-F).
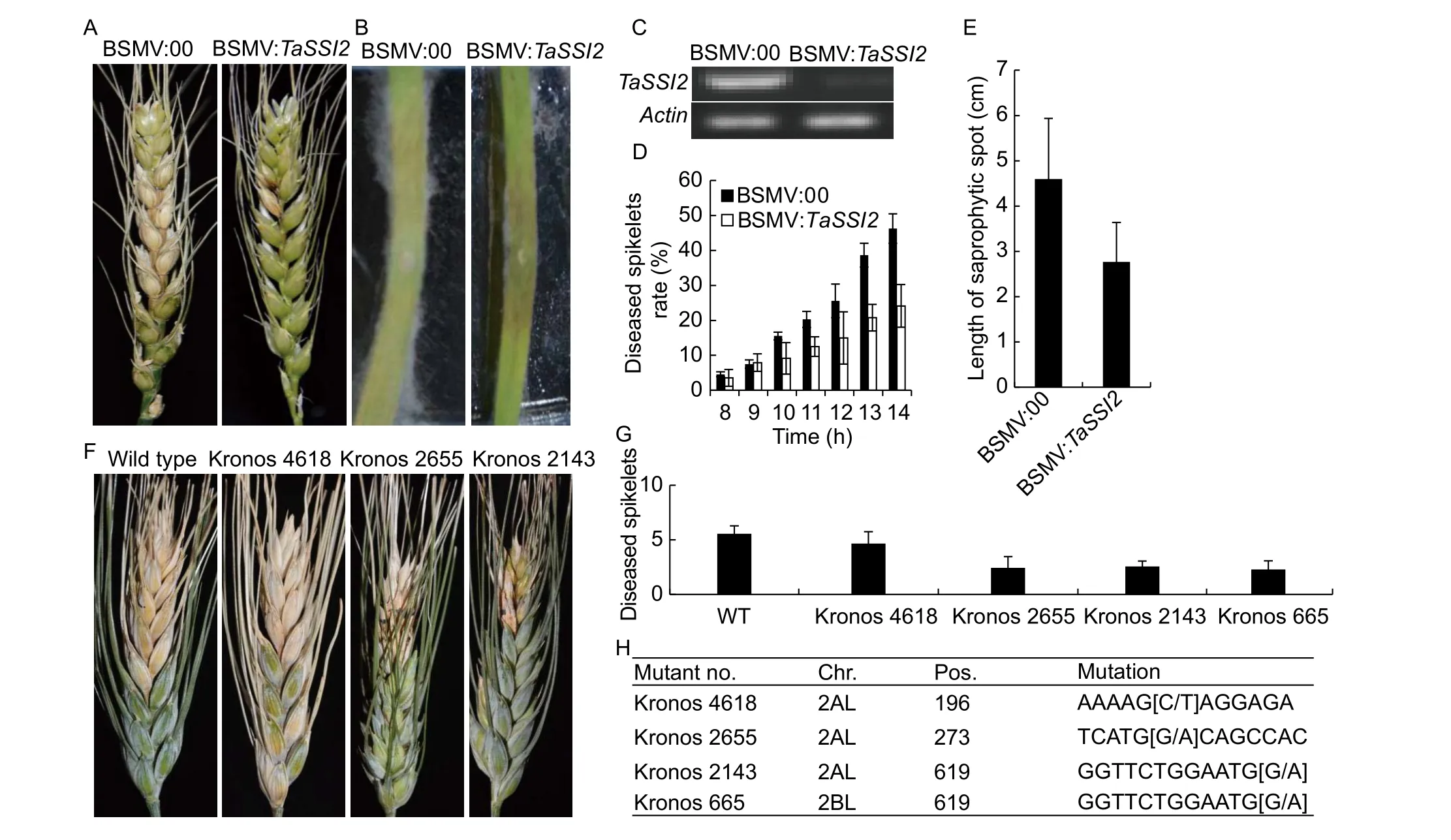
Fig. 3 Fusarium head blight (FHB) resistance of TaSSI2 by barley stripe mosaic virus (BSMV)-induced gene silencing (VIGS)and tilling mutants. A, phenotypes of TaSSI2-silenced spikes against Fusarium graminearum infection. B, phenotypes of TaSSI2-silenced leaves against F. graminearum infection. The central part of wheat third leaf was detached and cultured in vitro for F. graminearum treatment. C, semi-quantitative reverse transcription-PCR of TaSSI2 in BSMV-infected spikes. D, statistics of diseased spikelets of TaSSI2-silenced spikes compared with wild type plant of KN199. E, the saprophytic spot length (centimeter)of BSMV:00 and BSMV:TaSSI2. F, phenotype of Kronos tilling mutants against F. graminearum infection. G, statistics of diseased spikelet of Kronos tilling mutants. H, the mutation information of Kronos tilling mutants of TaSSI2. WT, wild type. Data are means±SD.
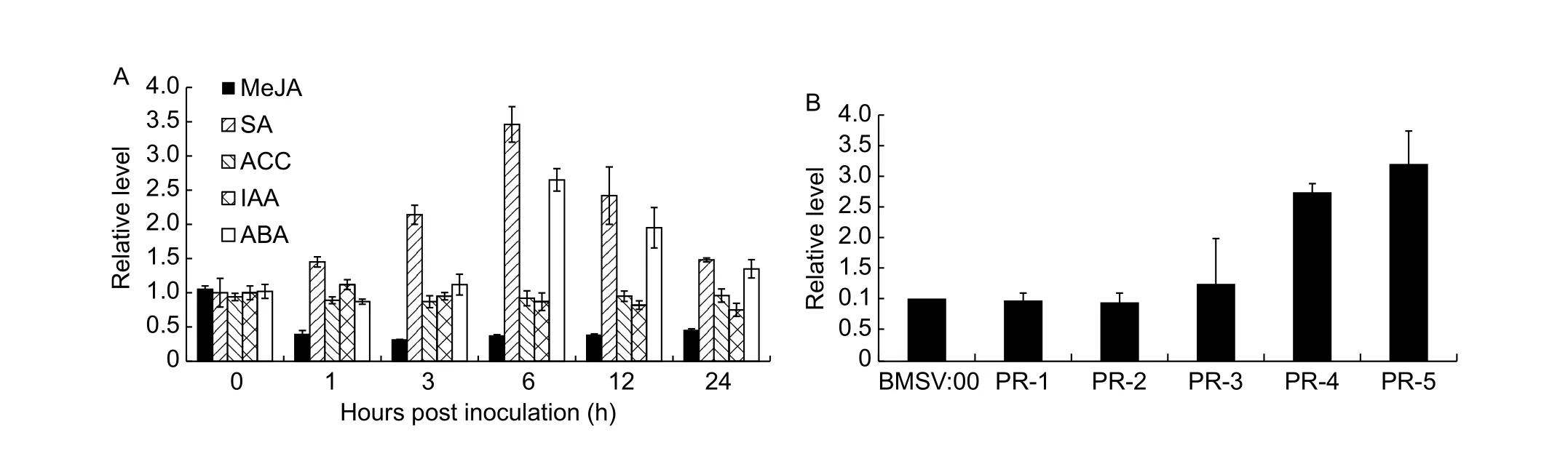
Fig. 4 Transcriptional regulation of TaSSI2. A, expression patterns of TaSSI2 against methyl jasmonate (MeJA), salicylic acid(SA), 1-amino cyclopropane-1-carboxy (ACC), indole acetic acid (IAA), and abscisic acid (ABA) treatments. The expression level was calculated by the comparative CT method with the wheat β-actin gene as the endogenous reference for normalization. The relative expression of TaSSI2 was calculated by comparing its transcript level to that in the mock controls across time points. B,the expression of pathogenesis-related (PR) genes was examined in the leaves of barley stripe mosaic virus (BSMV)-induced gene silencing (VIGS)-treated plants. The RNA was extracted by using leaves at 7 days post inoculation (DPI) BSMV. The relative expression level of PR genes was calculated compared with BSMV:00 as mock. Data are means±SD among 3 replicates.

Fig. 5 Drought tolerance of transgenic Brachypodium over-expressing TaSSI2. A, phenotype of TaSSI2 over-expression plants against drought condition. The plants were treated by 25% PEG10000 for 15 days and photographed. B, phenotype of TaSSI2 over-expression plants and wild types plants under mock condition. C, survival rate of TaSSI2 over-expression plants under drought condition. D, expression of TaSSI2 against drought treatment. E, the contents of stearic acid (18:0), oleic acid (18:1), linoleic(18:2), and linolenic acid (18:3) in wild-type and TaSSI2 over-expression plants. Data are means±SD.
3.6. Subcellular localization of the TaSSI2
It is generally believed that the subcellular localization of a protein is important to understand its cellular function. Therefore, we decided to examine the subcellular localization ofTaSSI2. We constructed the EGFP fusion proteins under the control of theCauliflower mosaic virus35S promoter and transitionally expressed the proteins inN.benthamianaleaves. For the negative control, the transient expression of empty:EGFP was observed in the whole cytosol with a weak signal. Sometimes, the fluorescence signal was also found in plasma-membrane and nuclei.While, TaSSI2:EGFP showed punctuate signals of varying sizes like some vesicles in cytosol (Fig. 6-A). In order to further determine the subcellular location of TaSSI2, co-localization with Golgi apparatus (G-rbCD3-968), chloroplast and plastids (pt-rbCD3-1000) were performed (Nelsonet al. 2007). TaSSI2 showed partially merged signals with the marker line of Golgi apparatus (Fig. 6-A). In order to confirm this result, we co-transformed the TaSSI2:EGFP and G-rbCD3-968 into the onion epidermis cells by bombardment, which produced the similar manner (Fig. 6-B).However, TaSSI2:EGFP didn’t show overlapped signal with chloroplast auto-fluorescence (Fig. 7-A), so with the plastids(data not shown). Thus, co-localization results showed that TaSSI2 may be associated with Golgi apparatus and secretory vesicles.
4. Discussion
FHB is becoming more serious in recent years, threating wheat production and food safety (McMullenet al. 1997).However, the resistant mechanism of plant cell is rather unknown due to rare resistant genes not successfully be cloned and analyzed (Kazanet al. 2012).
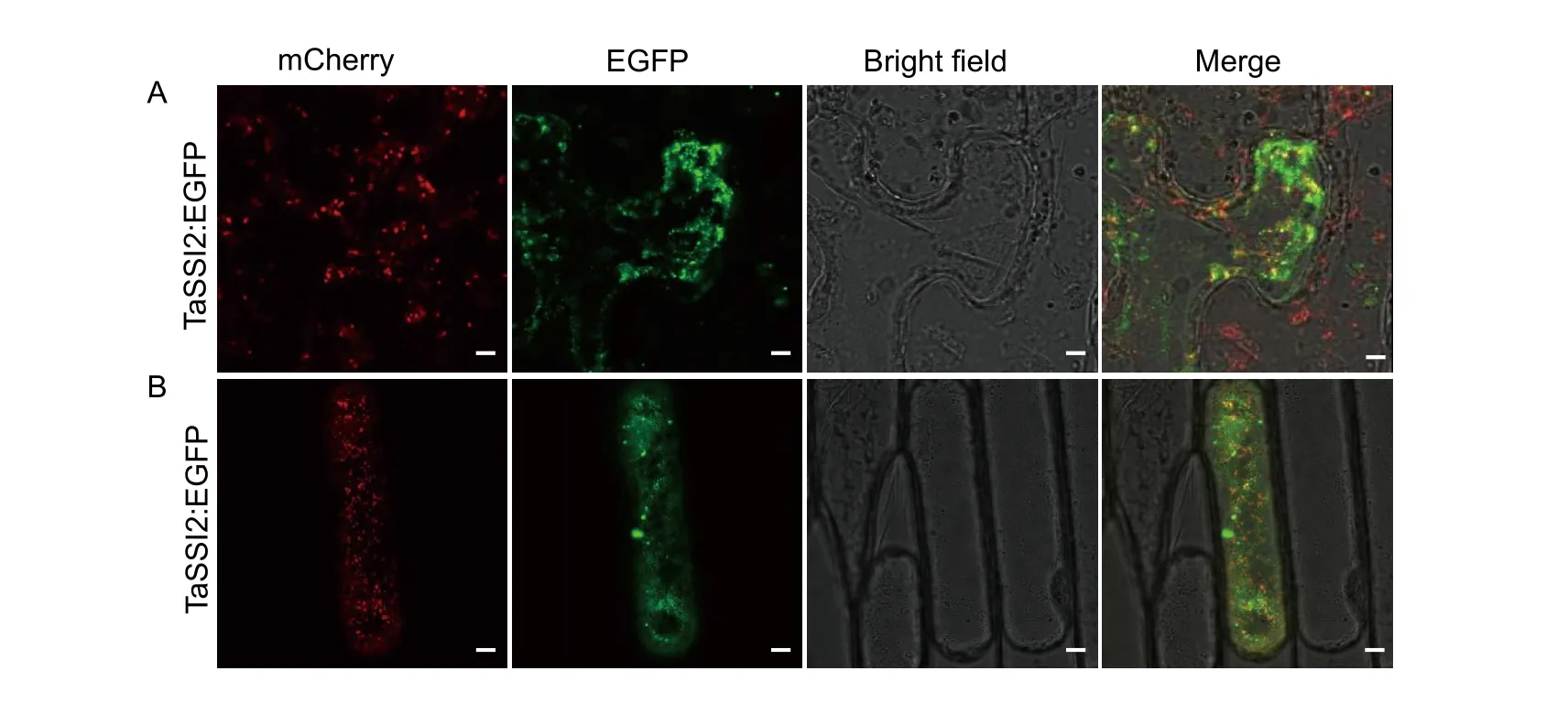
Fig. 6 Subcellular localization of TaSSI2. A, Agrobacterium strain GV3101 harboring construct of TaSSI2:EGFP was transiently expressed in Nicotiana leaves. B, TaSSI2:EGFP was transiently expressed in onion epidermis. The G-rbCD3-968 maker line representing Golgi apparatus was used for co-transfection with TaSSI2:EGFP, which was fused with mCherry for observation.Images were captured and merged by z-series optical sections after 3 days of agro-infiltration. Bar=20 μm.

Fig. 7 Co-localization of TaSSI2 in chloroplast. A, Agrobacterium strain GV3101 harboring construct of TaSSI2:EGFP was transiently expressed in Nicotiana leaves. The auto-fluorescence of chloroplast was used to merge with TaSSI2:EGFP. B, the bin vector containing 35S:EGFP was used as the control. Images were captured and merged by z-series optical sections after 3 days of agro-infiltration. Bar=20 μm.
As a hemi-biotrophic fungus, the pathogen infection pattern transits from biotrophic phase to necrotrophic phase as the disease develops. Such an infection pattern seems rather effective that little germplasm of wheat and barley have evolved major resistance toF.graminearum(Ruddet al. 2001). This might be partially related with antagonistic role of SA and JA signaling in plant cell. SA was often thought to induce expression of PR genes, production of ROS species, leading to hypersensitive response (HR)or systemic acquired resistance (SAR) in order to restrain biotrophic fungi or bacteria, however, JA was often thought to be involved in detoxification process and relieve the infection of necrotrophic pathogen (Makandaret al. 2012). Thus,the transition pattern of plant from SA to JA signaling might be crucial for FHB defence. In this study, we investigated the function of aSSI2gene in wheat, which might confer broad spectrum disease resistance. It was demonstrated that down-regulatingTaSSI2through BSMV-VIGS increased resistance toB.graminisf. sp.triticiand resistance to theF.graminearum. The negative regulation ofTaSSI2in FHB resistance was also proved by using tilling mutants of tetraploid wheat (Fig. 3). Suppressor of salicylate insensitivity ofnpr1–5(ssi2) mutant, which is defective in a gene encoding SACPD have been identified in a variety of plant genomes.Previously, mutation of theSSI2gene (AtSSI2) enhances the resistance to multiple pathogens, by constructively activation of SA dependent defense signaling (Shahet al.2001; Sekineet al. 2004). It also has been reported that over-expression of a wheatSACPDgene (TaSSI2-2DL)inArabidopsisssi2mutant compromise its resistance to powdery mildew (Golovinomyces cichoracearum), indicating a conserved biochemical functional betweenTaSSI2andAtSSI2(Songet al. 2013). Here, our result indicated that the activation of SA signaling may play important role for plant cells in resistance toF.graminearum. This is consistent with previous study that over-expression ofAtNPR1,a key regulator in SA signaling, in wheat could effectively suppress FHB symptom expanding to flanking spikelet(Makandaret al. 2006; Gaoet al. 2013). Here, we found that the expression ofTaSSI2was depressed by MeJA but positively regulated by SA, indicating the cross-talk between SA and JA in wheat is still to be elucidated.
InArabidopsis, it has been demonstrated that targeting ofSSI2to the chloroplasts is important for the rescue ofssi2-associated phenotypes (Kachrooet al. 2003). Besides,results from Mandal’s lab also showed that NOA1 and the 18:1 synthesizing SSI2 proteins were present inclose proximity within the nucleoids of chloroplasts (Mandalet al.2012). In this study, TargetP analysis predicted the presence of N-terminal transit peptides targeting the TaSSI2 to the chloroplast, mitochondria or secretory pathway (data not shown). The analysis of subcellular localization, by using tobacco and onion epidermis cells, showed that TaSSI2 may be associated with Golgi apparatus and secreted vesicles (Fig. 6). It may result from TaSSI2 translocation for functional activation. Studies from several laboratories suggest that secretory vesicle could carry proteins, lipids or polysaccharides to plasma membrane and extracellular space during certain physiological and environmental conditions (Leeet al. 2001). As a stearoyl-acyl carrier protein fatty acid desaturase, TaSSI2 might be involved in cell lipid metabolism and carry lipids targets out of the cell for membrane or wax synthesis. Actually, several lines of evidence have proved that cell plasma membrane, extra-cellular cuticle or wax is closely related with drought tolerance for plants (Klinkenberget al. 2014). Under drought stress, it was foundArabidopsisincreases the ratio of digalactosyl diglyceride (DGDG) to monogalactosyl diglyceride (MGDG)and FA unsaturation (Gigonet al. 2004). Recently, the expression of two stearoyl-acyl carrier protein fatty acid desaturase FAD3 and SAD6 was found to be stimulated in crown galls under hypoxia and drought condition, increasing the contents of unsaturated fatty acids and strongly suggested their involvement in drought tolerance (Klinkenberget al. 2014). In this study, the transgenicBrachypodiumover-expressingTaSSI2showed enhanced linolenic acid(18:3), but no apparent changes were observed for stearic acid (18:0) oleic acid (18:1) or linoleic (18:2). This is possible thatTaSSI2caused strong unsaturation reaction chain for fatty acid and resulted in a final increase linolenic acid,which is in agreement with the previous study to be involved in drought tolerance (Klinkenberget al. 2014).
5. Conclusion
ATaSSI2gene in wheat was proved to be involved in multiple biological functions including FHB, powdery mildew resistance and drought tolerance. The silencing or knock out of this gene resulted in enhanced FHB resistance by activating SA signaling. Moreover, the localization of TaSSI2 in Golgi apparatus and secreted vesicles support its role in translocation of lipids out of cell and increase the unsaturated fatty acids, linolenic acid (18:3), in plasma membrane or cuticle layer, participating in plant drought tolerance.
Acknowledgements
This work was supported by the National Natural Sci ence Foundation of China (3315203911, 31471488), the National Key Research and Development Program of China(2016YFD0100602), the Transgenic Spe cial Item, China(2016ZX08002003-002 and 2016ZX08009-003).
Appendicesassociated with this paper can be available on http://www.ChinaAgriSci.com/V2/En/appendix.htm
Bai G, Shaner G. 1994. Scab of wheat: Prospects for control.Plant Disease, 78, 760–766.
Bollina V, Kumaraswamy G K, Kushalappa A C, Choo T M,Dion Y, Rioux S, Faubert D, Hamzehzarghani H. 2010.Mass spectrometry-based metabolomics application to identify quantitative resistance-related metabolites in barley against Fusarium head blight.Molecular Plant Pathology,11, 769–782.
Bragg J N, Wu J, Gordon S P, Guttman M E, Thilmony R,Lazo G R, Gu Y Q, Vogel J P. 2012. Generation and characterization of the Western Regional Research Center Brachypodium T-DNA insertional mutant collection.PLoS ONE, 7, e41916.
Cahoon E, Shanklin J. 1997. Approaches to the design of acyl-ACP desaturases with altered fatty acid chain-length and double bond positional specificities. In: Williams J, Khan M, Lem N, eds.,Physiology,Biochemistry and Molecular Biology of Plant Lipids. Springer, the Netherlands. pp.374–376.
Chen X, Steed A, Travella S, Keller B, Nicholson P. 2009.Fusarium graminearumexploits ethylene signalling to colonize dicotyledonous and monocotyledonous plants.New Phytologist, 182, 975–983.
Das P, Ito T, Wellmer F, Vernoux T, Dedieu A, Traas J,Meyerowitz E M. 2009. Floral stem cell termination involves the direct regulation of AGAMOUS by PERIANTHIA.Development, 136, 1605–1611.
Ding L, Xu H, Yi H, Yang L, Kong Z, Zhang L, Xue S, Jia H,Ma Z. 2011. Resistance to hemi-biotrophicF.graminearuminfection is associated with coordinated and ordered expression of diverse defense signaling pathways.PLoS One, 6, e19008.
Emanuelsson O, Nielsen H, Brunak S, von Heijne G. 2000.Predicting subcellular localization of proteins based on their N-terminal amino acid sequence.Journal of Molecular Biology, 300, 1005–1016.
Ferrari S, Sella L, Janni M, De Lorenzo G, Favaron F, D’Ovidio R. 2012. Transgenic expression of polygalacturonaseinhibiting proteins inArabidopsisand wheat increases resistance to the flower pathogenFusarium graminearum.Plant Biology, 14, 31–38.
Gao C S, Kou X J, Li H P, Zhang J B, Saad A S I, Liao Y C.2013. Inverse effects ofArabidopsisNPR1gene on fusarium seedling blight and fusarium head blight in transgenic wheat.Plant Pathology, 62, 383–392.
Gigon A, Matos A R, Laffray D, Zuily-Fodil Y, Pham-Thi A T.2004. Effect of drought stress on lipid metabolism in the leaves ofArabidopsis thaliana(ecotype Columbia).Annals of Botany, 94, 345–351.
Hou W, Mu J, Li A, Wang H, Kong L. 2015. Identification of a wheat polygalacturonase-inhibiting protein involved in Fusarium head blight resistance.European Journal of Plant Pathology, 141, 731–745.
Jansen C, von Wettstein D, Schafer W, Kogel K H, Felk A,Maier F J. 2005. Infection patterns in barley and wheat spikes inoculated with wild-type and trichodiene synthase gene disruptedFusarium graminearum.Proceedings of the National Academy of Sciences of the United States of America, 102, 16892–16897.
Jiang C J, Shimono M, Maeda S, Inoue H, Mori M, Hasegawa M, Sugano S, Takatsuji H. 2009. Suppression of the rice fatty-acid desaturase geneOsSSI2enhances resistance to blast and leaf blight diseases in rice.Molecular Plant-Microbe Interactions, 22, 820–829.
Kachroo A, Fu D Q, Havens W, Navarre D, Kachroo P, Ghabrial S A. 2008. An oleic acid-mediated pathway induces constitutive defense signaling and enhanced resistance to multiple pathogens in soybean.Molecular Plant-Microbe Interactions, 21, 564–575.
Kachroo A, Kachroo P. 2009. Fatty acid-derived signals in plant defense.Annual Review of Phytopathology, 47, 153–176.
Kachroo A, Lapchyk L, Fukushige H, Hildebrand D, Klessig D,Kachroo P. 2003. Plastidial fatty acid signaling modulates salicylic acid- and jasmonic acid-mediated defense pathways in theArabidopsis ssi2mutant.The Plant Cell,15, 2952–2965.
Kachroo A, Shanklin J, Whittle E, Lapchyk L, Hildebrand D,Kachroo P. 2007. TheArabidopsisstearoyl-acyl carrier protein-desaturase family and the contribution of leaf isoforms to oleic acid synthesis.Plant Molecular Biology,63, 257–271.
Kachroo A, Venugopal S C, Lapchyk L, Falcone D, Hildebrand D, Kachroo P. 2004. Oleic acid levels regulated by glycerolipid metabolism modulate defense gene expression inArabidopsis.Proceedings of the National Academy of Sciences of the United States of America, 101, 5152–5157.Kachroo P, Shanklin J, Shah J, Whittle E J, Klessig D F. 2001.A fatty acid desaturase modulates the activation of defense signaling pathways in plants.Proceedings of the National Academy of Sciences of the United States of America, 98,9448–9453.
Kazan K, Gardiner D M, Manners J M. 2012. On the trail of a cereal killer: Recent advances inFusarium graminearumpathogenomics and host resistance.Molecular Plant Pathology, 13, 399–413.
Klinkenberg J, Faist H, Saupe S, Lambertz S, Krischke M,Stingl N, Fekete A, Mueller M J, Feussner I, Hedrich R,Deeken R. 2014. Two fatty acid desaturases, STEAROYLACYL CARRIER PROTEIN Delta9-DESATURASE6 and FATTY ACID DESATURASE3, are involved in drought and hypoxia stress signaling inArabidopsiscrown galls.Plant Physiology, 164, 570–583.
Lee H, Xiong L, Gong Z, Ishitani M, Stevenson B, Zhu J K.2001. TheArabidopsis HOS1gene negatively regulates cold signal transduction and encodes a RING finger protein that displays cold-regulated nucleo-cytoplasmic partitioning.Genes & Development, 15, 912–924.
Li G, Yen Y. 2008. Jasmonate and ethylene signaling pathway may mediate Fusarium head blight resistance in wheat.Crop Science, 48, 1888–1896.
Liu Z, Sun Q, Ni Z, Nevo E, Yang T. 2002. Molecular characterization of a novel powdery mildew resistance genePm30 in wheat originating from wild emmer.Euphytica,123, 21–29.
Ma X, Du X Y, Liu G J, Yang Z D, Hou W Q, Wang H W, Feng D S, Li A F, Kong L R. 2015. Cloning and characterization of a novel UDP-glycosyltransferase gene induced by DON from wheat.Journal of Integrative Agriculture, 14, 830–838.
Makandar R, Essig J S, Schapaugh M A, Trick H N, Shah J. 2006. Genetically engineered resistance to Fusarium head blight in wheat by expression ofArabidopsisNPR1.Molecular Plant-Microbe Interactions, 19, 123–129.
Makandar R, Nalam V, Chaturvedi R, Jeannotte R, Sparks A A, Shah J. 2010. Involvement of salicylate and jasmonate signaling pathways inArabidopsisinteraction withFusarium graminearum.Molecular Plant-Microbe Interactions, 23,861–870.
Makandar R, Nalam V J, Lee H, Trick H N, Dong Y, Shah J.2012. Salicylic acid regulates basal resistance to Fusarium head blight in wheat.Molecular Plant-Microbe Interactions,25, 431–439.
Maldonado A M, Doerner P, Dixon R A, Lamb C J, Cameron R K. 2002. A putative lipid transfer protein involved in systemic resistance signalling inArabidopsis.Nature, 419, 399–403.Mandal M K, Chandra-Shekara A C, Jeong R D, Yu K, Zhu S, Chanda B, Navarre D, Kachroo A, Kachroo P. 2012.Oleic acid-dependent modulation of NITRIC OXIDE ASSOCIATED1 protein levels regulates nitric oxidemediated defense signaling inArabidopsis.The Plant Cell,24, 1654–1674.
McMullen M, Jones R, Gallenberg D. 1997. Scab of wheat and barley: A re-emerging disease of devastating impact.Plant Disease, 81, 1340–1348.
Nelson B K, Cai X, Nebenführ A. 2007. A multicolored set ofin vivoorganelle markers for co-localization studies inArabidopsisand other plants.The Plant Journal, 51,1126–1136.
Okazaki Y, Saito K. 2014. Roles of lipids as signaling molecules and mitigators during stress response in plants.The Plant Journal, 79, 584–596.
Peraldi A, Beccari G, Steed A, Nicholson P. 2011.Brachypodium distachyon: A new pathosystem to study Fusarium head blight and other Fusarium diseases of wheat.BMC Plant Biology, 11, 100.
Pestka J J, Zhou H R, Moon Y, Chung Y J. 2004. Cellular and molecular mechanisms for immune modulation by deoxynivalenol and other trichothecenes: Unraveling a paradox.Toxicology Letters, 153, 61–73.
Qi P F, Balcerzak M, Rocheleau H, Leung W, Wei Y M, Zheng Y L, Ouellet T. 2016. Jasmonic acid and abscisic acid play important roles in host-pathogen interaction betweenFusarium graminearumand wheat during the early stages of Fusarium head blight.Physiological and Molecular Plant Pathology, 93, 39–48.
Qin Y, Ma X, Yu G, Wang Q, Wang L, Kong L, Kim W, Wang H W. 2014. Evolutionary history of trihelix family and their functional diversification.DNA Research, 21, 499–510.
Rawat N, Pumphrey M O, Liu S, Zhang X, Tiwari V K, Ando K,Trick H N, Bockus W W, Akhunov E, Anderson J A, Gill B S.2016. WheatFhb1encodes a chimeric lectin with agglutinin domains and a pore-forming toxin-like domain conferring resistance to Fusarium head blight.Nature Genetics, 48,1576–1580.
Rocha O, Ansari K, Doohan F M. 2005. Effects of trichothecene mycotoxins on eukaryotic cells: A review.Food Additives &Contaminants, 22, 369–378.
Rudd J C, Horsley R D, McKendry A L, Elias E M. 2001. Host plant resistance genes for Fusarium head blight: Sources,mechanisms, and utility in conventional breeding systems.Crop Science, 41, 620–627.
Sekine K T, Nandi A, Ishihara T, Hase S, Ikegami M, Shah J,Takahashi H. 2004. Enhanced resistance toCucumber mosaic virusin theArabidopsis thalianassi2mutant is mediatedviaan SA-independent mechanism.Molecular Plant-Microbe Interactions, 17, 623–632.
Shah J, Kachroo P, Nandi A, Klessig D F. 2001. A recessive mutation in theArabidopsisSSI2gene confers SA- and NPR1-independent expression of PR genes and resistance against bacterial and oomycete pathogens.The Plant Journal, 25, 563–574.
Shanklin J, Somerville C. 1991. Stearoyl-acyl-carrier-protein desaturase from higher plants is structurally unrelated to the animal and fungal homologs.Proceedings of the National Academy of Sciencesof the United States of America, 88,2510–2514.
Shen X, Kong L, Ohm H. 2004. Fusarium head blight resistance in hexaploid wheat (Triticum aestivum)-Lophopyrumgenetic lines and tagging of the alien chromatin by PCR markers.Theoretical and Applied Genetics, 108, 808–813.
Song N, Hu Z, Li Y, Li C, Peng F, Yao Y, Peng H, Ni Z, Xie C, Sun Q. 2013. Overexpression of a wheat stearoyl-ACP desaturase (SACPD) geneTaSSI2inArabidopsisssi2mutant compromise its resistance to powdery mildew.Gene, 524, 220–227.
Sorahinobar M, Niknam V, Ebrahimzadeh H, Soltanloo H,Behmanesh M, Enferadi S T. 2015. Central role of salicylic acid in resistance of wheat againstFusarium graminearum.Journal of Plant Growth Regulation, 35, 477–491.
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. 2011. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods.Molecular Biology and Evolution, 28, 2731–2739.
Torres-Franklin M L, Repellin A, Huynh V B, d’Arcy-Lameta A, Zuily-Fodil Y, Pham-Thi A T. 2009. Omega-3 fatty acid desaturase (FAD3, FAD7, FAD8) gene expression and linolenic acid content in cowpea leaves submitted to drought and after rehydration.Environmental and Experimental Botany, 65, 162–169.
Toyooka K, Goto Y, Asatsuma S, Koizumi M, Mitsui T, Matsuoka K. 2009. A mobile secretory vesicle cluster involved in mass transport from the Golgi to the plant cell exterior.The Plant Cell, 21, 1212–1229.
Xiang Y, Song M, Wei Z, Tong J, Zhang L, Xiao L, Ma Z,Wang Y. 2011. A jacalin-related lectin-like gene in wheat is a component of the plant defence system.Journal of Experimental Botany, 62, 5471–5483.
Xiao J, Jin X, Jia X, Wang H, Cao A, Zhao W, Pei H, Xue Z, He L, Chen Q, Wang X. 2013. Transcriptome-based discovery of pathways and genes related to resistance against Fusarium head blight in wheat landrace Wangshuibai.BMC Genomics, 14, 197.
Yu G, Hou W, Du X, Wang L, Wu H, Zhao L, Kong L, Wang H. 2014. Identification of wheat non-specific lipid transfer proteins involved in chilling tolerance.Plant Cell Reports,33, 1757–1766.
Yuan C, Li C, Yan L, Jackson A O, Liu Z, Han C, Yu J, Li D.2011. A high throughputBarley stripe mosaic virusvector for virus induced gene silencing in monocots and dicots.PLoS ONE, 6, e26468.
Zhang X L, Shen X R, Hao Y F, Cai J J, Ohm H, Kong L R.2011. A genetic map ofLophopyrum ponticumchromosome 7E, harboring resistance genes to Fusarium head blight and leaf rust.Theoretical and Applied Genetics, 122, 263–270.
猜你喜欢
杂志排行

Journal of Integrative Agriculture的其它文章
- Rapid mapping of candidate genes for cold tolerance in Oryza rufipogon Griff. by QTL-seq of seedlings
- A dCAPS marker developed from a stress associated protein gene TaSAP7-B governing grain size and plant height in wheat
- A major quantitative trait locus controlling phosphorus utilization efficiency under different phytate-P conditions at vegetative stage in barley
- Overexpression of IbSnRK1 enhances nitrogen uptake and carbon assimilation in transgenic sweetpotato
- Collision detection of virtual plant based on bounding volume hierarchy: A case study on virtual wheat
- lntegrated management strategy for improving the grain yield and nitrogen-use efficiency of winter wheat