豚鹿几种激素的季节性变化模式研究
2018-01-30严慧娟王巍易军牛李丽屈羽陈昂蒲阳邓家波钟燕余建秋
严慧娟, 王巍, 易军, 牛李丽, 屈羽, 陈昂, 蒲阳, 邓家波, 钟燕, 余建秋*
(1.成都动物园,成都野生动物研究所,成都610081;2. 四川省畜牧科学研究院,动物遗传育种四川省重点实验室,成都610066)
豚鹿Axisporcinus隶属于哺乳纲Mammalia偶蹄目Artiodactyla鹿科Cervidae斑鹿属Axis,中国仅有印支亚种A.porcinusannamiticus,主要分布于云南西部靠近中缅边境的耿马、西盟两县的南丁河沿岸,1975年被《濒危野生动植物种国际贸易公约(CITES)》列入附录Ⅰ。近20年来,全球野生豚鹿数量急剧减少,世界自然保护联盟(IUCN)在2008年将其收录到受威胁物种红色名录并列为濒危(EN)等级。在中国,豚鹿被列为国家Ⅰ级重点保护野生动物,是我国数量最稀少的鹿类之一。目前,我国豚鹿野生种群基本灭绝(汪松,1998),还保存有国家动物园系统的人工饲养群体,数量仅40只左右,已成为濒危小种群。豚鹿的保种及扩繁一直受基础群体数量有限、繁殖活动规律性不强和近亲交配等因素制约。通过调查成都动物园豚鹿群体繁殖成活率,发现豚鹿的初生死亡率高达60%,主要因为其繁殖行为规律不明确,无法进行相应的机体调节和环境控制措施,导致初生幼崽发生环境应激性死亡(邓家波等,2011)。因此,明确各季节豚鹿体内繁殖相关激素变化模式,确定其繁殖行为规律,对于提高繁殖成活率至关重要。
动物的繁殖行为涉及下丘脑-垂体-性腺轴系统复杂的神经内分泌过程(何世山,金小军,2003)。光周期循环和温度等一系列季节性节律变化因素能够转换为生理信号,刺激kisspeptin神经元(Silveiraetal.,2010),作用于下丘脑的GnRH神经元,改变GnRH和促性腺激素的分泌模式,调节生殖行为(Malpauxetal.,2002;Smithetal.,2008)。对反刍动物的研究发现,直接参与繁殖的激素主要包括促黄体素(LH)、外周血液促卵泡素(FSH)和催乳素(PRL),间接参与繁殖行为的激素为三碘甲腺原氨酸(T3)、四碘甲腺原氨酸(T4)(赖平等,2012;狄冉等,2015)。本研究通过监测2015年5月、8月和10月雌性豚鹿血液中繁殖相关激素含量的变化,结合温度、湿度和光照的变化,粗略绘制春季、夏季和秋季豚鹿季节性内分泌调节趋势图。揭示圈养条件下季节变化与豚鹿繁殖的关系,明确豚鹿繁殖活动规律,以便根据季节调整养殖模式,有效提高豚鹿繁殖成活率,逐步扩繁种群。
1 材料与方法
1.1 试验群体组建
在成都动物园的豚鹿群体中选择16月龄左右的8头雌鹿作为实验动物,固定实验群体。
1.2 温湿指数及光照时间测定
在鹿舍中部距地面1.5 m处挂置干湿球温度计,于2015年5—12月,记录每日09∶00、12∶00、20∶00的温度和湿度。按照Sandy(2000)提出的温湿度指数(temperature-humidity index,THI):THI=0.8×温度+相对湿度×(温度-14.4)+46.4计算;根据国际上THI值划分热应激标准:THI<72,无热应激;THI=72~79,温和应激;THI=80~89,中等应激;THI=90~98,严重应激;THI>98,致死应激。同时记录每天昼长时间。
1.3 血液样本采集和生化指标测定
实验期间,当连续3 d的THI值差异无统计学意义(P>0.05)时,于12∶00—14∶00采集8头豚鹿的外周静脉血,3 000 r·min-1离心10 min将血清和红细胞迅速分离,分装,于-20 ℃冻存。用T3、T4、LH、FSH和PRL的R & D上海卡麦舒ELISA试剂盒检测。
取微孔平板,加入样本10 μL,添加稀释液40 μL,再加入HRP标记抗体100 μL,于37 ℃封闭水浴60 min。反复洗涤5次,加入底物各50 μL于37 ℃避光孵育15 min,加入50 μL终止液,于15 min内在450 nm波长处测定光密度值。
1.4 数据分析
实验数据采用SPSS 17.0进行K-S检验,显著性水平为α=0.05。利用R语言对差异性状及群体进行层次聚类分析。
2 实验结果
2.1 豚鹿圈舍THI值和光照趋势
8月豚鹿圈舍内THI值为72~80,为温和应激。其余月份的THI值均在72以下,无热应激。昼长时间变化趋势分析发现,6—8月的光照时间较长,日均达到13.63 h以上。10—12月的光照时间较短,日均为10.66 h(图1)。
2.2 豚鹿激素水平季节性变化趋势
分析发现,8月豚鹿血清中T3、T4的含量极显著低于5月、10月(P<0.01)。LH、PRL的含量在8月均极显著下降(P<0.01),在10月极显著回升(P<0.01)。FSH在8月的含量极显著低于5月(P<0.01),但与10月的差异无统计学意义(P>0.05)(图2)。
2.3 各激素指标间相关性分析
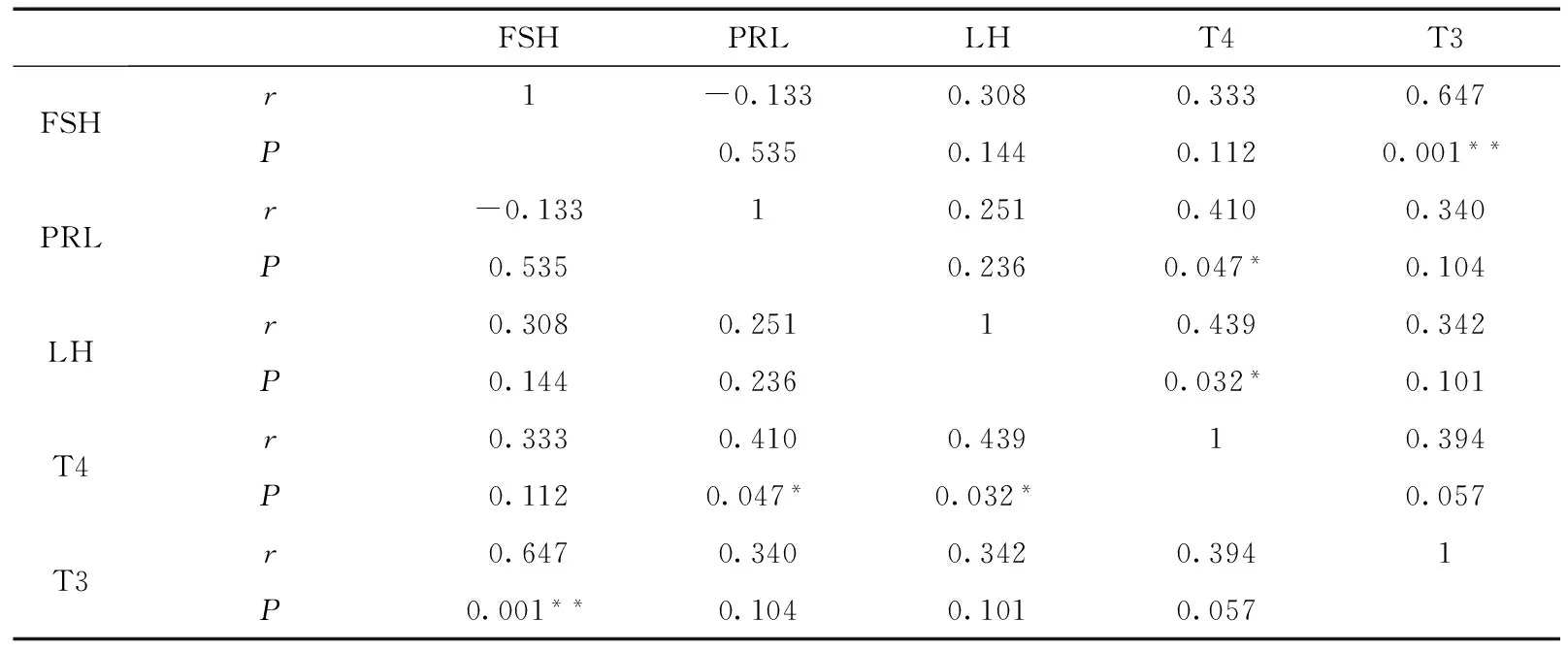
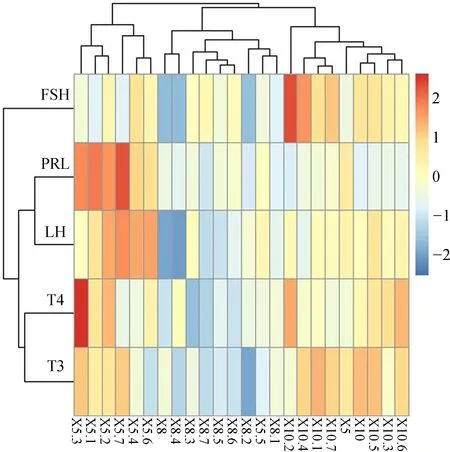
血清中LH、FSH、PRL、T3、T4含量相关性分析显示(表1),FSH与T3呈极显著正相关(P<0.01,r=0.647),PRL与T4呈显著正相关(0.01 图1 豚鹿圈舍温湿度指数和昼长时间 图2 豚鹿血清中激素含量变化趋势Fig. 2 The variation of hormone content in serum of Axis porcinus **P<0.01,*P<0.05; 下同the same below 表1 豚鹿血清中激素含量相关性分析(n=24)Table 1 Correlation analysis of hormone content in serum of Axis porcinus (n=24) 将5月、8月和10月各样本中LH、FSH、PRL、T3、T4活性情况进行层次聚类分析,结果发现,T3、T4聚为一类,LH、PRL聚为一类,并且与FSH为二级聚类(图3)。表明T3、T4活性的变化模式相似度较高,LH、PRL活性的变化模式相似度较高,但FSH的活性变化模式与其余4种激素有差异。 图3 豚鹿血清各激素聚类图Fig. 3 Cluster of hormones in serum of Axis porcinus T3、T4在血清中的含量变化与热应激的时间和应激程度存在密切联系(Yokusetal.,2006)。研究发现,T3、T4在各样本中的变化模式相同;8月圈舍THI值达72~80、日照时间达13.63 h以上时,豚鹿血清中T3、T4的含量极显著低于5月、10月,与同为反刍动物的奶牛在湿热胁迫下的表现一致(Collieretal.,1981;魏学良等,2005;樊华等,2007)。对马鹿Cervuselaphus的研究亦发现,T3、T4的分泌存在季节性变化(施振旦,1995)。推测较高的温湿度和长日照所形成的胁迫造成了机体应激,使血浆中的T3、T4含量均显著降低,从而调节自身体温。大量研究表明,T3、T4含量的改变并不引起褐黑激素的变化,而通过影响下丘脑(Websteretal.,1991),调节kisspeptin神经系统和GnIH神经系统对动物的季节性繁殖起调节作用(Bernal,2002;Yasuoetal.,2006)。本研究通过激素间的相关性分析,证明了T3、T4的活性与繁殖相关激素具有相关关系。因此,推测高温、高湿和长日照的胁迫可能引起血清中T3、T4含量的变化,间接影响豚鹿的繁殖表现。 LH是由垂体前叶促黄体素细胞嗜碱性细胞产生的一种糖蛋白激素,能够作用于成熟的卵泡并促发排卵(Wuetal.,2000)。研究发现,在繁殖期不同阶段,丘脑GnRH分泌的脉冲频率不同。繁殖期开始时,LH在排卵期前达到最大值,在繁殖末期降至最低,随后卵巢进入静止期(Clarke,1993;王训立等,2002)。FSH是由垂体前叶嗜碱性细胞分泌的一种糖蛋白激素,能够在繁殖期促使卵泡内膜细胞分泌雌激素,激发卵泡的最后成熟,诱发排卵并使颗粒细胞变成黄体细胞(解美娜等,2004)。对滩羊Procapraprzewalskii的研究发现,FSH和LH含量变化具有显著的季节性,且秋冬高于春夏(祁云霞等,2014)。另有研究发现,随春季光照延长,哺乳动物达到FSH分泌上升的光照阈值,随后LH也随着光照的进一步延长而增加(杨海明,2014)。PRL作为一种由垂体前叶腺嗜酸细胞分泌的蛋白质激素,可以促进乳腺生长发育,刺激并维持泌乳(Bole-Feysotetal.,1998)。研究发现,随光照和温度的变化,马岗鹅Bostaurus血浆中PRL的浓度呈周期性变化(黄运茂等,2007)。本研究发现,8月THI值较高时,豚鹿血清中的LH、PRL含量极显著低于5月、10月,FSH含量极显著低于5月,表明豚鹿繁殖相关激素的分泌与光照时长和THI值相关,季节变化能够在一定程度上影响豚鹿的繁殖效率。豚鹿繁殖激素的变化模式符合短光照繁殖类型动物随光照延长、繁殖激素表达降低的基本特征,推断豚鹿属于短光照繁殖类型动物。光照延长后,豚鹿GnRH活化的同时也激发雌激素的负反馈作用,使机体进入非繁殖期(Knightetal.,1983;贺建宁等,2013),另外,夏季的高温高湿胁迫也是限制豚鹿繁殖表现的重要因素。 本研究发现,豚鹿属于短光照繁殖类型动物,夏季的高温、高湿和长光照会影响豚鹿繁殖相关激素的分泌。由于圈养环境外在遮挡物较少,豚鹿直接受到强光和高湿热胁迫而产生应激,建议在饲料中添加抗应激类添加剂,或改善圈舍条件,如增加遮光设施、增加植被和喷淋设施降低温度等,以便缓解应激对机体生殖激素的逆向调节,在一定程度上提高其繁殖效率。 邓家波, 余建秋, 牛李丽, 等. 2011. 豚鹿繁殖成活率低的病因学调查研究[J]. 野生动物, 32(3): 120-122. 狄冉, 郭晓飞, 刘秋月, 等.2015.生殖激素对绵羊繁殖性能影响的研究进展[J]. 家畜生态学报, 36(9): 1-6. 樊华, 樊丽, 王峰, 等. 2007. 热应激对泌乳奶牛血液生化指标的影响[J]. 中国畜牧兽医, 34(4): 45-46. 何世山, 金小军. 2003. 高温对肉鸡血液生化指标的影响[J]. 浙江大学学报(农业与生命科学版), 29(3): 311-314. 贺建宁, 王金鑫, 狄冉, 等. 2013. 常年发情和季节性发情绵羊在不同季节生殖激素变化规律[J]. 畜牧兽医学报, 44(10): 1547-1553. 黄运茂, 施振旦, 李孝伟, 等. 2007. 光周期对马岗鹅产蛋、PRL和LH分泌季节性变化的影响[J]. 华南农业大学学报, 28(3): 94-96. 赖平, 王凭青, 张宝云, 等. 2012.哺乳动物季节性繁殖的神经内分泌调节机制[J]. 遗传, 34(3): 281-288. 祁云霞, 何小龙, 刘晓芳, 等. 2014. 巴美肉羊发情期血清FSH和LH浓度变化规律及其与产羔数关系分析[J]. 黑龙江畜牧兽医, (7): 17-20. 施振旦. 1995. 甲状腺功能对调节雄性马鹿体重和繁殖活动季节性变化的研究[J]. 兽类学报, 15(1): 17-24. 汪松. 1998. 中国濒危动物红皮书[M]. 北京: 科学出版社. 王训立, 周建华, 范春梅. 2002. 季节性变化对雌性恒河猴生殖功能的影响[J]. 中国实验动物学报, 10(1): 21-26. 魏学良, 张家骅, 王豪举, 等. 2005. 高温环境对奶牛生理活动及生产性能的影响[J]. 中国农学通报, 21(5): 13-15. 解美娜, 张才乔, 米玉玲, 等. 2004. 卵泡刺激素和雌激素对培养的鸡胚卵巢生殖细胞增殖的影响[J]. 动物学研究, 25(1): 53-56. 杨海明. 2014. 扬州鹅繁殖的光照调控及下丘脑基因差异表达研究[D]. 江苏扬州: 扬州大学. Bernal J. 2002. Action of thyroid hormone in brain[J]. Journal of Endocrinological Investigation, 25(3): 268-288. Bole-Feysot C, Goffin V, Edery M,etal. 1998. Prolactin and its receptor: actives, signal transduction pathways and phenotypes observed in PRL receptor knockout mice[J]. Endocrine Reviews, 19(3): 225-268. Clarke IJ. 1993. Variable patterns of gonadotropin-releasing hormone secretion during the estrogen-induced luteinizing hormone surge inovariectomized ewes[J]. Endocrinology, 133(4): 1624. Collier RJ, Eley RM, Sharma AK,etal. 1981. Shade management in subtropical environment for milk yield and composition in holstein and jersey cows 1[J]. Journal of Dairy Science, 64(5): 844-849. Knight PG, Cunningham FJ, Gladwell RT. 1983. Concentrations of immunoreactive luteinizing hormone releasing hormone in discrete brain regions of the cockerel: effects of castration and testosterone replacement therapy[J]. Journal of Endocrinology, 96(3): 471. Malpaux B, Tricoire H, Mailliet F,etal. 2002. Melatonin and seasonal reproduction: understanding the neuroendocrine mechanisms using the sheep as a model[J]. Reproduction Supplement, 59(1): 167-179. Sandy S. 2000. Is your dairy management program ready for the summer heat[D]. Texas: The Texas A & M University System. Silveira LFG, Teles MG, Trarbach EB,etal. 2010. Role of kisspeptin/GPR54 system in human reproductive axis[J]. Frontiers of Hormone Research, 39: 13-24. Smith JT, Coolen LM, Kriegsfeld LJ,etal. 2008. Variation in kisspeptin and R Famide-related peptide (RFRP) expression and terminal connections to gonadotropin-releasing hormone neurons in the brain: a novel medium for seasonal breeding in the sheep[J]. Endocrinology, 149(11): 5770-5782. Webster JR, Moenter SM, Woodfill CJI,etal. 1991. Role of the thyroid gland in seasonal reproduction: Ⅱ. Thyroxine allows a season-specific suppression of gonadotropin secretion in sheep[J]. Endocrinology, 129: 176-183. Wu J, Nayudu PL, Kiesel PS,etal. 2000. Luteinizing hormone has a stage-limited effect on preantral follicle development in vitro[J]. Biology of Reproduction, 63(1): 320-327. Yasuo S, Nakao N, Ohkura S,etal. 2006. Long-day suppressed expression of type 2 deiodinase gene in the mediobasal hypothalamus of the Saanen goat, a short-day breeder: implication for seasonal window of thyroid hormone action on reproductive neuroendocrine axis[J]. Endocrinology, 147(1): 432-440. Yokus B, Cakir DU, Kanay Z,etal. 2006. Effects of seasonal and physiological variations on the serum chemistry, vitamins and thyroid hormone concentrations in sheep[J]. Journal of Veterinary Medicine, 53(6): 271-276.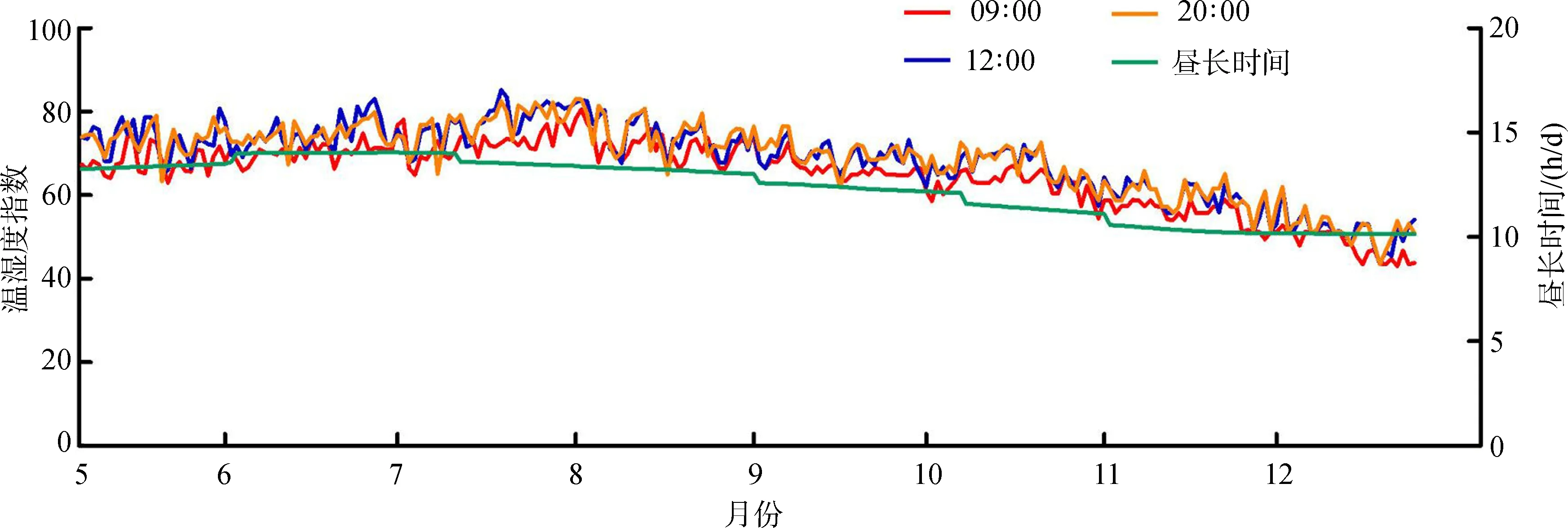
Fig. 1 The trend of temperature-humidity index in breeding houses ofAxisporcinusand daylength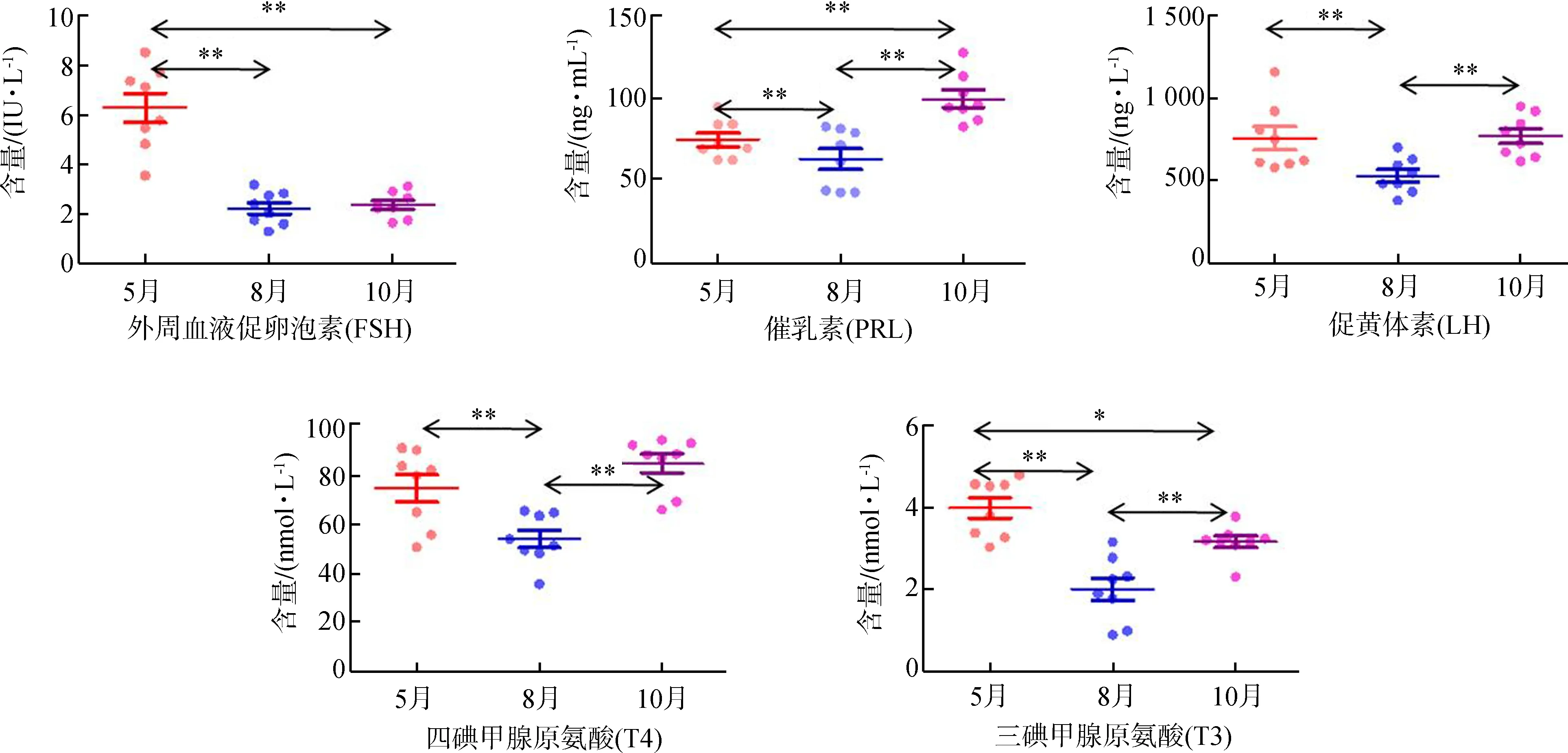
2.4 各激素指标间层次聚类分析
3 讨论
