发酵床垫料栽培基质重复利用对辣椒生长和基质性状的影响①
2018-01-17张晶,罗佳,马艳
张 晶,罗 佳,马 艳
(江苏省农业科学院农业资源与环境研究所,南京 210014)
基质栽培是设施农业无土栽培的最主要形式,它突破了传统设施栽培的连作障碍和土壤盐渍化等问题,代表了现代设施农业发展的趋势。草炭为广泛使用的传统基质原料,但来源受产地限制,运输成本高,且草炭属于不可再生资源,长期过量开采会对环境造成毁灭性破坏。因此,依据当地资源,研究开发农业废弃物作为基质原料已成一大趋势。大量研究表明,秸秆[1-2]、菇渣[3-4]、椰糠[5]、畜禽粪便[6-9]等,在经过发酵堆肥后,均可作为栽培基质在生产上使用。
与传统的土壤栽培相比,基质栽培具有诸多优点,然而基质的高成本和重复利用是制约基质栽培发展的 2个主要因素[10],合理地重复利用基质可有效降低生产成本与劳动强度。然而,无论是何种基质,都存在栽培后的污染问题,如何将使用过的旧基质重新利用已成为研究者关注的新问题。程智慧等[11]研究表明栽培 1茬辣椒后的混合农作物基质可以在蔬菜育苗中继续使用。李威等[12]研究发现在连茬种植番茄的有机基质中轮作蒜苗后再种植番茄,番茄生长、产量和品质均能达到理想的效果,而且还能延长栽培基质的使用年限。蒋卫杰等[13]研究了以不同农业废弃物为主的混合基质连茬种植番茄后的损耗情况,结果表明不同基质损耗程度差异较大,葵花杆、菇渣和玉米秸的损耗程度远大于草炭和锯末。
发酵床养殖技术是基于控制畜禽粪尿排放与污染的一种新型生态养殖模式。有研究表明,养殖后的废弃垫料中富含有机质和氮、磷、钾等养分,资源化利用价值巨大[14]。合理有效利用发酵床废弃垫料资源,不仅能延伸产业链、提高经济效益,还可以实现废物再利用,促进生态农业的可持续发展。目前,关于发酵床垫料的资源化利用研究相对较少,且多是围绕其作为有机肥料直接施用土壤的可行性方面[14-16],关于垫料基质化利用方面则未见相关报道。本试验以发酵床垫料复合基质为材料,研究新配基质和旧基质在辣椒种植上的应用效果以及种植不同茬次辣椒后基质理化和微生物学性状的变化情况。本研究结果可为发酵床垫料的资源化利用提供新的思路,既能提高废弃垫料的附加值,又能降低基质栽培成本,从而促进循环农业的健康发展。
1 材料与方法
1.1 供试材料
供试基质材料为充分腐熟的秸秆发酵床养猪垫料与蛭石、珍珠岩和泥炭,按一定比例混合制成。基本理化指标如下:体积质量 0.24 g/cm3、总孔隙度72.06%、pH 6.77、EC值 3.69 mS/cm、全氮15.0 g/kg、全磷28.4 g/kg、全钾21.6 g/kg、速效氮8.2 g/kg、有效磷168 mg/kg、速效钾4.5 g/kg。供试土壤取自江苏省南京市六合区,基本理化性状如下:有机质17.3 g/kg,全氮1.1 g/kg,有效磷77 mg/kg,速效钾131 mg/kg,pH 7.6,EC值0.31 mS/cm。供试辣椒品种为苏椒14,由淮安市农业科学研究院提供。
1.2 试验设计
试验于2015年3—7月在江苏省农业科学院六合动物科学基地设施大棚进行。大棚长42 m,宽8 m。试验共设置3个处理:对照,土壤栽培(CK);新基质栽培处理(1茬);旧基质栽培处理(2茬),旧基质为种完1茬辣椒后的基质。采用槽式基质栽培。基质槽横切面为等腰梯形,上口宽25 cm,下口宽23 cm,高23 cm,总长4 m,槽内铺设滴灌带用于日常浇水。每槽装基质0.2 m3,种植1行辣椒,株距40 cm,种植10棵,每个处理3次重复,大棚内各处理随机排列。辣椒于2015年3月27日移栽定植,生长期间采取统一追肥管理,定植后15 d进行第1次追肥,以后分别在盛花期和结果期进行第2次和第3次追肥,3次追肥量一致。每次追肥量为每个槽 0.2 kg N-P2O5-K2O(15∶15∶15)复合肥,追肥方式采用条施。其他田间管理均按常规进行。6月11日开始采收,7月29日收获完毕。
定植30 d后每个处理取3棵苗测定株高、根长和整株鲜重。辣椒收获后统计产量,每个基质槽用土钻采取4个点的基质,混匀后用于理化和微生物学性状测定。
1.3 测定项目与方法
1.3.1 基质理化性质测定 基质体积质量和总孔隙度测定参考连兆煌[17]《无土栽培原理与技术》。取一已知体积(V)的容器,称重(W1),装满自然风干的待测基质,称重(W2),将装有基质的容器浸泡在水中24 h后,称重(W3);体积质量 =(W2-W1)/V;总孔隙度(%)=(W3-W2)/V×100(重量以g为单位,体积以cm3为单位)。基质养分测定参考鲍士旦[18]《土壤农化分析》。速效氮采用KCl浸提,流动分析仪测定;有效磷采用NaHCO3浸提,钼锑抗比色法测定;速效钾采用NH4Ac浸提,火焰光度法测定;全碳和全氮测定采用碳氮元素分析仪;全磷测定采用钼锑抗比色法;全钾测定采用火焰光度计法。将风干基质与去离子水以1:5比例混合振荡,静置后取上清,分别用电导率仪和pH计测定EC和pH。
1.3.2 基质中重金属含量测定 重金属 Cu、Zn、Pb、Hg、As、Cd和Cr含量的测定方法参照国家有关标准执行[19]。
1.3.3 基质总DNA提取 称取0.3 g基质样品,按照MPbio土壤DNA提取试剂盒(FastDNA SPIN Kit for Soil)操作说明书提取基质样品总DNA,提取的总DNA放置-20 ℃冰箱保存备用。
1.3.4 基质总细菌和真菌数量测定 基质总细菌和真菌的数量利用实时荧光定量 PCR(Real-Time PCR)测定[20]。真菌定量扩增采用引物 NS1/Fungi(5′-GTAGTCATATGCTTGTCTC-3′/ 5′-ATTCCCCGTTACCCGTTG-3′),细菌定量扩增采用引物347F/531R(5′-GGAGGCAGCAGTRRGGAAT-3′/5′-CTNYGTMT TACCGCGGCTGC-3′)。定量扩增采用 ABI 7500 荧光定量PCR仪进行,扩增条件为:10 µl SYBR Premix Ex Taq,0.4 µl上游引物和下游引物,0.4 µl ROX Reference Dye Ⅱ,2 µl模板 DNA 和 6.8 µl无菌水。扩增程序为: 95 ℃预变性30 s;95 ℃预变性5 s,60 ℃延伸34 s,循环40次。根据各样品Ct值计算每克基质所含的拷贝数。
1.3.5 基质微生物群落结构测定 采用变性梯度凝胶电泳(denaturing gradient gel electrophoresis,DGGE)测定基质微生物群落结构。细菌PCR扩增采用16 S rDNA通用引物Eub338/Eub518(5′-GC clamp-ACTCCTACGGGAGGCAGCAG-3′/5′-ATTACCGCG GCTGCTGG-3′)[21];真菌 PCR扩增采用18 S rDNA通用引物 NS1/Fungi(5′-GTAGTCATATGCTTGTCTC-3′/5′-GC clamp-ATTCCCCGTTACCCGTTG-3′)[22],其中 GC clamp 为 5′-CGCCCGCCGCGCCCCGCGCCC GGCCCGCCGCCCCCGCCCC-3′。PCR扩增反应体系:10×PCR buffer(Mg2+free)5 µl,Mg2+(25 mmol/L)4 µl,dNTPs(各 2.5 mmol/L)4 µl,引物(10 µmol/L)各 1 µl,模板 DNA 1 µl,Taq DNA 聚合酶(5 U/µl)0.3 µl,加入无菌水补足至50 µl。PCR扩增条件:94 ℃ 5 min;94 ℃ 45 s,60 ℃ 45 s(真菌退火温度为 57 ℃),72 ℃45 s,循环32次;72 ℃ 10 min。采用D-Code点突变检测系统(D Code Universal Mutation Detection System,BIO-RAD,USA)对 PCR产物进行 DGGE 分析。聚丙烯酰胺凝胶浓度为8%,细菌变性剂梯度为40%~60% (100%的变性剂为7 mol/L尿素和40% 去离子甲酰胺的混合物),真菌变性剂梯度为 20% ~40%。电泳条件:80 V电压、60 ℃恒温电泳16 h,电泳结束后银染胶片并扫描保存。
1.4 数据分析
试验数据经Excel处理后应用SPSS 22.0软件进行统计分析,采用单因素方差分析比较处理间差异的显著性水平。DGGE电泳图谱分析采用Quantity One软件。
2 结果与分析
2.1 栽培基质中主要重金属含量
新配栽培基质中主要重金属含量见表1。Zn、Pb、Hg、As、Cd和Cr这6种重金属含量均在我国土壤环境质量标准(GB 15618-1995)[23]中农业用地标准(二级标准)的限量范围内,Cu含量超出二级标准,但在三级标准以内(≤400 mg/kg)[23]。目前为止,我国尚未制定有机基质产品中重金属含量的限定标准,由于基质的使用性质类似于农田土壤,因此我们以国家土壤环境质量标准(GB 15618-1995)为参照。总体来说,基质中Cu和Zn含量较高,分析可能的原因是养殖场所用的饲料中Cu、Zn含量较高,导致了以垫料为主要原料的基质中Cu和Zn含量高。

表1 基质中主要重金属含量Table 1 Contents of major heavy metal elements in substrate
2.2 有机基质栽培对辣椒生长发育和产量的影响
由表2可以看出,定植30 d后,有机基质栽培的第1茬辣椒与土壤栽培对照(CK)相比,在株高、根长和整株鲜重方面均无显著差异。此外,第2茬辣椒和第1茬以及对照相比,在株高、根长和整株鲜重方面也均无显著性差异,整株鲜重与第1茬和对照相比分别减少 11.25% 和 12.72%。有机基质栽培的辣椒产量与对照相比,无显著差异。其中,第2茬辣椒产量最高,达到21.92 kg,较对照(19.98 kg)相比,高出了8.85%;第1茬辣椒产量为18.86 kg,与对照相比,减少了5.61%,处理间差异不显著。第2茬和第1茬相比,产量增加了 13.96%,处理间差异不显著。结果表明,以发酵床垫料为主要原料的有机基质适用于辣椒栽培,且基质连续使用2茬对辣椒的生长发育及产量没有明显影响,这样可显著降低基质栽培成本。
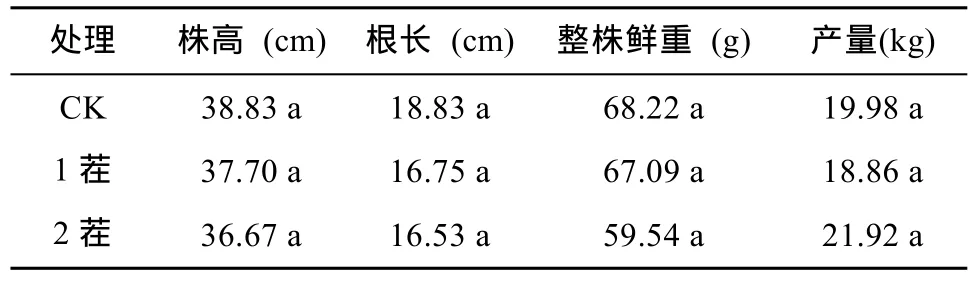
表2 不同茬次辣椒生长发育指标及产量结果Table 2 Growth indexes and yields of peppers under different stubbles
2.3 辣椒连茬种植后有机基质理化性状的变化特征
由表3可以看出,种植第1茬辣椒对基质的理化性状影响最大,基质的体积质量、EC、pH和大部分养分含量与新基质(0茬)相比均发生显著性变化。其中,基质的体积质量和pH显著增加,体积质量由0.24 g/cm3升高至0.35 g/cm3,pH由6.77升高至7.93;EC值急剧降低,由3.69 mS/cm降低至0.695 mS/cm,减少了81.17%;全碳、全氮、全钾、速效氮和速效钾含量1茬后显著降低,全碳含量由167.3 g/kg降低至151.4 g/kg,全氮含量由15.0 g/kg降低至12.6 g/kg,全钾含量由21.6 g/kg降低至17.8 g/kg,速效氮含量由8 244.4 mg/kg降低至646.8 mg/kg,减少了92.15%,速效钾含量由4 545.7 mg/kg降低至674.5 mg/kg,减少了85.16%;全磷含量1茬后显著增加,由28.4 g/kg升高至33.1 g/kg;基质总孔隙度和有效磷含量1茬后变化不显著。

表3 辣椒连茬栽培对基质理化性状的影响Table 3 Effects of continuous pepper planting on physical and chemical properties of substrate
种植2茬辣椒后基质大部分理化指标与1茬时相比变化不显著,其中基质体积质量、总孔隙度、EC、pH、全磷、全钾、速效钾和速效氮等指标变化均不显著;全碳和全氮含量显著降低,分别由151.4 g/kg和12.6 g/kg降低至136.5 g/kg和11.4 g/kg;有效磷含量由157.1 g/kg升高至172.3 g/kg,显著增加。
种植 2茬辣椒后基质大部分理化指标与新基质相比均发生显著变化,其中EC值、速效氮和速效钾含量变化最为明显,EC值由3.69 mS/cm降低至 0.507 mS/cm,减少了 86.26%;速效氮含量由8 244.4 mg/kg降低至614.4 mg/kg,减少了92.55%;速效钾含量由4 545.7 mg/kg降低至593.4 mg/kg,减少了86.95%。
综上,种植1茬、2茬辣椒后的有机基质与新基质相比,大部分理化指标均发生了显著变化,其中EC值、速效氮和速效钾含量变化最为明显,均大幅度降低,这可能与种植过程中的淋洗作用有关。种植2茬辣椒后的基质与1茬时相比,大部分理化指标变化不显著,说明种植1茬辣椒后,基质的理化性状趋于稳定,且基质的体积质量、总孔隙度和EC值均在适宜作物生长范围内[24],这样有利于旧基质的重复使用。然而,在配合施肥基础上,该有机基质能否继续种植第 3茬辣椒而不影响其产量以及其理化性状是否会发生显著改变有待于进一步研究。
2.4 辣椒连茬种植后有机基质微生物学性状的变化特征
2.4.1 总细菌和总真菌数量变化 种植不同茬次辣椒后基质中微生物数量变化情况如表4所示。Real-Time PCR结果表明:连续种植2茬辣椒对基质中细菌数量影响不大,1茬后细菌总量为 3.25×1015copies/g基质,2茬后细菌总量为 3.08×1015copies/g基质,与新基质(0茬)相比(2.42×1015copies/g基质)数量均稍有增加,但差异均不显著。栽种辣椒后基质中真菌数量变化显著,1茬后真菌数量显著降低,由6.34×109copies/g基质减少至1.36×109copies/g基质,2茬后基质中真菌数量与1茬时相比差异不显著,总量为1.59×109copies/g基质。

表4 辣椒连茬栽培对基质细菌和真菌数量的影响(copies/g基质)Table 4 Effects of continuous pepper planting on populations of bacteria and fungi in substrate
2.4.2 微生物群落结构变化 种植不同茬次辣椒后基质中细菌和真菌DGGE图谱如图1所示。利用Quantity One软件对DGGE图谱进行聚类分析(图2)和多样性分析(表 5)。细菌多样性分析结果显示,种植 1茬、2茬辣椒后,基质中细菌的丰度分别为 22和21,与新基质(19)相比,均略有增加。多样性指数间差异性与丰度结果一致,种植1茬、2茬辣椒后,基质中细菌的多样性指数分别为2.746 8和2.766 9,与新基质(2.684 0)相比,均略有增加。DGGE图谱直观显示:种植1 茬和2茬辣椒后基质样品间细菌条带相似度高,与新基质(0茬)相比,优势条带位置差异明显,且一些共有条带亮度差异显著。聚类分析结果也显示新基质明显区分于种植 1茬辣椒和种植 2茬辣椒后的基质,且种植1茬辣椒和种植2茬辣椒后的基质样品聚成一大类,说明这两个处理间的细菌群落结构相似度高。以上结果表明初次种植辣椒能改变有机基质中优势细菌群落,而种完1茬辣椒后基质中优势细菌种类趋于稳定。
真菌多样性分析结果显示,种植1茬、2茬辣椒后,基质中真菌的多样性指数和丰度与新基质相比明显提高。1茬和2茬基质的多样性指数相差不大,分别为3.221 6和3.314 3,均显著高于新基质(2.309 8);丰度分别为28和31,同样都显著高于新基质(13)。DGGE图谱也显示,种植1 茬和2茬辣椒后基质样品间真菌条带相似度高,与新基质相比,条带数目明显增多。聚类分析结果也与细菌一致,1茬和 2茬基质样品聚为一类,且明显区分于0茬样品。以上结果表明初次种植辣椒能显著改变有机基质中真菌群落结构,而种完1茬辣椒后基质中优势真菌类群趋于稳定。
综上,种植第1茬辣椒能明显改变有机基质中优势微生物群落,并能显著提高基质中真菌多样性,但连续种植 2茬辣椒对基质中优势微生物群落影响较小,细菌和真菌群落结构变化不大。然而,继续种植第 3茬辣椒后基质中优势微生物群落是否会发生明显改变仍有待进一步研究。
3 讨论

图1 辣椒连茬栽培对基质中细菌(A)和真菌(B)群落结构影响的DGGE图谱Fig. 1 Effects of continuous pepper planting on community structures of bacteria (A) and fungi (B) in substrate
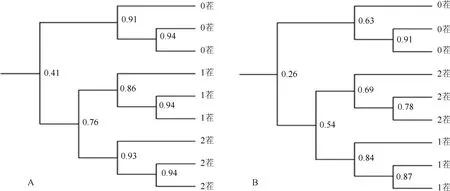
图2 辣椒连茬栽培对基质中细菌(A)和真菌(B)群落结构影响的DGGE图谱聚类分析Fig. 2 Cluster diagrams of bacteria (A) and fungi (B) DGGE profiles for continuous pepper planting substrate

表5 辣椒连茬栽培对基质中细菌和真菌多样性指数、丰度和均匀度影响Table 5 Effects of continuous pepper planting on Shannon-Wiener index, Richness and Evenness of bacterial and fungal communities in substrate
发酵床养殖中,畜禽粪尿直接排放在发酵床上,由于饲料中添加的重金属大部分会随粪便排出体外,导致垫料中不可避免地含有一些对环境和生物有害的重金属元素。所以将垫料复配成有机基质资源化利用前,有必要对基质中重金属含量进行检测。本研究测定结果表明,新配基质样品中重金属元素Cu、Zn、Pb、Hg、As、Cd和Cr虽然均能检出,但含量均处于国家相关标准限量范围之内,其中Cu和Zn含量相对较高,有超标风险。本研究结果表明,辣椒栽培过程中基质pH升高成碱性,有利于降低重金属生物有效性[25],由此推测发酵床垫料基质中重金属污染风险较小。然而,为确保发酵床垫料资源化利用顺利实施,建议在发酵床养殖过程中控制高含量Cu、Zn饲料的添加,从源头阻控风险。本研究从发酵床垫料资源化利用和改善辣椒种植条件出发,将以发酵床垫料为主要原料的有机基质用于设施辣椒栽培。结果表明,无论是第1茬还是第2茬辣椒在植株长势和最终产量上与土壤栽培的辣椒相比均无显著差异,说明发酵床垫料的基质化利用思路是可行的,且用完1茬的旧基质可以重复使用。
在基质栽培过程中,栽培基质只有具备适宜的理化性质,才能为作物生长提供良好的根际环境。发酵床垫料中含有较高浓度的盐分,将其添加到有机基质中可能导致基质EC值升高,继而影响作物正常生长。本研究所用的发酵床垫料基质初始 EC值高达 3.69 mS/cm,从辣椒栽培结果来看,高EC值并未对辣椒生长和最终产量产生影响,说明本研究所选用的辣椒品种对高盐分环境不敏感。为了更好推广发酵床垫料基质化利用新思路,筛选并丰富适宜发酵床垫料基质栽培的作物种类是今后研究工作的重心之一。
郭世荣[24]提出关于理想基质的标准:体积质量在 0.1~0.8 g/cm3范围,总孔隙度在 54%~96%范围,pH 7.0左右,作物栽培效果较好。本试验的测定结果表明,连续栽种 2茬辣椒后基质体积质量为 0.34 g/cm3,总孔隙度为69.61%,EC值为0.507 mS/cm,均在适宜范围。栽培后基质pH有所上升,而栽培结果表明,基质pH偏碱性对辣椒生长没有明显影响。新配基质的养分测定结果表明,发酵床垫料作为主要原料贡献了丰富的氮磷钾等营养元素,其中速效氮和速效钾含量高达8 244.4 mg/kg和4 545.7 mg/kg。然而,由于基质的保肥性能较差,速效养分很容易在浇水过程中淋洗损失,再加上辣椒生育期长,因此在栽培过程中还需追施化肥。在配合施肥的基础上,种植1茬、2茬的基质,各种营养元素基本能满足辣椒生长所需。综上,发酵床垫料复合基质连续种植2茬辣椒不影响其产量主要原因可能是该基质良好的理化性状和丰富的养分支持。
基质在栽培完作物后,受外界环境、作物本身及施肥的影响,其理化性状均会发生变化。本研究测定结果表明,种植1茬、2茬辣椒后基质EC值和速效氮、速效钾含量大幅度降低,这可能与种植过程中的淋洗作用有关。为了防止淋洗液污染土壤和地下水,应该采取相应的防范措施,比如可以在基质栽培场所铺上地膜,阻止淋洗液渗入土壤。
PCR-DGGE技术能够更加直观地比较和分析微生物群落结构的变化规律,因而具有不可替代的优势,被广泛应用。马宁宁和李天来[26]利用PCR-DGGE技术研究裸地及不同连作年限的设施番茄栽培土壤中细菌和真菌群落结构时发现,设施条件下栽培番茄明显改变了土壤土著细菌的群落结构,但连作年限对其影响较小;土著真菌的群落结构稳定性优于土著细菌,但其优势种群在不同连作年限的土样中变化较大。陆海飞等[27]研究发现长期有机无机肥配施可显著提高土壤细菌多样性,并改变土壤细菌和真菌的群落结构。关于土壤栽培条件下的微生物群落结构变化研究较多,而对于有机基质栽培中微生物群落结构变化的相关报道很少。本研究利用PCR-DGGE技术检测种植辣椒前后的基质,结果表明种植第1茬辣椒明显改变了基质中细菌和真菌的群落结构,并能明显提高基质中真菌的群落多样性,但连续栽种2茬辣椒对基质中细菌和真菌多样性影响较小,细菌和真菌群落结构变化不大。
4 结论
本研究表明以发酵床垫料为主要原料的有机基
质可以作为辣椒的栽培基质,且连续种植2茬辣椒均不影响其产量。连续使用2茬后,基质的主要理化性状:体积质量、总孔隙度和EC值均在作物适宜生长范围内,说明该基质性状稳定,适宜重复使用。从养分测定结果来看,发酵床垫料基质中丰富的氮磷钾营养元素配合施用适量化肥可以满足辣椒生长所需。PCR-DGGE检测结果表明种植第1茬辣椒明显改变了基质中细菌和真菌的群落结构,并能明显提高基质中真菌的群落多样性,但继续种植第2茬辣椒对基质中细菌和真菌多样性影响较小,细菌和真菌群落结构变化不大。然而,该基质能否继续种植第3茬甚至更多茬辣椒而不影响其产量以及基质的理化和微生物学性状是否会发生显著改变仍需进一步研究。
[1] Belal E B, El-Mahrouk M E. Solid-state fermentation of rice straw residues for its use as growing medium in ornamental nurseries[J]. Acta Astronautica, 2010, 67(9):1081-1089
[2] 崔元玗, 张升, 孙晓军, 等. 棉花秸秆为蔬菜栽培基质的可行性研究[J]. 北方园艺, 2012(19): 37-38
[3] Medina E, Paredes C, Pérez-Murcia M D, et al. Spent mushroom substrates as component of growing media for germination and growth of horticultural plants[J].Bioresource Technology, 2009, 100(18): 4227-4232
[4] Zhang R H, Duan Z Q, Li Z G. Use of spent mushroom substrate as growing media for tomato and cucumber seedlings[J]. Pedosphere, 2012, 22(3): 333-342
[5] 王必尊, 何应对, 唐粉玲, 等. 基于椰糠配比基质对香蕉组培苗生长的影响[J]. 江苏农业科学, 2013, 41(2):146-149
[6] 董晓涛, 杨志. 家畜粪便发酵液对基质栽培茄子生长发育及产量的影响[J]. 江苏农业科学, 2010(3): 187-189
[7] 李志刚, 刘晓刚, 李健. 硫酸铵与鸡粪配比在含生物质炭育苗基质中的应用效果[J]. 中国土壤与肥料, 2012(1):83-88
[8] 张宏, 沈章军 阳贵德, 等. 鸡粪改良对铜尾矿基质中无机氮组分及 3种豆科植物生长发育的影响[J]. 农业环境科学学报, 2011, 30(11): 2285-2293
[9] 张舒玄, 常江杰, 李辉信, 等. 奶牛粪蚯蚓堆肥的基质配方及对草莓育苗的影响[J]. 土壤, 2016, 48(1): 59-64
[10] 李国景, 徐志豪, Benoit F, 等. 环保型可重复利用的海棉无土栽培基质的应用研究[J]. 浙江农业学报, 2001,13(2): 61-66
[11] 程智慧, 于艳辉, 张庆春. 辣椒栽培有机基质在蔬菜育苗中的二次利用初探[J]. 西北农业学报, 2010, 19(4):123-126
[12] 李威, 孟焕文, 程智慧, 等. 轮作叶菜对大棚番茄连作基质重复利用效果的影响[J]. 西北农林科技大学学报(自然科学版), 2012, 40(1): 164-170
[13] 蒋卫杰, 余宏军, 刘伟. 无土栽培基质重复利用对番茄生长、产量和基质性状的影响[J]. 厦门大学学报, 2001,40(z2): 37-42
[14] 胡海燕, 于勇, 张玉静, 等. 发酵床养猪废弃垫料的资源化利用评价[J]. 植物营养与肥料学报, 2013, 19(1): 252-258
[15] 罗佳, 刘丽珠, 王同, 等. 养猪发酵床垫料有机肥对辣椒产量及土壤微生物多样性的影响[J]. 土壤, 2015, 47(6):1101-1106
[16] 刘宇锋, 罗佳, 严少华, 等. 发酵床垫料特性与资源化利用研究进展[J]. 江苏农业学报, 2015, 31(3): 700-707
[17] 连兆煌. 无土栽培原理与技术[M]. 北京: 中国农业出版社, 1994: 61-62
[18] 鲍士旦. 土壤农化分析. 3版[M]. 北京: 中国农业出版社,2000: 30-106
[19] 中国石油和化学工业协会. 肥料中砷、镉、铅、铬、汞生态指标(GB/T 23349-2009)[S]. 北京: 中国标准出版社,2009
[20] Fierer N, Jackson J A, Vilgalys R, et al. Assessment of soil microbial community structure by use of taxon-specific quantitative PCR assays[J]. Applied and Environmental Microbiology, 2005, 71(7): 4117-4120
[21] Nakatsu C H, Torsvik V, Ovreas L. Soil community analysis using DGGE of 16S rDNA polymerase chain reaction products[J]. Soil Science Society of America Journal, 2000, 64(4): 1382-1388
[22] May L A, Smiley B, Schmidt M G. Comparative denaturing gradient gel electrophoresis analysis of fungal communities associated with whole plant corn silage[J].Canadian Journal of Microbiology, 2001, 47(9): 829-841
[23] 国家环境保护局. 土壤环境质量标准(GB 15618-1995)[S]. 北京: 标准出版社, 1995
[24] 郭世荣. 无土栽培学[M]. 北京: 中国农业出版社, 2003:135-142
[25] Vega F A, Covelo E F, Andrade M L. Competitive sorption and desorption of heavy metals in mine soils: Influence of mine soil characteristics[J]. Journal of Colloid and Interface Science, 2006, 298(2): 582-592
[26] 马宁宁, 李天来. 设施番茄长期连作土壤微生物群落结构及多样性分析[J]. 园艺学报, 2013, 40(2): 255-264
[27] 陆海飞, 郑金伟, 余喜初, 等. 长期无机有机肥配施对红壤性水稻土微生物群落多样性及酶活性的影响[J]. 植物营养与肥料学报, 2015, 21(3): 632-643
