粳稻优势生态型不同世代恢复系的恢复效果
2018-01-31王坚,刘炜
王 坚, 刘 炜
(宁夏农林科学院 农作物研究所,宁夏永宁 750105)
杂交稻是利用两个遗传背景不同的亲本进行杂交,其杂交种在生活力、适应性、生殖力、生长势、抗逆性以及产量、品质等方面或整体水平上表现出优于两个亲本,杂交稻在生产中已被广泛利用。杂交粳稻通过“籼粳架桥”实现粳稻三系配套,在生产中推广应用[1]。但由于粳稻亲本间遗传基础狭窄,恢复系遗传基础贫乏、亲本老化,不育系不育细胞质比较单一,不育系和恢复系间遗传差异小、类型相似,缺乏遗传多样性,导致杂种优势不明显,在产量上难以大幅度超越常规粳稻[2]。邱福林等[3]认为北方杂交粳稻骨干亲本遗传差异有70%相似。杂种优势群是遗传基础丰富、共祖关系密切、主要特征特性趋向相近和一般配合力较强的自交系类群,群内无明显的杂种优势,而群之间可能获得强优势组合,可以有效拓宽亲本间的遗传基础,提高杂种优势。在杂交玉米育种中,利用配合力和分子标记对杂种优势群划分[4-8],通过不同群杂交获得大面积推广品种[9-10]。谷子[11]、小麦[12-13]和油菜[14]等作物也利用杂种优势群来拓宽亲本间的遗传基础,提高杂种优势。粳稻通过杂种优势值划分并确定西北粳、台湾粳、日本粳和韩国粳为优势生态型[15-17],将这些优势生态型通过回交转育成恢复系,可以达到丰富粳型杂交稻恢复系的遗传基础。在转育过程中回交代数过多费时费工,配合力也可能下降[18]。如何用最少的回交次数得到最高的配合力,为此在回交选育过程中对不同回交世代与各不育系进行测恢和配合力鉴定,希望通过对回交各世代杂种优势分析,找到回交次数与配合力的关系,为杂种优势群回交转育恢复系提供最佳方案。
1 材料与方法
1.1 材料及种植方式
1.1.1 供试亲本材料 受体材料为粳型优势生态群中的台湾粳‘嘉农428’和‘嘉农育251’,恢复基因的供体材料为‘C418’;不育系为‘秋光A’ ‘宁粳16A’ ‘216A’ ‘552A’、‘宁粳23A’ ‘宁粳24A’和‘花43A’,对照为‘宁粳41号’。
1.1.2 供试回交材料 以供体材料‘C418’为父本,粳型优势生态群台湾粳‘嘉农428’和‘嘉农育251’为母本,在抽穗期人工去雄授粉,得到F1代种子。再以‘嘉农428’、‘嘉农育251’为母本与各自F1为父本杂交,得到BC1F1代种子,继续种植BC1F1代种子和‘嘉农428’、‘嘉农育251’,在苗期对BC1F1代植株进行恢复基因分子检测,有恢复基因和无恢复基因植株个数卡方检测,选择符合1∶1的群体中有恢复基因的植株为父本继续与‘嘉农428’、‘嘉农育251’杂交得到BC2F1代种子。
1.1.3 供试杂交材料 以粳型优势生态群台湾粳‘嘉农428’、‘嘉农育251’回交的到BC1F1和BC2F1。在苗期进行恢复基因分子检测,有恢复基因和无恢复基因植株个数卡方检测,选择符合1∶1的群体中有恢复基因的植株为父本与不育系‘秋光A’ ‘宁粳16A’ ‘216A’ ‘552A’ ‘宁粳23A’ ‘宁粳24A’和‘花43A’杂交得到F1代种子。
1.1.4 种植方式 2012年夏季在宁夏开始回交,冬季在三亚加代,到2015年将各代杂交种种植于宁夏农林科学院农作物研究所的实验基地,4月中旬大棚育秧,5月中旬单株插秧,单株插秧行距27 cm,株距10 cm, 正常田间管理。
1.2 分子标记引物设计及PCR扩增方法
1.2.1 回交后代的恢复系分子标记辅助选择引物设计 根据 Rf1a基因位点保持系(rf1a/rf1a)比恢复系(Rf1a/Rf1a)缺少574 bp的片段,在缺失区域上游设计Rfa-7F:GGACCGGGGGATTTTACCTG,下游设计Rfa-7R:AACCCAACTGAGACCATGCC[19]。引物由上海生工生物工程技术服务有限公司合成。
1.2.2 DNA的提取和PCR扩增 在水稻3~4叶期取新鲜叶片,采用CTAB法提取DNA。PCR反应总体系体积为20 μL,其中10×Buffer(含Mg2+15 mmol/L) 2 μL,dNTP(各2.5 mmol/L)1 μL,100 μmol/L的引物各0.1 μL,1 UTaq酶,DNA模板1 μL,加ddH2O补足20 μL。PCR反应程序:94 ℃预变性5 min;94 ℃变性30 s,60 ℃退火30 s,72 ℃延伸40 s,共35个循环;最后72 ℃延伸10 min。扩增产物通过10 g/L 脂糖凝胶电泳,用Gold-View核酸染料染色后在紫外灯下观察,并用凝胶成像系统拍照。
1.3 农艺性状测定与育性鉴定
1.3.1 主要农艺性状的测定 回交各代材料与各不育系的杂交种和对照,大棚育秧,单株插秧,正常田间管理,成熟后单株带根取样,阴干对各材料的单株产量、株高、有效穗数、千粒质量、穗长、每穗粒数和结实率等主要农艺性状进行单株测定。
1.3.2 植株育性鉴定方法 在抽穗期对所有F1代材料进行单株观察,选择未开花的穗套袋。同时取穗用卡诺液固定,再用I2-KI法对各材料进行花粉染色镜检, 成熟时逐株考查自交结实率和自然结实率。以花粉可育率作为育性鉴定的主要指标,根据以上调查结果综合评价。花药瘦小、花粉败育和结实率低于5%的植株为不育株,反之为可育株。可育株数和不育植株个数进行卡方检测,选择符合1∶1的群体,在成熟时将可育株单株带根取样,阴干后考种。
1.4 数据处理
超对照优势计算公式:V=(F1-CK)/CK×100%,V为超对照伏势,F1为杂交种某一性状值,CK为对照某一性状值。采用Excel 2007对各农艺性状数据进行处理。
2 结果与分析
2.1 恢复基因分离检测
以‘嘉农428’、‘嘉农育251’为母本与各自F1为父本杂交,人工去雄,每穗只受同1植株父本花粉,得到BC1F1代种子为一个群体,在苗期用Rfa-7F/Rfa-7R引物对群体中每个植株进行PCR扩增,检测恢复基因,有恢复基因和无恢复基因植株个数见表1,按1∶1进行卡方检测,χ2最大为0.562 5,所有P>0.05,都符合1∶1分离。选取其中一个群体中有恢复基因植株继续为父本,继续与各自受体材料杂交得到BC2F1代群体,在苗期对群体中每个植株进行恢复基因分子检测,有恢复基因和无恢复基因植株个数,按1∶1进行卡方检测,结果χ2最大为0.235 3,所有P<0.05,BC2F1代群体有恢复基因和无恢复基因植株个数仍然都符合1∶1分离。
2.2 不同回交世代与各不育系杂种优势
2.2.1 ‘嘉农育251’不同回交世代与各不育系杂种优势 以‘C418’为供体,‘嘉农育251’为受体,连续回交到BC1F1和BC2F1代与不育系‘秋光A’ ‘宁粳16A’ ‘216A’ ‘552A’ ‘宁粳23A’ ‘宁粳24A’和‘花43A’杂交F1代平均单株产量见表2。由表2可以看出,‘嘉农育251’回交BC1F1代与各不育系杂交F1代平均单株产量均大于BC2F1代与各不育系杂交F1。BC1F1代中,与‘宁粳24A’平均单株产最高,‘秋光A’次之,BC2F1代中,与‘552A’平均单株产最高,BC1F1代中与‘宁粳24A’平均单株产比BC2F1代与‘宁粳24A’ 高15.27 g,BC1F1和BC2F1代与‘秋光A’的差距也较大(14.24 g),与‘花43A’和‘552A’的差距较小,分别为1.94 g和2.30 g。就标准差看除‘花43A’与BC1F1代杂交F1代单株产量的标准差小于BC2F1代杂交F1代,其余都是BC1F1代的F1代的大于BC2F1代,单株产量最大的其标准差也最大。从变异系数看,多数BC1F1代的杂交种变异系数大于BC2F1代的杂交种。BC1F1代与各不育系杂交的F1代群体中除‘宁粳23A’外,其他各不育系的单株最高产量高于BC2F1代与各不育系杂交F1代,BC1F1代与‘秋光A’和‘宁粳24A’的杂交的F1代单株产量中最高达118.70 g,BC2F1代只有75.50 g。就超对照优势看,BC2F1代与各不育系杂交F1代平均单株产量只有‘552A’高于对照,而BC1F1代与‘秋光A’ ‘宁粳16A’ ‘216A’ ‘552A’和‘宁粳24A’杂交F1代都高于对照。BC1F1代与各不育系杂交F1代超对照植株比例除‘216A’略低于BC2F1代的单株产量,其余都高于BC2F1代与这些不育系杂交F1代,超对照10%的植株比例除‘花43’也是 BC1F1的杂交种低于BC2F1的杂交种,其余BC1F1代与各不育系杂交F1代超对照10%的植株比例都高与BC2F1。
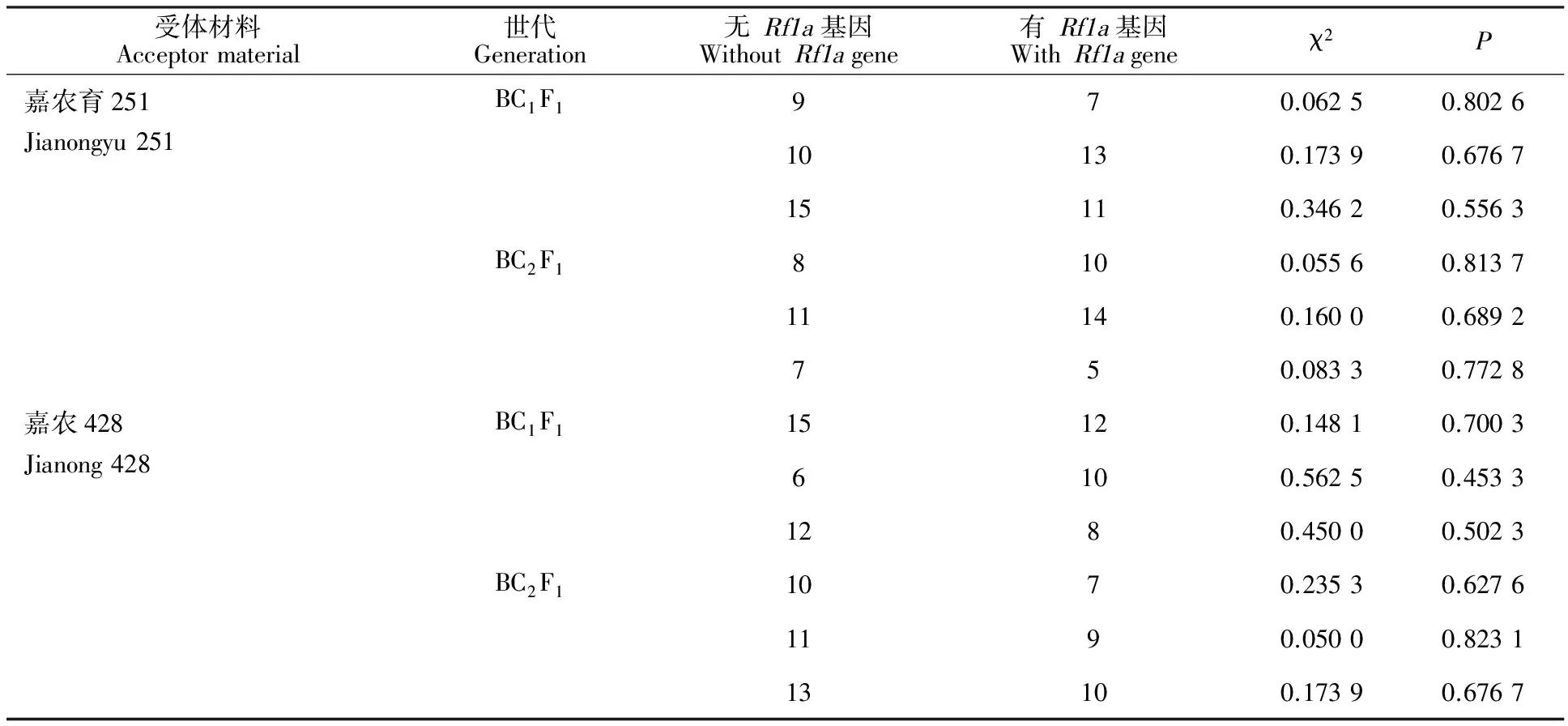
表1 Rf1a基因回交分离卡方检测Table 1 Chi-square testing on Rf1a gene backcross and segregation

表2 ‘嘉农育251’不同回交代数与各不育系杂种优势Table 2 Yield heterosis between ‘Jianongyu 251’ backcrossed generations and different sterile lines
2.2.2 ‘嘉农育428’ 不同回交世代与各不育系杂种优势 ‘嘉农428’回交BC1F1、BC2F1代与各不育系杂交F1代平均单株产量见表3。从表3 可以看到F1代都表现出一定的杂种优势,超对照优势最高达62.0%,超对照优势100%的比例最高达到71.0%;除‘宁粳16A’与‘嘉农428’回交BC1F1代杂交F1代平均单株产量低于BC2F1代杂交F1代平均单株产量,其他不育系‘秋光A’ ‘216A’ ‘552A’ ‘宁粳23A’ ‘宁粳24A’和‘花42A’ 与‘嘉农428’回交BC1F1代杂交F1代平均单株产量都高于与BC2F1代杂交F1代平均单株产量。‘嘉农428’回交BC1F1代和BC2F1代与‘秋光A’和‘宁粳16A’杂交F1代平均单株产量接近,BC2F1代杂交F1代平均单株产量的标准差和变异系数大于BC1F1代,其他不育系‘216A’ ‘552A’ ‘宁粳23A’ ‘宁粳24A’和‘花43A’与BC1F1代杂交F1代平均单株产量远高于BC2F1代杂交F1代平均单株产量,而且标准差和变异系数也大幅度的高于BC2F1代。除‘宁粳16A’ 外,BC1F1代与各不育系杂交F1单株产量最高值均大于BC2F1代杂交F1,BC1F1代与‘216A’杂交F1最高达160.80 g,与‘花43A’杂交F1最高为119.20 g,BC2F1代与各不育系杂交F1相对小较多,其中最大的与‘宁粳16A’杂交F1只有58.60 g。就超对照优势看只有‘216A’ ‘552A’ ‘宁粳23A’ 和‘花43A’与BC1F1代杂交F1代平均单株产量有超对照优势,BC2F1代与各不育系杂交F1平均单株产量无超对照优势,但单株有些超对照还有部分超过对照10%。大多数BC1F1代与各不育系杂交F1单株产量超对照植株比例高于BC2F1代,超过对照10%也是大多数BC1F1代的高于BC2F1代。
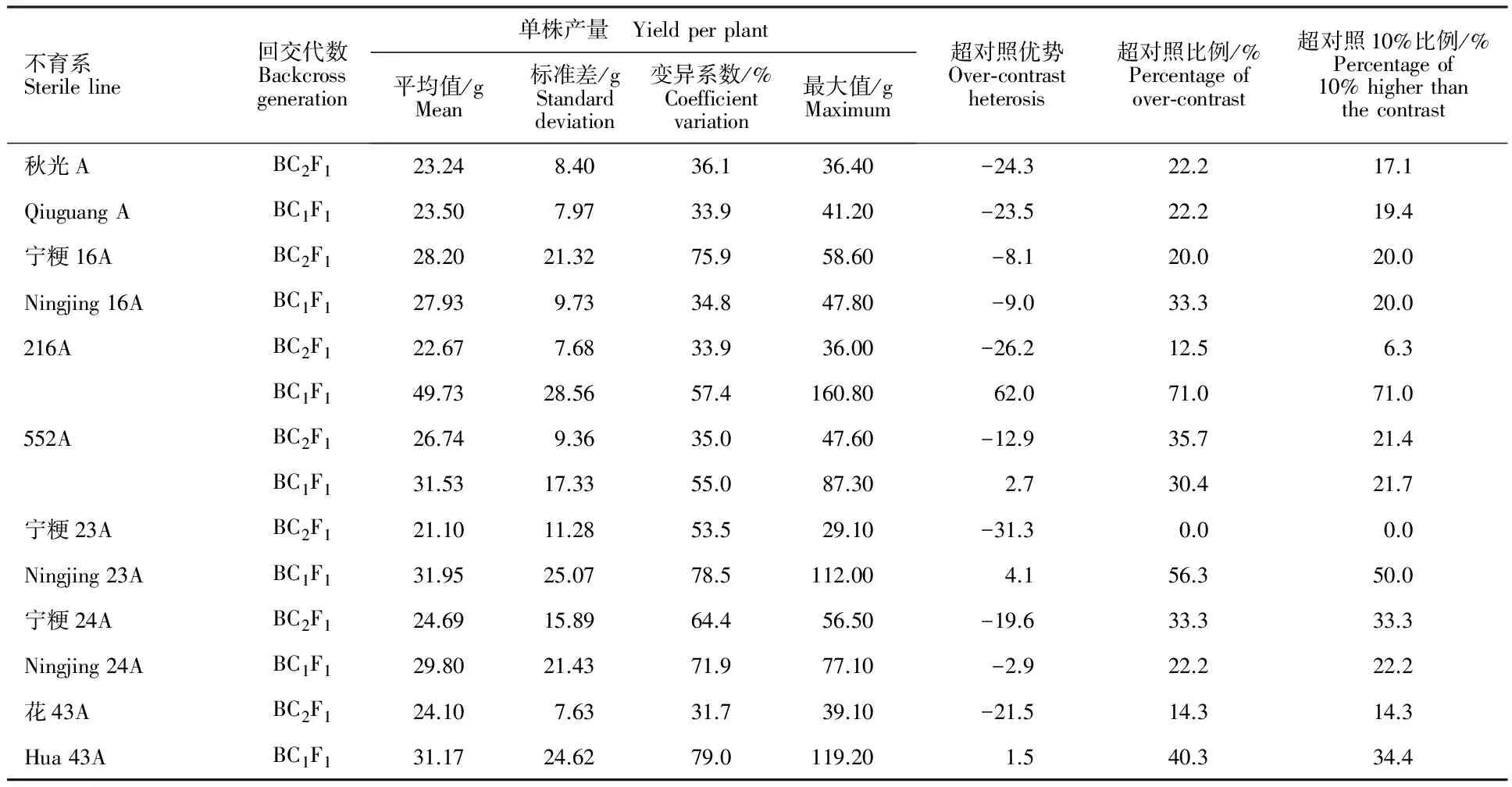
表3 ‘嘉农428’不同回交代数与各不育系杂种优势Table 3 Yield heterosis between ‘Jianong 428’ backcrossed generations and different sterile lines
2.3 不同回交世代与各不育系F1代主要农艺性状
2.3.1 ‘嘉农育251’ 不同回交世代与各不育系F1代主要农艺性状 ‘嘉农育251’回交到BC1F1和BC2F1代与‘秋光A’ ‘宁粳16A’ ‘216A’ ‘552A’ ‘宁粳23A’ ‘宁粳24A’和‘花43A’杂交F1代主要农艺性状见表4。由表4可以看出株高整体变化较小,‘嘉农育251’回交BC2F1和BC1F1代与同一不育系杂交F1代较接近,与‘宁粳23A’杂交F1代相等都为101 cm, 差距最大的为‘秋光A’不到8 cm,F1代株高标准差和变异系数也较为接近,表明株高比较稳定差距较小。‘216A’ ‘552A’ ‘宁粳23A’与BC1F1和BC1F1代杂交F1有效穗数较为接近,‘秋光A’ ‘宁粳16A’ ‘宁粳24A’和‘花43A’与‘嘉农育251’回交BC1F1杂交F1代有效穗数比与BC2F1杂交F1代多。各不育系与‘嘉农育251’回交BC1F1和BC2F1代杂交F1代的千粒质量变化很小在24~26 g,多数与回交BC1F代杂交F1代的千粒质量略小于BC2F1代杂交F1代。穂长与千粒质量的变化相似,各不育系与‘嘉农育251’回交BC1F1和BC2F1代杂交F1代的穂长变化很小,多集中在17~19 cm,多数与回交BC1F1代杂交F1代的略小于BC2F1代。各不育系与‘嘉农育251’回交BC1F1和BC2F1代杂交F1代每穗粒数差距较大,最高184.6,最小值为123.1,标准差和变异系数在各农艺性状中最大,标准差在20以上,变异系数在16%以上。除‘花43A’和‘秋光A’与‘嘉农育251’回交BC1F1每穗粒数略小于BC2F1代杂交F1代,‘宁粳16A’ ‘216A’ ‘552A’ ‘宁粳23A’ ‘宁粳24A’与BC1F1代杂交F1代每穗粒数都大于BC2F1代杂交F1代。BC1F1和BC2F1代与各不育系杂交F1代结实率变化也较大,在52.4%到76.9%,变异系数在20%以上。

表4 ‘嘉农育251’不同回交代数与各不育系F1主要农艺性状Table 4 Main agronomic characters of F1 generation between ‘Jianongyu 251’ backcrossed generations and different sterile lines
2.3.2 ‘嘉农育428’不同回交世代与各不育系F1代主要农艺性状 ‘嘉农428’回交到BC1F1和BC2F1代与‘秋光A’ ‘宁粳16A’ ‘216A’ ‘552A’ ‘宁粳23A’ ‘宁粳24A’和‘花43A’杂交F1代主要农艺性状见表5。由表5可以看出,BC2F1代与‘秋光A’ ‘宁粳16A’ ‘宁粳23A’和‘宁粳24A’杂交F1代株高大于与BC1F1代杂交F1代, ‘216A’ ‘552A’和‘花43A’与BC2F1代杂交F1代株高小于与BC1F1代杂交F1代且差距较小,各不育系与BC1F1代杂交F1代株高的标准差和变异系数都大于与BC2F1代杂交F1代,表明BC1F1代有较大的变异幅度。‘嘉农428’回交到BC1F1和BC2F1代与各不育系有效穗数变化较大,总体看BC1F1代与各不育系杂交F1代有效穗数比BC2F1代与各不育系杂交F1代,其标准差和变异系数都大于与BC2F1代。‘嘉农428’回交到BC1F1和BC2F1代与各不育系杂交F1代千粒质量除‘552A’变化较大,多数BC1F1代与各不育系杂交F1代略大于BC2F1代与各不育系杂交F1代。
表5 ‘嘉农428’不同回交代数与各不育系F1主要农艺性状Table 5 Main agronomic characters of F1 generation between ‘Jianong 428’ backcrossed generations and different sterile lines
不同不育系与‘嘉农428’回交系杂交F1代穂长变化较大,而不同回交代与不育系杂交F1代变化较小。每穗粒数除‘秋光A’和‘宁粳16A’ ,BC1F1代与‘216A’ ‘552A’ ‘宁粳23A’‘宁粳24A’和‘花43A’杂交F1代每穗粒数及其标准差和变异系数大于BC2F1代与各不育系杂交F1代。‘嘉农428’回交到BC1F1和BC2F1代与各不育系杂交F1代的结实率变化较大,不同不育系间杂交F1代变化大,各代间的杂交F1代变化也大。
3 讨 论
对BC1F1代和BC2F1代群体的恢复基因检测和测恢,结果各世代有恢复基因和无恢复基因都符合1∶1的分离,说明在粳型优势生态群遗传背景下,恢复基因符合一对主效基因孟德尔分离规律,与多数研究一致。
回交BC1F1代与各不育系杂交F1代平均单株产量均大于BC2F1代与各不育系杂交F1,其单株产量标准差和变异系数大于BC2F1代。BC1F1代与各不育系杂交F1代超对照优势比BC2F1代超对照优势高,超对照植株比例也高,有效穗数和每穗粒数多;株高、穂长和千粒质量较接近。在乔善宝等[20]的研究中也指出回交 1 次选系优于回交 2 次。
杂种优势是一种受许多因素影响的复杂生物学现象,其遗传基础都是一些假说,较为经典的包括显性、超显性和上位性3个假说。这些假说在许多试验中被验证,如Xiao等[21]、Stuber等[22]、和You等[23]的研究以及与产量相关的一些基因Ghd7[24]、GhdB[25]、Ghd7.l[26]和Hdl[27]都表现为显性效应。Li等[28]、Luo等[29]和Bian等[30]研究发现大部分quantitative trait locus (QTL)表现超显性。Li等[31]和Dan[32]等的研究很好地解释了上位性假说。因此不断的累加优势基因是提高杂种优势的有效办法。
将优势生态型转育成恢复系对回交转育各时代进行测恢,可以看到回交BC1F1代与各不育系杂交F1代平均单株产量绝大多数大于BC2F1代与各不育系杂交F1,超对照优势高,超对照植株比例大,有效穗数多,每穗粒数多。回交BC1F1代与各不育系杂交F1代在单株产量以及其他多个性状表现出较强的优势。说明在BC1F1代群体中带有更多的优势基因,这可能是由于回交的供体材料‘C418’为北方杂交稻骨干恢复系,具有非常好的配合力,随着回交代数的增加,‘C418’基因比例的减少,造成更多的优势基因减少。为保留更多的优势基因在回交转育过程中回交代数不宜太多,在回交一两代后进行自交选育可以保留更多的基因型,有利选育出配合力高的株系。
Reference:
[1] 袁 勤,倪林娟,曹黎明,等.杂交粳稻的选育与应用 [J].上海农业学报,2002,18(1) :25-28.
YUAN Q,NI L J,CAO L M,etal.Breeding selection and application of hybrid japonica rice [J].ActaAgriculturaeShanghai,2002,18(1) :25-28.
[2] 邓华凤,何 强,舒 服,等.中国杂交粳稻研究现状与对策 [J].杂交水稻,2006,21(1):1-6.
DENG H F,HE Q,SHU F,etal.Status and technical strategy on development of japonica hybrid rice in China [J].HybridRice,2006,21(1):1-6.
[3] 邱福林,庄杰云,华泽田,等.北方杂交粳稻骨干亲本遗传差异的SSR标记检测 [J].中国水稻科学,2005,19(2):101-104.
QIU F L,ZHUANG J Y,HUA Z T,etal.Inspect of genetic differentiation of main parents of japonica hybrid rice in the Northern China by simple sequence repeats(SSR) [J].ChineseJournalofRiceScience,2005,19(2):101-104.
[4] 陈 灿,员海燕,雷云天.玉米改良群体MM中选系的产量配合力及杂种优势分析 [J].西北农林科技大学学报(自然科学版),2013,41(3):93-98.
CHEN C,YUN H Y,LEI Y T.Analysis of yield combining ability and heterosis of selective lines from the improved maize population MM [J].JournalofNorthwestA&FUniversity(NaturalScienceEdition),2013,41(3):93-98.
[5] 吴敏生,王守才,戴景瑞.AFLP 分子标记在玉米优良自交系优势群划分中的应用 [J] .作物学报,2000,26(1):9-13.
WU M SH,WANG SH C,DAI J R,etal.Application of AFLP marker to heterotic grouping of elite maize inbred lines [J].ActaAgronomicaSinica,2000,26(1):9-13.
[6] 袁力行,傅骏骅,张世煌,等.利用RFLP和SSR标记划分玉米自交系杂种优势群的研究[J].作物学报,2001,27(2):149-156.
YUAN L X,FU J H,ZHANG SH H,etal.Heterotic grouping of maize inbred lines using RFLP and SSR Markers [J].ActaAgronomicaSinica,2001,27(2):149-156.
[7] 黄益勤,李建生.利用RFLP标记划分45份玉米自交系杂种优势群的研究 [J] .中国农业科学,2001,34(3):244-250.
HUANG Y Q,LI J SH.Classification of hererotic groups with RFLPS among 45 maize inbred lines [J].ScientiaAgriculturaSinica,2001,34(3):244-250.
[8 ] 赵新亮,郭霭光.利用SRAP分子标记划分玉米自交系类群初探[J].西北农业学报,2007,16(3):77-81.
ZHAO X L,GUO A G.Preliminary study on the classification of maize inbred lines by SRAP marker technology.[J].ActaAgriculturaeBoreali-occidentalisSinica,2007,16(3):77-81.
[9] 黎 裕,王天宇.我国玉米种质基础与骨干亲本的形成 [J].玉米科学,2010,18(5):1-8.
LI Y,WANT T Y.Germplasm base of maize breeding in China and formation of foundation parents [J].JournalofMaizeSciences,2010,18(5):1-8.
[10] 刘志斋,吴 迅,刘海利,等.基于40个核心SSR标记揭示的820份中国玉米重要自交系的遗传多样性与群体结构 [J].中国农业科学2012,45(11):2107-2138.
LIU ZH ZH ,WU X,LIU H L,etal.Genetic diversity and population structure of important Chinese maize inbred lines revealed by 40 core simple sequence repeats(SSRs) [J].ScientiaAgriculturaSinica,2012,45(11):2107-2138.
[11] 刘正理.谷子杂种优势群的构建方法及研究进展 [J].河北农业科学,2010,14(11):102-104.
LIU ZH L.Establish method of heterotic group in foxtail millet and its research progress [J].JournalofHebeiAgriculturalSciences,2010,14(11):102-104.
[12] 逯腊虎,李振兴,倪中福,等.小麦杂种优势群研究Ⅵ.普通小麦与穗分枝小麦、轮回选择后代材料、西藏半野生小麦和斯卑尔脱小麦早熟诱变系的SSR分子标记遗传差异研究 [J].麦类作物学报,2007,27(2):201-206.
LU L H,LI ZH X,NI ZH F,etal.Study on wheat heterotic groupⅥ.genetic diversity revealed by SSR marker between common wheat,ear-branched wheat,wheat lines derived from recurrent selection,tibetan wheat and early spelt wheat mutant lines [J].JournalofTriticeaeCrops,2007,27(2):201 -206.
[13] 史秀秀,毕晓静,马守才,等.黄淮麦区杂交小麦亲本的杂种优势和配合力分析 [J].麦类作物学报,2013,33(6):1111-1118.
SHI X X,BI X J,MA SH C,etal.Combining ability and heterosis in hybrid wheat of parents from Huang-huai wheat production area [J] .JournalofTriticeacCrops,2013,33(6):1111-1118.
[14] 邹小云,宋来强,陈伦林,等.利用SRAP标记划分甘蓝型油菜杂种优势群 [J].江西农业学报,2011,23(4):1-4.
ZHOU X Y,SONG L Q,CHEN L L,etal.Classification of heterosis groups for rapeseed(Brassicanapus) by SRAP markers [J].ActaAgriculturaeJiangxi,2011,23(4):1-4.
[15] 刘 炜,李自超,史延丽,等.试用配合力进行粳型水稻杂种优势生态型的划分 [J].作物学报,2004,30(1):66-72.
LIU W,LI Z CH,SHI Y L,etal.Heterotic ecotypes grouping of japonica rice by combining ability [J].ActaAgronomicaSinica,2004,30(1):66-72.
[16] 刘 炜,史延丽,马洪文,等.根据杂种优势值划分粳型水稻杂种优势生态型 [J].西北植物学报,2005,25(1):64-69.
LIU W,SHI Y L,MA H W,etal.Study on heterotic ecotypes of japonica rice based on the heterosis [J].ActaBotanicaBoreali-OccidentaliaSinica,2005,25(1):64-69.
[17] 刘 炜,史延丽,李自超,等.粳型水稻杂种优势生态型与杂种优势模式的研究 [J].西北农林科技大学学报(自然科学版),2005,33(1):108-114.
LIU W,SHI Y L,LI Z CH,etal.Studies on heterotic ecotypes and heterotic patter s of japonica rice [J].JournalofNorthwestA&FUniversity(NaturalScienceEdition),2005,33(1):108-114.
[18] 熊尧宇,李芦江,文水清,等.玉米自交系08-641不同选择方向回交改良后代主要性状的配合力[J].玉米科学,2012,20(1):10-14.
XIONG Y Y,LI L J,WEN SH Q.etal.Combining ability of the backcross modified line developed from 08-641 selected by different directions[J].JournalofMaizeSciences,2012,20(1):10-14.
[19] 王 坚,刘 炜.杂交粳稻BT型雄性不育恢复基因功能标记优化研究 [J].宁夏农林科技,2015,56(10):29-33.
WANG J,LIU W.A Study of optimization of functional markers for restorer gene of BT-type cytoplasmic male sterility [J].NingxiaJournalofAgricultureandForestryScienceandTechnology,2015,56(10):29-33.
[20] 乔善宝,王玉花,杨克诚,等.不同供体及不同回交次数对玉米自交系 R08 的改良效应[J].作物学报,2009,35(12):2187-2196
QIAO SH B,WANG Y H,YANG K CH.etal.Effects contributed by different donor parents and backcross times on R08 improvement[J].ActaAgronomicaSinica,2009,35(12):2187-2196.
[21] XIAO J,LI J,YUAN L,etal.Dominance is the major genetic basis of heterosis in rice as revealed by QTL analysis using molecular markers [J].Genetics,1995,140(2):745-754.
[22] STUBER C W,LINCOLN S E,WOLFF D W,etal.Identification of genetic factors contributing to heterosis in a hybrid from two elite maize inbred lines using molecular markers[J].Genetics,1992,132(3):823-839.
[23] YOU A Q,LU X G, JIN H J,etal.Identification of quantitative trait loci across recombinant inbred lines and testcross populations for traits of agronomic importance in rice[J].Genetics,2006,172(2):1287-1300.
[24] XUE W Y,XING Y Z,WENG X Y,etal.Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice[J].NatureGenetice,2008,40(6):761-767.
[25] YAN W H,WANG P,CHEN H X,etal.A major QTL, Ghd8 , plays pleiotropic roles in regulating grain productivity,plant height,and heading date in rice[J].MolecularPlant,2011,4(2):319-330.
[26] YAN W H,LIU H Y,ZHOU X C,etal.Natural variation in Ghd7.1 plays an important role in grain yield and adaptation in rice[J].CellResearch,2013,23(7):969-971.
[27] GARCIA A A F,WANG S C,MELCHINGER A E,etal.Quantitative trait loci mapping and the genetic basis of heterosis in maize and rice[J].Genetics,2008,180(3):1707-1724.
[28] LI Z K,LUO L J,MEI H W,etal.Overdominant epistatic loci are the primary genetic basis of inbreeding depression and heterosis in rice.I.Biomass and grain yield[J].Genetics,2001,158(4):1737-1753.
[29] LUO X J ,FU Y C,ZHANG P J,etal.Additive and over-dominant effects resulting from epistatic loci are the primary genetic basis of heterosis in rice [J].JournalofIntegrativePlantBiology,2009,51(4):393-408.
[30] BIAN J M,JIANG L,LIU L L,etal.Identification of japonica chromosome segments associated with heterosis for yield in indica × japonica rice hybrids[J].CropScience,2010,50(6):2328-2337.
[31] LI Z,PINSON S R M,PARK W D,etal.Epistasis for three grain yield components in rice(OryzasativaL.) [J].Genetics,1997,145(2):453-465.
[32] DAN Z W,HU J,ZHOU W,etal.Hierarchical additive effects on heterosis in rice(OryzasativaL.) [J].FrontiersinPlantScience,2015,6:738.
