A new alvarezsaurid dinosaur from the Late Cretaceous Qiupa Formation of Luanchuan, Henan Province, central China
2018-01-18JunhangLiXuHualiChangSonghaiJiaJimingZhangDiansongGaoYiyangZhangChengjunZhangFangDing
Jun-hang Lü, Li Xu, Hua-li Chang, Song-hai Jia, Ji-ming Zhang, Dian-song Gao, Yi-yang Zhang,Cheng-jun Zhang, Fang Ding
a Institute of Geology, Chinese Academy of Geological Sciences, Beijing 100037, China
b Henan Geological Museum, Zhengzhou 450016, China
c College of Earth Sciences &Key Laboratory of Mineral Resources of Gansu Province, Lanzhou University, Lanzhou 730000, China
A B S T R A C T
An alvarezsaurid dinosaur skeleton was discovered from the Late Cretaceous Qiupa Formation of Luanchuan, Henan Province of centtral China. It represents a new alvarezsaurid dinosaur Qiupanykus zhangi gen. et sp. nov. A phylogenetic analysis recovers Qiupanykus nested within the unresolved clade, which includes Asian and north American taxa. The skeleton of the new specimen is preserved in association with eggshells. The eggshell morphologies show that these eggs belong to oviraptorid eggs, skeletal remains of which were discovered from the same area. The alvarezsaurid skeleton associated with eggshell fragments may indicate that these eggs were broken by the strong thumb-claws of the former and that alvarezsaurid dinosaurs may be egg-eaters.
Keywords:
Vertebrate paleontology
Alvarezsaurid dinosaur
Qiupanykus
Late Cretaceous
Central China
1. Introduction
Alvarezsaurid dinosaurs, a highly specialized non-avian theropod group, are characterized by being small, long-legged running animals once regarded to be a group of flightless birds (Perle A et al., 1993, 1994; Chiappe LM et al., 1996,1998). The first described alvarezsaurid dinosaurs are Mononykus and Shuvuuia in the 1990’s, which were discovered from the Late Cretaceous of Mongolia. Now they are known from the early Late Jurassic and Late Cretaceous of China, and the Early and Late Cretaceous of North and South America as well.
The genera from Eurasia and North America are Mononykus, Shuvuuia, Parvicursor and Albinykus (Perle A et al., 1993; Karhu AA and Rautian AS, 1996; Chiappe LM et al., 1998; Suzuki SL et al., 2002; Hutchinson J and Chiappe L, 1998; Naish D and Dyke GD, 2004; Alifanov VR and Barsbold R, 2009; Choiniere JN,2010; Xu X et al., 2010,2 011; Nesbitt SJ et al., 2011; Averianov A and Sues HD,2017) and those from South America are Patagonykus (Novas FE, 1996, 1997), Alvarezsaurus (Bonaparte JF, 1991),Achillesaurus (Martinelli AG and Vera EI, 2007) and Bonapartenykus (Agnolin FL et al., 2012).
An alvarezsaurid dinosaur from Luanchuan, Henan Province was briefly reported in 2012 (Lü JC et al., 2012),and the one associated with egg fragments was also briefly reported in 2017 (Kundrat M et al., 2017).
Herein described is a new alvarezsaurid dinosaur discovered from the uppermost Cretaceous deposits of the Qiupa Formation, Tantou Basin, Henan Province of central China(Fig.1). It represents the youngest alvarezsaurid dinosaur from China so far.
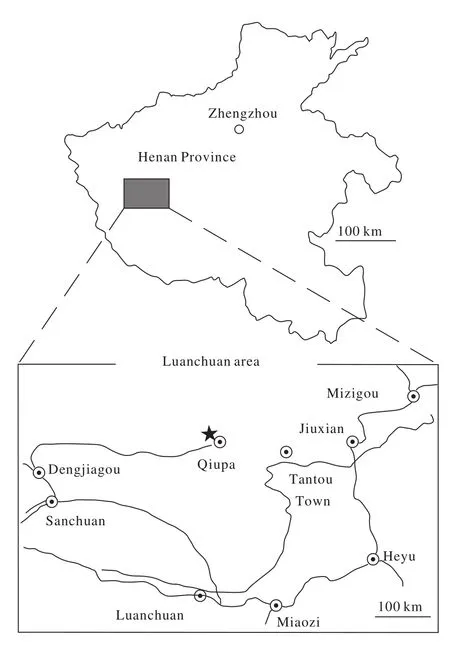
Fig. 1. Map of the holotype locality in Luanchuan of Henan Province, China. The star represents the fossil site.
2. Geological setting
The specimens were discovered from the Qiupa Formation (Late Maastrichtian) (Bureau of Geology and Mineral Resources of Henan Province, 1989), which is dominated by brownish red thick-bedded siltstone, calcareous mudstone, interbedded with thin fine conglomerates, and parallel and cross laminations which indicate shallow lacustrine and braided river delta facies (Jiang XJ et al., 2011).
3. Systematic paleontology
Maniraptora (Gauthier, 1986)
Alvarezsauridae (Bonaparte JF, 1991)
Qiupanykus zhangi gen. et sp. nov.
Etymology: The generic name refers to the Qiupa town,Luanchuan County, where the specimen was discovered. The specific name is in honor of Shuancheng Zhang for his logistic support with fossil searching and excavations in the field.
Holotype: Incomplete skeleton comprising most posterior axial elements and most of hindlimb elements. The specimen 41HIII-0101 is housed in the collections of the Henan Geological Museum, Zhengzhou, China.
Holotype locality and horizon: The specimen (41HIII-0101) was found in Guanping, Qiupa Town, Luanchuan County of Henan Province; Qiupa Formation (Bureau of Geology and Mineral Resources of Henan Province, 1989; Lü JC et al., 2007)
Diagnosis: A small-sized alvarezsaurid dinosaur with the following combination of characters: posterior sacral vertebrae bearing a strong ventral keel; proximal caudals with transverse processes centrally positioned on centrum; pubic articular surface of the pubic peduncle of ilium is reduced and knob-like; fibular crest of tibia large and quadrangular; functional sacrum made up of eight vertebral elements (two anterior caudal plus six sacral vertebrae); a small pneumatic foramen is present in caudal vertebrae.
Description:
The skeleton is not well-preserved, but includes four isolated cervical vertebrae (Fig. 2, Table 1). Three of cervical vertebrae are naturally articulated. The neural spine is weak,short and centered on neural arch, giving the arch an “X”shape in dorsal view. The posterior articular end is wider than the proximal end, but the morphologies of both ends are not clear. The lateral surface of the centrum is slightly concave.The epipophyses of cervical vertebrae are small and placed distally on postzygapophyses, above the postzygopophyseal facets. The anterior extent of the cervical centra are level with or shorter than posterior extent of neural arch. There is a carotid process on posterior cervical vertebrae. There is no pneumatic opening on the cervical vertebra. The ventral surfaces of the cervical vertebrae are narrow. A shallow ventral groove appears near the posterior articular end.
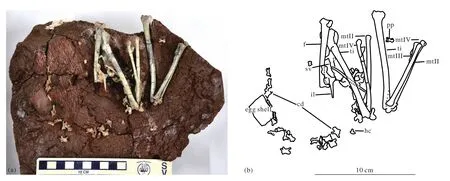
Fig. 2. The photograph (a) and outline drawings (b) of Qiupanykus zhangi: Abbreviations: cd: caudal vertebrae; f: femur; hc: haemal arch; il:ilium; mt(II-IV): metatarsls II-IV; ti: tibia; p: pubis; pp: pedal phalanx; sv: sacral vertebrae.

Table 1. Measurements of Qiupanykus zhangi (41HIII-0101).
No dorsal vertebrae are preserved. Six sacral vertebrae are preserved. The neural canal of the first sacral vertebra is large compared with the centrum. The centra are strongly compressed laterally and form weak ventral keels in the posterior sacrals. There are no pleurocoels on the centra.
There are 25 caudal vertebrae preserved. The neural arch is completely fused with the centrum without any sign of suture in the caudals, which indicates that this animal reached adult stage. The anterior two caudal vertebral centra are strongly appressed into the sacral vertebral column that consists of six vertebrae. It appears thatQiupanykus zhangihad a functional sacrum made up of eight vertebral elements. Most distal caudal vertebrae are missing. In posterior view, the articular ends are ball-shaped in anterior caudal vertebrae, and the neural canal is oval and large. The height of the centrum exceeds its width. The anterior articular end is concave. The prezygapophyses extend beyond the anterior margin of the anterior articular ends. A deep ventral groove is present on the centrum. The postzygapophyses end near the level of the margin of the posterior articular end. The anterior caudal vertebrae do not bear small pneumatic foramen, but the posterior preserved caudal vertebrae bear small pneumatic foramina situated inside shallow depressions on the lateral surface of the centra. The distal caudal vertebrae bear a weak ridge instead of neural spines and their anterior articular ends are rectangular with deep concavities and the posterior articular ends are slightly convex.
One haemal arch is preserved. Its distal end expands much more anteroposteriorly.
A portion of the right ilium and ischium are preserved.The anterior process of the ilium is missing. There is a large and horizontally projected antitrochanter near the weak developed ischial pedicale, which is similar to that ofMononykusolecranus(Perle A et al., 1994). There is a knoblike anterior pubic tubercle below the pubic-iliac contact on the ilium. In ventral view, the acetabulum is twisted. The ventral surface of the postacetabular process is smooth and convex, thus there is no brevis fossa.
The pubes are not well preserved. They are rod-like,slender. There is no pubic apron. Their distal ends are not preserved, thus it is not clear whether there is a pubic boot or not.
The right hindlimb is nearly naturally articulated, only missing its pes. The femur is strongly curved cranially, and the curve apex is approximately at the end of the proximal third of the bone. The femur is slightly sigma-shaped in posterior view, as is the case inParvicursor(Karhu AA and Rautian AS, 1996). The lesser trochanter of the femur is reduced and closely appressed to greater trochanter to form a trochanteric crest. The femoral head is distinct with a neck. It is oriented at a right angle to the femoral shaft. There is no trace of the fourth trochanter. On the caudal surface of the distal femoral part, the fossa poplitea is pronounced. The larger medial condyle of the distal end extends farther posteriorly than the smaller lateral condyle. The cranial part of the distal surface of the lateral condyle forms a distally protruding eminence. There is no patelar groove on the anterior surface of the distal end. A transverse groove is present on the ventral articular end of the distal end of femur.
The tibia is long and slightly laterally curved in its middle part. It is longer than the femur: the length ratio of tibia to femur is 1.33. The medial and lateral condyles of the proximal end directed caudally and they are deeply separated; the medial condyle is significantly more strongly protruded caudally relative to the lateral condyle. The medial edge of the medial articular facet forms a proximally directed eminence. The low fibular crest extends distally to the end of the proximal quarter of the bone. In proximal view, there is a ball-shaped process for articulation with the medial surface of the proximal end of the fibula. The distal end of the tibia is expanded,and the lateral malleolus of tibia has a strong distal projection.The astragalus and calcaneum are short and wide and completely fused. The ascending process of the astragalus is restricted and fails to cover medial surface of tibia.
The fibula is strongly reduced. Only a proximal portion of the left fibula is preserved. The lateral surface of the fibula is slightly convex, and its medial surface is flattened.
The metatarsal II is slightly shorter than metatarsal IV.They are unfused proximally. The fused distal tarsi adhere to the proximal ends of the metatarsal II and IV. In the distal part of the tarsometatarsus, metatarsals II and IV tightly envelop the sides of the proximal part of a very short distal metatarsal III rudiment, located entirely on the anterior side of the tarsometatarsus. There is a sharp, distally extended ridge on the posterior surface of the distal portion of the metatarsal III. The posterior surface of the proximal portion of the metatarsal III is slightly concave.
Only two digits and pedal claws are preserved. The phalangeal articular surfaces are well developed. There are three pedal ungual phalanges, which scattered near the skeleton,thus it is not sure, which digits they belong to. One is near the distal end of the metatarsal. The other two are isolated. The pedal ungual phalange is moderate in length and tapered, and slightly curved. There is a deep L-shaped groove on the lateral surface of the ungual, which is similar to that ofMononykus(Chiappe LM et al., 2002). The pedal ungual also bears a groove on ventral surface, but lacks a pair of small ventral foramina (Longrich NR and Currie PJ, 2009) that are present in Albertonykus borealis (Longrich NR and Currie PJ, 2009).
4. Systematic position of Qiupanykus
Qiupanykus is assigned to alvarezsaurid based on the following characters: short prezygapophyses in caudal vertebrae,procoelous caudal vertebrae ;the femur is slightly sigmashaped in posterior view ( Karhu AA and Rautian AS, 1996;Chiappe LM et al., 1996; Averianov A and Sues HD, 2017).
We conducted a phylogenetic analysis to investigate the systematic position of Qiupanykus among alvarezsaurid dinosaurs using the modified data matrix of Xu X et al. (2011).With the addition of undescribed alvarezsaurid (41HIII-0104)and Qiupanykus into the modified data matrix, 16 taxa (Tyrannosauridae and Dromaeosauridae as outgroups and 14 taxa as the ingroup), and 77 osteological characters were analyzed using TNT (Tree Analysis Using New Technology) version 1.1 (Appendix). A traditional search (tree bisection-reconnection swapping algorithm, 1000 random seeds, 1000 replicates,10 trees saved per replication) yielded 20 most parsimonious trees with 111 steps.
The strict consensus of the 20 most parsimonious trees(Fig. 3) shows that the two alvarezsaurid dinosaurs (41HIII-0104) and Qiupanykus) are nested in different clades.Qiupanykus is close to Xixianykus and Linhenykus. The apomorphies of the Qiupanykus are: 19 (1): posterior sacral vertebrae bearing a strong ventral keel; 22 (0): Proximal caudals with transverse processes centrally positioned on centrum; 49(1) pubic peduncle of ilium with reduced and knob-like articular surface for pubis and 64(0) fibular crest of tibia large and quadrangular.
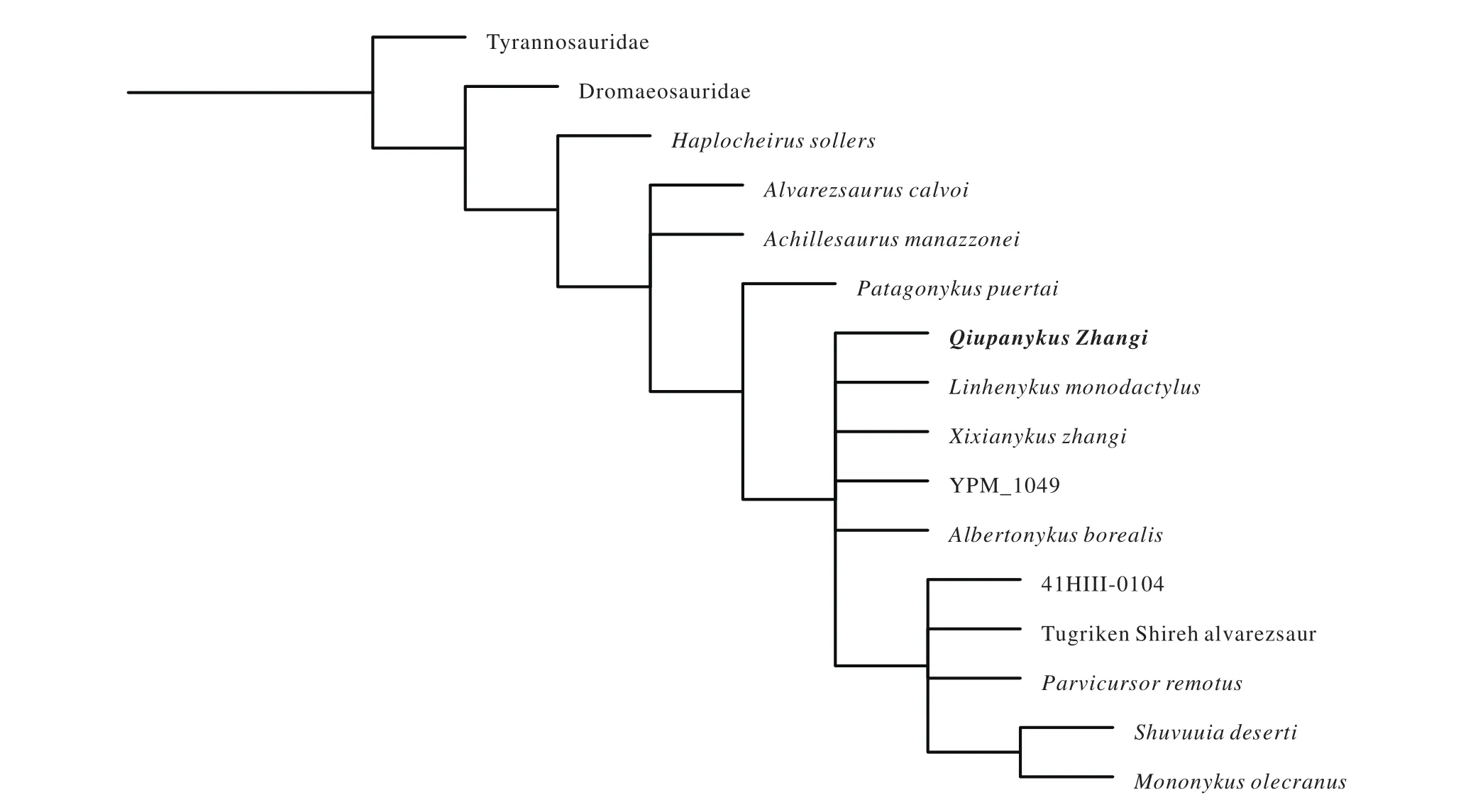
Fig. 3. The strict consensus of the 20 most parsimonious trees, showing the systematic position of Qiupanykus among alvarezsaurid dinosaurs.
5. Behaviour of alvarezsaurid dinosaurs
The skeleton of Qiupanykus is associated with an eggshell fragment near its tail. The thickness of the eggshell fragment is 1.8 mm. Some features, such as linearituberculate ornamentation type, including nodes and short ridges, two structural layers (non-prismatic), and relatively thick shell, are all identical to the Luanchuan oviraptorosaur eggshells (Tanaka K et al., 2011). Using the formula (Elongatoolithidae) by Tanaka K et al. (2016), the estimated egg mass of the eggshell is: Log10egg mass = 1.569×log10(1.8)+2.655, the thickness of the egg shell is 1.8 mm, then the Egg mass is 1136.377 g(95% CI: 715 g to 1809 g). The femur circumference of Qiupanykus is 17.09 mm. Based on the formula (log10BM =2.754×log10(femur circumference-0.683) by Campione NE et al. (2014), the estimated body mass for Qiupanykus: is 515 g (log10BM = 2.754×log1017.09-0.683). The estimated egg mass is much heavier than the estimated body mass of Qiupanykus. Thus, the egg could not be laid by Qiupanykus.
Alvarezsaurid dinosaurs bear highly specialized arms,whose purpose is still a mystery. They are regarded that the special arms were used to burrow (Perle A et al., 1993) or break open termite mounds (like modern anteaters), and possible to feed on insects (Senter P, 2005). However, the skeleton is associated with eggshell fragments from Luanchuan area, and the eggshell fragment morphologies indicate that those eggs belong to oviraptorid eggs (Tanaka K, personal communication, 2017). There is another case found from north-western Patagonia of Argentina, where an alvarezsaurid skeleton is preserved with eggs (Agnolin FL et al., 2012).Although Agnolin et al. thought the eggs associated with the alvarezsaurid Bonapartenykus ultimus were laid by an alvarezsaurid dinosaur, they pointed out that the external ornamentation patterns of Arriagadoolithus expressed on the outer shell surface is similar to elongathoolithid eggs. Arriagadoolithus was possibly laid by oviraptorosaurid dinosaurs.Thus, there are three possibilities about the relationship between the alvarezsaurid skeletons and eggs (egg fragments)associated with them: (1) Eggshell fragments were buried with alvarezsaurid skeleton by coincidence, and the eggshell is nothing to do with the skeleton. (2) The eggs were laid by alvarezsaurid dinosaurs and (3) The eggshell fragments were from eggs broken by alvarezsaurid dinosaurs and the eggs were not laid by them. However, considering the strong thumb claw of alvarezsaurid dinosaurs, we prefer to the third interpretation. Alvarezsaurid dinosaurs perhaps use their special claw to break eggs, and they are perhaps egg-eaters (Fig. 4).

Fig. 4. Life scene of Qiupanykus zhangi (drawn by Zhao Chuang).
6. Conclusion
The skeleton of Qiupanykus is associated with oviraptorid egg shell fragments suggesting that at least the derived alvarezsaurid dinosaurs may be an egg-eaters, which use their special arms (the strong thumb claws) to pierce the hard eggshell.Qiupanykus is a relatively small sized alvarezsaurid and it represents the youngest alvarezsaurid dinosaur from China so far.
Acknowledgements
We thank Dr Dale Winkler (Southern Methodist University, Dallas, TX, USA) for providing thorough comments on the earlier version of this Paper. We also appreciate the life scene of Qiupanykus drawn by Zhao Chuang. This study was supported by the National Natural Science Foundation of China (41672019 and 41688103) and the project from the China Geological Survey (20160126) to Junchang Lü.
Appendix
Character list and datamatrix for the phylogenetic analysis of Luanchuan alvarezsaurid dinosaurs (Modified from Xu et al., 2011 with additions of Qiupanykus and 41HIII-0104).
1. Tooth crowns serrated (0), or unserrated (1) (Perle et al., 1993)
2. Maxilla bearing teeth posteriorly (0), posterior of maxilla edentulous (1)(Perle et al., 1993)
3. Centra of cervical vertebrae amphiplatyan or biconvex (0), or opisthocoelous (1) (Perle et al., 1993)
4. Carotid processes of cervical vertebrae absent (0), or present (1)
5. Cervical vertebrae with tall neural spine (0), or neural spine low and ridgelike (Novas, 1996)
6. Cervical vertebrae with prominent epipophyses (0), or epipophyses reduced or absent (1)
7. Cervical centra with broad ventral surface (0), or with ventral surface narrow, 33% width of centrum or less (1), or ventral surface extremely narrow,25% width of centrum or less (2) (ORDERED)
8. Cervical centra with convex or flat sides (0), or cervical centra bearing deep lateral depressions (1)
9. Cervical centra with a flat or rounded ventral surface (0), or ventral surface bearing a prominent sulcus (1) (Novas, 1996)
10. Dorsal vertebrae amphiplatyan (0), opisthocoelous (1)(Perle et al., 1993)
11. Dorsal centra subcylindrical (0), laterally compressed and ventrally keeled (1) (Novas, 1996)
12. Dorsal vertebrae articulating via hyposphene-hypantrum joints (0), hyposphene-hypantrum articulations absent (1)(Perle et al., 1993)
13. Parapophyses of dorsal vertebrae below diapophyses (0), or parapophyses elevated to the level of the diapophyses (1) (Perle et al., 1994)
14. Parapophyses of dorsal vertebrae closely appressed to centrum (0), or parapophyses borne on elongate stalks lateral to centra (1)
15. Dorsal infradiapophyseal fossa: well developed (0) or hypertrophied, with infraprezygapophyseal fossa and infrapostzygapophyseal fossa not visible in lateral view (1) (Perle et al., 1994)
16. Postzygapophyses of dorsal vertebrae oriented horizontally (0), angled towards the saggital plane (1) (Novas, 1996)
17. First sacral and last dorsal articulating via amphiplatyan or opisthocoelous articulation (0), or procoelous articulation (1) (Perle et al., 1994)
18. First sacral centrum, ventral surface flat or rounded ventrally (0), or first sacrum bearing a strong ventral keel (1). Mononykus bears a weak ridge on the first sacral, and is coded as ‘0’. (Chiappe et al., 1998)
19. Posterior sacral vertebrae flat or ventrally rounded (0), or bearing a strong ventral keel (1), or bearing a hypertrophied ventral keel (2) (Perle et al., 1994;Novas, 1996) (ORDERED).
20. First caudal vertebra free (0), or incorporated into sacrum (1). Comparisons of Patagonykus and Mononykinae suggest that the latter have incorporated caudal 1 into the sacrum. The first caudal appears to be partially incorporated into the sacrum in the Tugriken Shireh alvarezsaurid.
21. Caudal vertebrae amphiplatyan (0), or procoelous (1) (Novas, 1996) 22.Proximal caudals with transverse processes centrally positioned on centrum(0), transverse processes anteriorly displaced (1)
23. Sternum: broad, flat plates (0), keeled and boatlike (1)
24. Coracoid and scapula with strong caudal projections bounding glenoid:absent (0), or present (1)
25. Coracoid with short ventrolateral process (0), elongate ventrolateral process (1) (Novas, 1996)
26. Coracoid with prominent biceps tubercle (0), biceps tuberclereduced or absent (1)
27. Coracoid convex cranially and concave caudally (0), coracoid platelike(1)
28. Scapular blade straight or curving caudally in lateral view (0), blade curves cranially in lateral view (1)
29. Humeral internal tuberosity hypertrophied and proximally projecting: absent (0), or present (1)
30. Deltopectoral crest of humerus 33% or less the length of the humerus (0),deltopectoral crest elongate and more than 33% the length of the humerus (1)
31. Humeral ectepicondyle small and proximally positioned (0), or hypertrophied and distally positioned (1) (Perle et al., 1994).
32. Ulna articulates with radial condyle of humerus (0), articulation between ulna and radial condyle lost (1) (Perle et al., 1994). Note: the distal end of the humerus retains a small, planar facet in Patagonykus (Novas, 1997) here interpreted as the radial condyle.
33. Distal articular surface of ulna: unexpanded or spatulate to permit lateral flexion of manus (0); or bulbous, trochlear articular surface extended onto dorsal surface of ulna to permit hyperextension (1)
34. Olecranon process of ulna weakly developed (0), or hypertrophied(1)(Perle et al., 1993)
35. Distal articular surface of radius unexpanded (0), or craniocaudally expanded (1) (Perle et al., 1994)
36. Radius/ulna articulation: radius articulates with cuplike cotyle of ulna (0),small, paired articular facets (1), joined by large, triangular symphysis (2)(Novas, 1996) (UNORDERED)
37. Metacarpal I and distal carpals separate (0), or fused to form a carpometacarpus (1) (Perle et al., 1993)
38. Metacarpal I elongate (0), or broad, and wider than long (1) (Novas,1996)
39. Metacarpal I with smooth medial edge (0), or prominent tuberosity (‘extensor process’) on medial surface (1)
40. Manual digit I slender (0), or subequal to humerus in diameter (1)
41. Manual phalanx I-1 or subcircular in section (0), or flattened and bearing a prominent ventral sulcus (1) (Perle et al., 1994)
42. Manual phalanx I-1 with proximolateral process projecting over proximal articular surface: absent (0), or present (1) (Perle et al., 1994)
43. Ventral surface of manual ungual I bearing axial groove: absent (0), or present (1)
44. Manual ungual I with well-developed flexor tubercle (0), flexor tubercle reduced to a low keel (1) (Novas, 1997), or absent (2) (Perle et al., 1994).(UNORDERED)
45. Lateral grooves of manual ungual I, unenclosed (0), proximal end of grooves partially enclosed by notches (1) (Chiappe and Coria, 2003), proximal end of grooves passing through ventral foramena (2) (Perle et al., 1994).(UNORDERED)
46. Proximal articular surface of manual ungual I taller than broad (0), or at least as broad as tall (1).
47. Cuppedicus fossa of ilium present (0), or absent (1) (Novas, 1996)
48. Pubic peduncle of ilium anteroposteriorly broad (0), or anteroposteriorly reduced (1) (Novas, 1997).
49. Pubic peduncle of ilium with well-developed articular surface for pubis(0), or pubic articular surface reduced and knob-like (1)
50. Posterior wing of ilium no longer than anterior wing (0), or posterior wing of ilium longer than anterior wing (1)
51. Posterior alae of ilia separated medially (0) or contact medially (1)
52. Acetabular rim of ilium terminates at end of pubic peduncle (0), or terminates above end of pubic peduncle (1)
53. Brevis fossa with strongly projecting medial shelf (0), or medial shelf of brevis fossa reduced to low ridge (1)
54. Pubis with pubic apron (0), pubic apron absent (1)
55. Pubis with well-developed pubic boot (0) pubic boot absent (1)
56. Pubes and ischia separate distally (0), or pubic shaft and ischium with extensive distal contact (1) (Perle et al., 1993)
57. Knob-like anterior pubic tubercle absent (0), or present below pubic-iliac contact (1) (Hutchinson and Chiappe, 1998)
58. Femoral lesser trochanter broad and winglike (0), or reduced and closely appressed to greater trochanter of femur to form a trochanteric crest (1) (Perle et al., 1994)
59. Prominent caudal flange running from greater trochanter to femoral shaft,absent (0) or present (1)
60. Femoral distal condyles project caudally (0), or extend inward to bound popliteal fossa (1) (Perle et al., 1994)
61. Femoral lateral condyle poorly developed (0), conical and distally projecting (1) (Novas, 1996)
62. Femoral medial condyle bulbous (0), or broad and flattened (1)
63. Tibia with single cnemial crest (0), tibia with accessory cnemial crest (1)(Perle et al., 1993)
64. Fibular crest of tibia large and quadrangular (0), or reduced and rounded(1)
65. Distal end of tibia symmetrical in anterior view (0), lateral malleolus of tibia with strong distal projection (1)
66. Hindlimb: ascending process of astragalus broad (0), or restricted and failing to cover medial surface of tibia (1) (Perle et al., 1994)
67. Calcaneum with large fibular facet (0), fibular facet reduced or absent (1)(Perle et al., 1993)
68. Shaft of metatarsal III subcircular in section and well-developed proximally (0), or shaft of metatarsal III triangular in section and proximally reduced (arctometatarsalian) (1) (Perle et al., 1993)
69. Dorsal surface of metatarsal III relatively narrow and flat (0) or transversely expanded and slightly concave (1)
70. Sharp, distally extended ridge on ventral surface of metatarsal III: absent(0), or present (1)
71. Metatarsal IV longer than II (0), or metatarsals II and IV subequal in length (1)
72. Digits II and IV subequal in length (0), or digit IV longer than II (1) 1
73. Digit III robust (0), or digit III more slender than either digit II or IV (1)
74. Proximal articular surface of pedal phalanx IV-4 undivided (0), or ventrally notched and horseshoe-shaped in proximal view (1)
75. Pedal phalanx IV-4 shorter than IV-3: present (0), or absent (1)
76. Pedal unguals with arcuate or Y-shaped lateral grooves (0), or deep, L-shaped grooves (1) (Chiappe et al., 2002)
77. Distal end of tibia with lateral malleolus anteroposteriorly compressed (0)or expanded (1) (Novas, 1996)
Tyrannosauridae
00000000?00000000000000000000000000000000000000000000000000000 000011?000000?
Dromaeosauridae
(01)00(01)0000?010010000000000000000000000000000000000000000000 10000000000000100100
Achillesaurus_manazzonei
??????????????????1?10????????????????????????111???1???????????0??0????????0
Alvarezsaurus_calvoi
??00011100??0?????1?10?011?0??????????????01011111?0??????0?????1?000000100??
Patagonykus_puertai
??10??21?0?00100??1010?0110?1?11?1?111011?0111110??1?000001011??1000???????10
Albertonykus_borealis
???????????????????????????????111?2??????1221?????????????????1???10 1???1??1
Mononykus_olecranus
1111012111111111112111111111111111121111111221110??1???11111111 11111101111011
Shuvuuia_deserti
11111111111111111121111111111111111211111112(12)111011?11111???????11?1101?????1
Parvicursor_remotus
?????????1?111???1??11???????????????????????????????111?1?11?111111 1?01??1??
YPM_1049
???????????????????????????????????????????????????1???????????????101???????
Tugriken_Shireh_alvarezsaur
??1011111?111111112111?1????????????111?11122?1101111111111111?1
11?110011?111
Xixianykus_zhangi
?????????1111111102011????????????????????????110??111?10110100111 111???????1
Haplocheirus
000011?0?0?000?1000000?11000101?11000001101010?001??00000000100 0??0000100?00?
Linhenykus
??111111111111111?2?01100?0???0?1???1111111211?10???????0?1010?11 1?1?00??1?01
41HIII-0104
????11?1?11111?1????1??11?10????????1111111201???111??????1????111?11?1111101
Qiupanykus
0??1?1111????????01110????????????????????????111???11??11?111101??1000????00
杂志排行

China Geology的其它文章
- A super-large graphite deposit discovered in granite rocks at Huangyangshan,Xinjiang, China
- The oldest shale gas reservoirs in southern margin of Huangling uplift, Yichang,Hubei, China
- A discovery of Silurian tight shale gas in Jianshi, Hubei, China
- Three source rocks discovered in the Mid-Lower Jurassic, Dunhuang Basin in China
- Metamorphic petrology and geology in China: A review
- New prospecting progress using information and big data of coal and oil exploration holes on sandstone-type uranium deposit in North China