盐渍化地区刺槐、新疆杨和柽柳林中AM真菌特性及其影响因子
2018-01-31张新璐陈雪冬
张新璐,唐 明,陈雪冬,齐 梅,陶 晶,盛 敏
(1. 西北农林科技大学 林学院,陕西杨凌 712100;2. 西北农林科技大学 生命科学学院,陕西杨凌 712100)
丛枝菌根(Arbuscular Mycorrhizal, AM)真菌是重要的土壤生物成员之一,其可通过扩大植物根系吸收面积、活化土壤养分、增加植物对矿质元素和水分的吸收以及激素调节等作用来提高植物的抗逆性[1-3]。许多研究者认为生境异质性是影响微生物活性与功能的重要因素[4]。对菌根真菌而言,原始生境决定了其菌株特性,菌株来源不同其共生特性和抗逆性也往往不同,从而确定生境异质性对AM真菌特性的影响将为菌根技术的广泛应用提供保障[5]。
土壤盐渍化是全球普遍存在的资源和生态问题,严重制约着中国农林业的发展和中国生态环境建设。中国盐渍土总面积约0.991亿hm2,主要分布在中国西北、东北、华北、黄淮海平原以及沿海地区[6]。已有研究表明,AM真菌在盐渍土中分布广泛,且其与植物共生能显著提高植物的抗盐性,然而其抗盐性的高低与生境(植物种类、气温、降雨量、土壤理化及生物学性质)有关[7-9]。为充分发挥AM真菌在盐渍土改良中的作用,确定盐渍化地区AM真菌特性的关键影响因素已成为当前首要解决的基本问题。
据统计,中国西北地区盐渍土约占全国盐渍土总面积的60%左右,是中国受盐渍化影响最严重的区域之一[10]。在该地区,经过长期的自然选择和人工培育,一些树木已能进行生长,如刺槐(RobiniapseudoacaciaL.)、新疆杨(PopulusalbaL. var.pyramidalisBge.)和柽柳(TamarixchinensisL.)等[11]。它们不仅可以保持水土、防风固沙、造林绿化,也被广泛用于盐渍土改良,是盐渍化地区难得的绿化造林树种。目前研究发现,刺槐、新疆杨和柽柳均能与AM真菌形成共生结构[12-14],然而关于这些树种的AM真菌在不同盐渍化地区是否存在生境异质性尚不明确。因此,本试验选择位于中国西北部的3个盐渍化地区(内蒙古磴口、宁夏平罗和甘肃敦煌)中的刺槐、新疆杨和柽柳为研究对象,分析不同树种根内AM真菌侵染状况及根际AM真菌的生长和繁殖水平,探明AM真菌生长及共生特性与树种、气候条件和土壤因子间的相互关系,为菌根技术在盐渍土改良中的高效利用提供理论指导。
1 材料与方法
1.1 样品采集
分别于内蒙古磴口、宁夏平罗和甘肃敦煌盐渍化地区的刺槐、新疆杨和柽柳林中设5个20 m×20 m的小样方,每个小样方内随机选取5株植物,每株按东西南北4个方位,去除5 cm厚的表层土后,在5~20 cm土层范围内采集植物根系和根际土,然后分别将根系和根际土混合作为该样方的代表性样品。所选样区的地理位置和气候条件见表1。
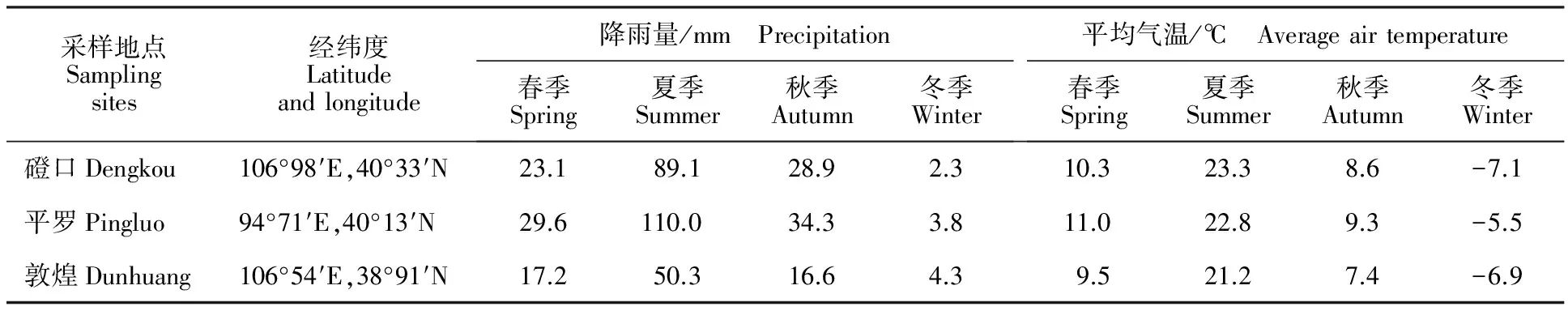
表1 采样点概况Table 1 Location, precipitation and average air temperature of sampling sites
1.2 测定项目及方法
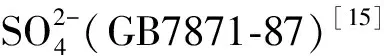
采用高锰酸钾滴定法测定过氧化氢酶活性[17];采用磷酸苯二钠比色法测定碱性磷酸酶活性[17];采用苯酚钠-次氯酸钠比色法测定脲酶活性[17];采用3,5-二硝基水杨酸(DNS)比色法测定蔗糖酶活性[17];采用三苯基四氮唑氯化物(TTC)比色法测定脱氢酶活性[18]。
1.2.2 土壤中总球囊酶素和易提取球囊酶素质量分数 采用Wright等[19]的方法测定土壤中易提取球囊霉素和总球囊霉素的质量分数。
1.2.3 AM真菌侵染率 根系样品采用透明压片法制片,镜检丛枝、泡囊和菌丝等结构特征,采用十字交叉法测定丛枝、泡囊、菌丝及AM真菌总的侵染率[20]。
1.2.4 AM真菌孢子密度和菌丝密度 孢子密度测定方法:分别从各土壤样品中取100 g鲜土2份,一份在105 ℃下烘干至恒质量,测定土壤含水量,另一份用湿筛倾析法分离孢子,在解剖镜下分格计数,计算AM真菌的孢子密度[21]。
土壤含水量=(土壤湿质量-土壤干质量)/土壤湿质量×100%
孢子密度= 孢子总数/[土壤湿质量×(1-土壤含水量)]
菌丝密度测定方法:采用抽滤法测定土壤中AM真菌的菌丝密度[22]。
菌丝密度=11/14×总交叉点数×网格单元格长度×滤膜上样块面积/[网格面积×土样湿质量×(1-土壤含水量)]
1.3 统计分析
采用R语言的Vegan程序包对试验数据进行统计分析[23]。
2 结果与分析
2.1 不同盐渍化地区刺槐、新疆杨及柽柳林的土壤理化性质
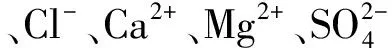
此外,研究还发现盐渍化地区林地土壤理化性质与其所处地理位置有关(Pseudo-F= 8.07,P<0.01)。利用vegdist()函数计算各盐渍化地区土壤理化性质间的相异性,并将其相异性与各盐渍化地区间的间隔距离进行相关分析。分析结果表明,不同盐渍化地区林地土壤理化性质的相异性与其间隔距离呈显著正相关(r=0.21,P<0.001),即林地间隔越远,其土壤理化性质差异越大(图2)。
2.2 不同盐渍化地区刺槐、新疆杨和柽柳林中土壤酶的活性
方差分析结果表明,土壤酶活性在不同树种间无显著差异,但同一树种在不同盐渍化地区间具有一定的差异(表3)。其中,刺槐、新疆杨和柽柳林中土壤过氧化氢酶活性以及柽柳林中土壤碱性磷酸酶和脱氢酶活性为磴口最低;柽柳林中土壤脲酶活性为平罗最低;新疆杨林中土壤蔗糖酶活性为平罗最高,磴口最低。
2.3 不同盐渍化地区刺槐、新疆杨和柽柳林中AM真菌的特性
研究发现,同一树种的AM真菌特性在不同盐渍化地区间表现出一定的差异(表4)。具体而言,刺槐和新疆杨根际土中AM真菌的孢子密度为磴口最高;刺槐根际土中AM真菌的菌丝密度为敦煌最高,而柽柳根际土中AM真菌的菌丝密度则为平罗最高;新疆杨根系的菌丝侵染率为平罗最高,而其丛枝侵染率则为敦煌最高。
“*”表示在0.05水平下差异显著 “*” represents significant difference at 0.05 level
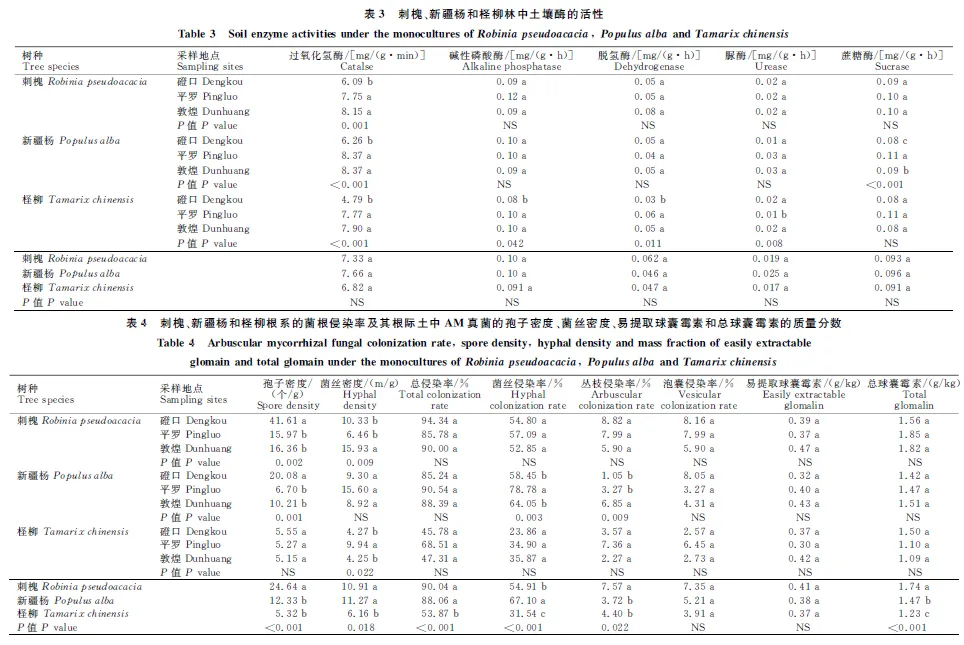

图2 不同盐渍化地区土壤理化性质的相异性与其间隔距离间的相互关系Fig.2 Correlation between dissimilarities of soil physico-chemical properties and geographic distances among different saline areas
此外,方差分析结果表明,树种不同其根内AM真菌的总侵染率、菌丝侵染率、丛枝侵染率及其根际土中AM真菌的孢子密度、菌丝密度和总球囊霉素质量分数显著不同,而根际土中易提取球囊霉素质量分数及根内泡囊侵染率在不同树种间无显著性差异(表4)。其中,刺槐根际土中AM真菌的孢子密度和总球囊霉素质量分数及根内AM真菌的总侵染率和丛枝侵染率最高,新疆杨根际土中AM真菌的菌丝密度及根内菌丝侵染率最高,而柽柳林中AM真菌的各项指标均相对较低。
2.4 土壤性质、树种和气候条件对AM真菌特性的影响
选择生境指标(土壤因子、树种及气候条件)为解释变量,利用varpart()变差分析函数比较土壤因子、树种及气候条件对AM真菌特性的贡献程度。变差分析结果表明土壤因子、树种及气候条件共解释了61.8%的变差,其中土壤因子单独解释或与树种或与气候条件一起共解释了53.9%的变差,说明土壤因子对AM真菌特性的影响最大,且其影响与树种和气候条件密切相关(图3)。
2.5 刺槐、新疆杨和柽柳林中AM真菌特性与土壤因子和气候条件的关系
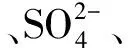
图3 土壤因子、树种和气候条件对AM真菌特性影响的变差分析Fig.3 Variation partitioning of arbuscular mycorrhizal fungal attributes by soil factors, tree species and climate conditions

(1)节点表示各AM真菌、土壤因子及气候指标;连线表示连线两端的节点具有显著相关性(P<0.05);连线上的数值为Pearson相关系数 The nodes represent soil, climate and AM fungal parameters, the edge represent a significant (P<0.05) relationship between the nodes, and the number on each edge is the Pearson’s correlation coefficient;(2)SD.AM真菌的孢子密度 AM fungal spore density;HD.菌丝密度 Hyphal density;HC.菌丝侵染率 Hyphal colonization rate;AC.丛枝侵染率 Arbuscular colonization rate;Pspring、Psummer、Pautum和Pwinter.分别表示春、夏、秋、冬季降雨量 Precipitation in spring, summer, autumn and winter;Tspring、Tsummer、Tautum和Twinter.分别表示春、夏、秋、冬季平均气温 Average air temperature in spring, summer, autumn and winter
图4刺槐(A)、新疆杨(B)和柽柳(C)林中AM真菌特性与土壤因子和气候条件的关系网络图
Fig.4NetworksofcorrelationsamongAMfungi,soilfactorsandclimateconditionsofRobiniapseudoacacia(A),Populusabla(B)andTamarixchinensis(C)
3 讨 论
微生物趋于生境选择,其生物活性、功能及其差异是反映生境异质性的重要指标[24]。从整体看,中国西北地区盐渍土中刺槐、新疆杨和柽柳根内AM真菌的侵染力及根际土中AM真菌的生长与繁殖能力均具有一定的生境异质性。选择与生境有关的土壤因子、树种及气候条件进行变差分析发现,AM真菌特性的大部分变差由土壤因子单独解释或与树种或与气候条件一起共同解释,说明土壤因子对AM真菌特性的影响最大,且其影响与树种和气候条件密切相关。该结论与一些研究者在其他一些自然生态系统中所获得的结果基本相似[25-26]。
Carvalho等[8]研究认为气温和降雨量的变化可以直接影响盐渍土中AM真菌的侵染率和孢子密度。一般来讲,降雨可通过改变土壤湿度、调节植物蒸腾速率对AM真菌进行影响;而气温一方面可直接影响AM真菌的生长速度,也可通过调节植物生长而对AM真菌进行间接影响[32]。本研究结果表明,盐渍化地区刺槐、新疆杨和柽柳根系AM真菌的侵染状况以及根际AM真菌的菌丝生长和产孢量均与气温和降雨量有关,且树种不同其AM真菌特性与气温和降雨量间的相互关系也有所不同。例如刺槐根际AM真菌的菌丝密度与春夏秋冬季气温和夏秋季降雨量呈显著负相关,而柽柳根际AM真菌的菌丝密度与春秋冬季气温和春夏季降雨量呈显著正相关。Kabir等[33]在农田生态系统中也发现AM真菌的菌丝密度与气候条件有关。综上,盐渍化地区AM真菌特性具有生境异质性,且其异质性主要由土壤因子、树种及气候条件等生境因子的变化所致,然而关于这些生境因子对AM真菌的影响机制尚不完全清楚,需待进一步深入的研究。
Reference:
[1] POZO M J,JUNG S C,MARTNEZ-MEDINA A,etal.Root allies:Arbuscular Mycorrhizal Fungi Help Plants to Cope with Biotic Stresses∥Symbiotic Endophytes[M].Heidelberg Berlin:Springer,2013:289-307.
[2] 许平辉,王飞权,齐玉岗,等.丛枝菌根真菌对茶树抗旱性的影响[J].西北农业学报,2017,26(7):1033-1040.
XU P H,WANG F Q,QI Y G,etal.Effect of arbuscular mycorrhiza fungi on drought resistance in tea plant(Camelliasinensis) [J].ActaAgricultureBoreali-occidentalisSinica,2017,26(7):1033-1040.
[3] LIU H G,WANG Y J,HART M,etal.Arbuscular mycorrhizal symbiosis regulates hormone and osmotic equilibrium ofLyciumbarbarumL. under salt stress[J].Mycosphere,2016,7(6):828-843.
[4] FIERER N,JACKSON R B.The diversity and biogeography of soil bacterial communities[J].ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,2006,103(3):626-631.
[5] ESTRADA B,BAREA J M,AROCA R,etal.A nativeGlomusintraradicesstrain from a Mediterranean saline area exhibits salt tolerance and enhanced symbiotic efficiency with maize plants under salt stress conditions[J].PlantandSoil,2013,366(1-2):333-349.
[6] 张建锋.盐碱地的生态修复研究[J].水土保持研究,2008,15(4):74-78.
ZHANG J F.Discussion on ecological rehabilitation of salt-affected soils[J].ResearchofSoilandWaterConservation,2008,15(4):74-78.
[7] 刘润进,刘鹏起,徐 坤,等.中国盐碱土壤中AM菌的生态分布[J].应用生态学报,1999,10(6):721-724.
LIU R J,LIU P Q,XU K,etal.Ecological distribution of arbuscular mycorrhizal fungi in saline-alkaline soils of China[J].ChineseJournalofAppliedEcology,1999,10(6):721-724.
[8] CARVALHO L M,CACADOR I,MARTINS-LOUCO M A.Temporal and spatial variation of arbuscular mycorrhizas in salt marsh plants of the Tagus estuary(Portugal)[J].Mycorrhiza,2001,11(6):303-309.
[9] GUO X H,GONG J.Differential effects of abiotic factors and host plant traits on diversity and community composition of root-colonizing arbuscular mycorrhizal fungi in a salt-stressed ecosystem[J].Mycorrhiza,2014,24(2):79-94.
[10] 汪 林,甘 泓,于福亮,等.西北地区盐渍土及其开发利用中存在问题的对策[J].水利学报,2001(6):90-95.
WANG L,GAN H,YU F L,etal.Salted soil and its development in northwest China[J].JournalofHydraulicEngineering,2001(6):90-95.
[11] 林士杰,张忠辉,张大伟,等.盐碱地树种选择及抗盐碱造林技术研究进展[J].中国农学通报,2012,28(10):1-5.
LIN SH J,ZHANG ZH H,ZHANG D W,etal.Research progress on the trees species choice in saline-alkali land and drought resistant forestation technology[J].ChineseAgriculturalScienceBulletin,2012,28(10):1-5.
[12] SHENG M,CHEN X D,ZHANG X L,etal.Changes in arbuscular mycorrhizal fungal attributes along a chronosequence of black locust(Robiniapseudoacacia) plantations can be attributed to the plantation-induced variation in soil properties[J].ScienceoftheTotalEnvironment,2017(599):273-283.
[13] 梁倩倩,李 敏,刘润进,等.全球变化下菌根真菌的作用及其作用机制[J].生态学报,2014,34(21):6039-6048.
LIANG Q Q,LI M,LIU R J,etal.Function and functioning mechanisms of mycorrhizal fungi under global changes[J].ActaEcologicaSinica,2014,34(21):6039-6048.
[14] 盛 敏,唐 明,迪丽努尔,等.西北盐碱土主要植物丛枝菌根研究[J].西北农林科技大学学报(自然科学版),2007,35(2):74-78.
SHENG M,TANG M,DILNUR,etal.Study on arbuscular mycorrhizae of common halophytes from northwest sali-alkali soil[J].JournalofNorthwestA&FUniversity(NaturalScienceEdition),2007,35(2):74-78.
[15] 鲍士旦,江荣风,杨超光,等.土壤农化分析[M].北京:中国农业出版社,2000.
BAO SH D,JIANG R F,YANG CH G,etal.Analysis of Soil Characteristics[M].Beijing:China Agriculture Press,2000.
[16] 陈云增,陈志凡,马建华,等.沙颍河流域典型癌病高发区土壤硝态氮对地下水和蔬菜硝酸盐积累的影响[J].环境科学学报,2016,36(3):990-998.
CHEN Y Z,CHEN ZH F,MA J H,etal.Effects of soil nitrate nitrogen on the nitrate accumulation in groundwater and vegetables in a typical high cancer incidence area of Shaying River basin[J].ActaScientiaeCircumstantiae,2016,36(3):990-998.
[17] 关松荫.土壤酶及其研究法[M].北京:农业出版社,1986.
GUAN S Y.Soil Enzymes and Its Method log[M].Beijing:Agricultural Press,1986.
[18] CHANDER K,BROOKES P C.Is the dehydrogenase assay invalid as a method to estimate microbial activity in copper-contaminated soils?[J].SoilBiologyBiochemistry,1991,23(10):909-915.
[19] WRIGHT S F,UPADHYAYA A.Extraction of an abundant and unusual protein from soil and comparison with hyphal protein of arbuscular mycorrhizal fungi[J].SoilScience,1996,161(9):575-586.
[20] PHILLIPS J M,HAYMAN D S.Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection[J].TransactionsoftheBritishMycologicalSociety,1970,55(1):158-168.
[21] 弓明钦,陈应龙,仲崇禄.菌根研究及应用[M].北京:中国林业出版社,1997.
GONG M Q,CHEN Y L,ZHONG CH L.Mycorrhizal Research and Application[M].Beijing:China Forestry Publishing House,1997.
[22] 何跃军,钟章成,董 鸣.AMF对喀斯特土壤枯落物分解和对宿主植物的养分传递[J].生态学报,2012,32(8):2525-2531.
HE Y J,ZHONG ZH CH,DONG M.Nutrients transfer for host plant and litter decompositon by AMF in Karst soil[J].ActaEcologicaSinica,2012,32(8):2525-2531.
[23] OKSANEN J,BLANCHET G F,KINDT R,etal.Vegan:community ecology package.R package version 2.0-10[J/OL].http:∥CRAN.R-project.org/package=vegan.[2013-06-02].
[24] ODLAND A,DEL MORAL R.Thirteen years of wetland vegetation succession following a permanent drawdown, Myrkdalen Lake, Norway[J].PlantEcology,2002,162(2):185-198.
[25] XU T L,VERESOGLOU S D,CHEN Y L,etal.Plant community, geographic distance and abiotic factors play different roles in predicting AMF biogeography at the regional scale in northern China[J].EnvironmentalMicrobiologyReports,2016,8(6):1048-1057.
[26] XU X H,CHEN C,ZHANG ZH,etal.The influence of environmental factors on communities of arbuscular mycorrhizal fungi associated withChenopodiumambrosioidesrevealed by MiSeq sequencing investigation[J].ScientificReports,2017,7(7):45134.
[27] DHAR P P,MRIDHA M A U.Arbuscular mycorrhizal associations in different forest tree species of Hazarikhil forest of Chittagong, Bangladesh[J].JournalofForestryResearch,2012,23(1):115-122.
[28] 张海波,梁月明,冯书珍,等.土壤类型和树种对根际土丛枝菌根真菌群落及其根系侵染率的影响[J].农业现代化研究,2016,37(1):187-194.
ZHANG H B,LIANG Y M,FENG SH ZH,etal.The effects of soil types and plant species on arbuscular mycorrhizal fungi community and colonization in the rhizosphere[J].ResearchofAgriculturalModernization,2016,37(1):187-194.
[29] LIU H G,WANG Y J,TANG M.Arbuscular mycorrhizal fungi diversity associated with two halophytesLyciumbarbarumL.andElaeagnusangustifoliaL.in Ningxia, China[J].ArchivesofAgronomyandSoilScience,2017,63(6):796-806.
[30] JOHNSON N C,GRAHAM J H,SMITH F A.Functioning of mycorrhizal associations along the mutualism-parasitism continuum[J].Newphytologist,1997,135(4):575-585.
[31] PHILIPPOT L,RAAIJMAKERS J M,LEMANCEAU P,etal.Going back to the roots:the microbial ecology of the rhizosphere[J].NatureReviewsMicrobiology,2013,11(11):789-799.
[32] 孙向伟,王晓娟,陈 牧,等.生态环境因子对AM真菌孢子形成与分布的作用机制[J].草业学报,2011,20(1):214-221.
SUN X W,WANG X J,CHEN M,etal.Effects of eco-environmental factors on the production and distribution of arbuscular mycorrhizal fungal spores[J].ActaPrataculturaeSinica,2011,20(1):214-221.
[33] KABIR Z, O′HALLORAN I P, FYLES J W,etal.Seasonal changes of arbuscular mycorrhizal fungi as affected by tillage practices and fertilization:hyphal density and mycorrhizal root colonization[J].PlantandSoil,1997,192(2):285-293.
