西非塞内加尔盆地海相优质烃源岩控制因素讨论
2018-01-12王建新孙玉梅杨永才
孙 涛 , 王建新, 孙玉梅, 杨永才
(中海油研究总院, 北京 100028)
西非塞内加尔盆地,位于巨型的被动大陆边缘MSGB(毛塞几比)盆地中,此盆地北起毛里塔尼亚,向南延伸至几内亚。塞内加尔占盆地的主要地区,面积约为95 000 km2。塞内加尔盆地的石油勘探始于20世纪50年代,陆续获得少量的油气发现,由于规模太小,这些发现均未投入开发[1]。经历了多年油气勘探低潮之后,近三年来在盆地的深水区块获得多个商业性油气田发现,如F油田储量达2.5亿桶,S油田储量达3.3亿桶,打开了塞内加尔盆地油气勘探的新局面。油源对比结果显示烃源岩为赛诺曼—土伦阶海相富有机质页岩,这套富有机质页岩,其有机碳(TOC)最大可达40%,为海相优质烃源岩。缺氧保存条件和生产力是有机质富集的两个最基本要素,这点已得到广泛认可。但是,对于二者之间哪个更重要存在“保存模式”和“生产力模式”之争。Schlanger 和Jenkyns[2]于1976年提出了“大洋缺氧事件”(OAE)模式,即大洋洋流的停滞、底层大洋氧气供应不足而导致海洋氧含量最小层(OMZ)的扩张,使得深部大洋缺氧,进而导致的有机碳的高效率保存—此为大洋缺氧事件的“保存模式”。然而,关于缺氧事件的起因和发展仍存争论。此后,随着碳同位素地层学和古生物学的深入,发现黑色页岩的分布与海相碳酸盐岩碳同位素的正偏[3]、海洋底栖生物的灭绝和浮游生物的大幅度更替有关[4],暗示大洋缺氧事件对应着大洋中碳库的显著转移,并且对应着海洋生产力的提高[5],因此黑色页岩的形成很可能是海洋生产力在短时期内迅速提升的产物—此为“生产力模式”。这两个模式的区别在于,前者需要一个近乎停滞的大洋环境;而后者要求大洋洋流循环的加速,以便大洋底部的营养元素能够被输送到大洋的表层,促进表层生产力的提高。
本文以塞内加尔盆地为研究对象,从有机地球化学的角度探讨了晚白垩世赛诺曼—土伦时期古海洋生产力和保存条件,从而明确海相优质烃源岩的控制因素,为今后该地区的油气勘探提供技术支持。
1 区域地质背景
塞内加尔盆地位于非洲西北海岸中部(图1),包括了毛里塔尼亚、塞纳加尔、几内亚及几内亚-比绍的部分海岸区域,盆地的形状大致呈现南北方向,其坐标经纬度为: 22°~10° N,10°~2°W。主要转换断层把盆地划分三个次盆,分别为:① 毛里塔尼亚次盆,其范围包括塞内加尔河至西萨哈拉南部; ②北部次盆,范围为冈比亚河和塞内加尔河之间的区域;③卡萨芒斯次盆,其范围从冈比亚河南部向南延伸至几内亚比绍(图1)。盆地类型为典型的大西洋型被动大陆边缘型盆地[6-8]。

图1 塞内加尔盆地及部分钻井位置图 [5]
塞内加尔盆地的形成和演化与非洲板块与北美洲分离紧密相关(超级大陆Pangaea的解体),其分离开始于中生代三叠纪[9-11],盆地演化可以分为前裂谷(元古代-晚泥盆世)、裂谷期(二叠纪—早侏罗世)和漂移期(中侏罗世-全新世)3个演化阶段(图2)。
前裂谷期沉积包括前寒武系-泥盆系(图2),地震资料揭示地层厚度超过5 000 m,以陆相碎屑岩为主;裂谷期主要发育膏盐层,膏盐层的最大厚度超过2 000 m(图2)。漂移期为典型的海相沉积,主要岩性为海相碳酸盐岩,其次为泥页岩及泥灰岩。其中,晚白垩世的赛诺曼-土伦阶主要岩性为泥页岩,该套泥页岩富含有机质,为该盆地的主力烃源岩。
2 晚白垩世赛诺曼-土伦阶烃源岩有机质母源地球化学分析
本次研究选择了11口钻井,分别为Mb-1、DN-2、Gd-1、Rd-1、CM-1、CM-2、CM-7、CM-10、F-1、S-1、DSDP 367,钻井的位置见图1。根据前人的研究,其中Mb-1、DN-2、Gd-1及Rd-1钻遇的赛诺曼—土伦阶沉积于陆架环境,CM-7、CM-10钻遇的赛诺曼—土伦阶沉积于半深海环境[12]。DSDP 367的岩心样品没有任何浊积岩,并且发育细的纹层以及较低碳酸盐含量,为典型的深海沉积环境[13]。
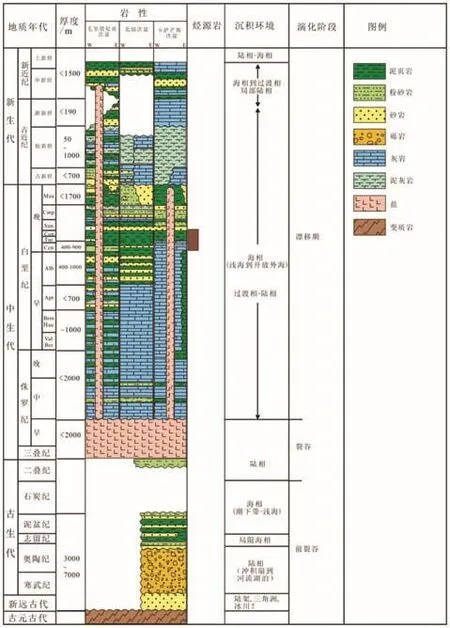
图2 塞内加尔盆地地层柱状图 [IHS, 2012]
2.1 TOC和岩石热解有机质母源特征
塞内加尔盆地C-T陆架沉积(Mb-1、DN-2、Gd-1及Rd-1)的样品的TOC值通常小于3%,氢指数(HI)通常小于300 mg HC/gTOC,投点至HI/OI曲线[14],有机质主要为II2-III型 (图3B),暗示了陆源有机质输入占优势[15];而半深海C-T样品(CM-7和CM-10)TOC通常大于3%,最高可达8.7% (图3A),HI通常大于300 mg HC/ g TOC,最高可达660 mg HC/g TOC(表1),有机质类型主要为II型(图3B),反映了水生生物输入为主。深海样品DSDP 367 TOC值高达40%,HI介于300~900 mg HC/g TOC之间,有机质类型主要为II型(表1)。
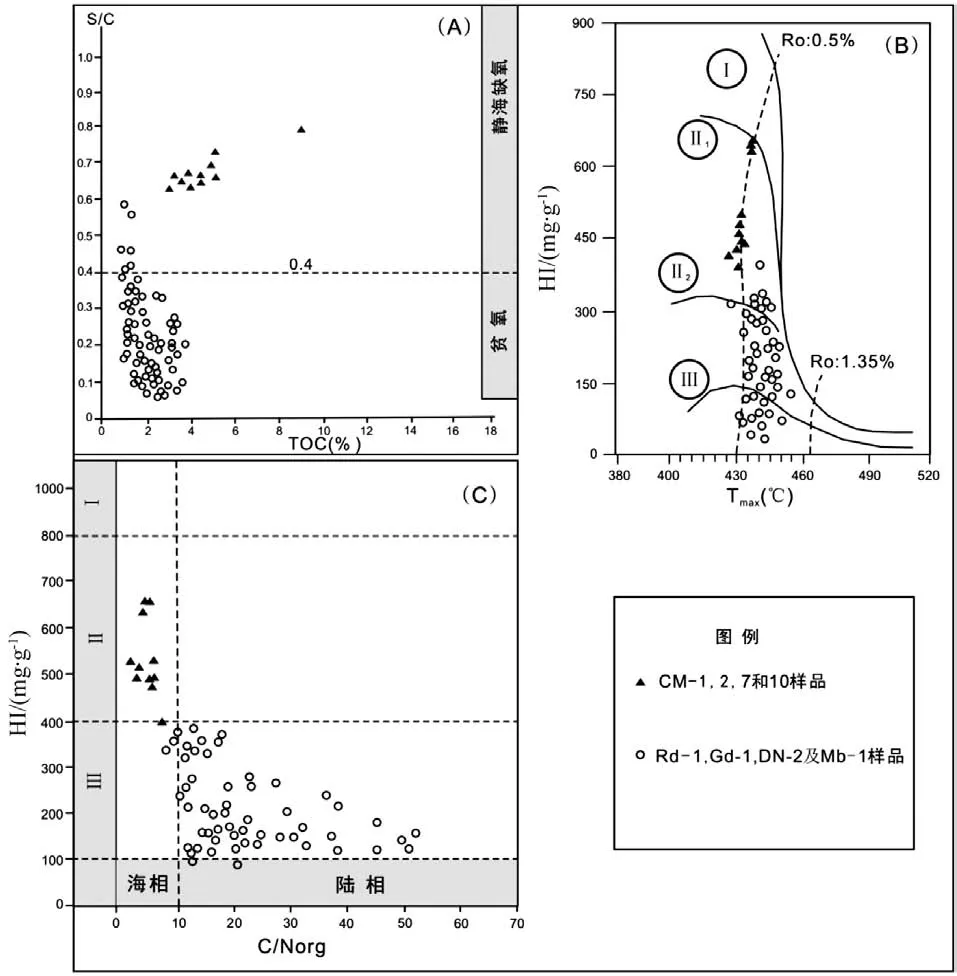
图3 样品的地球化学特征
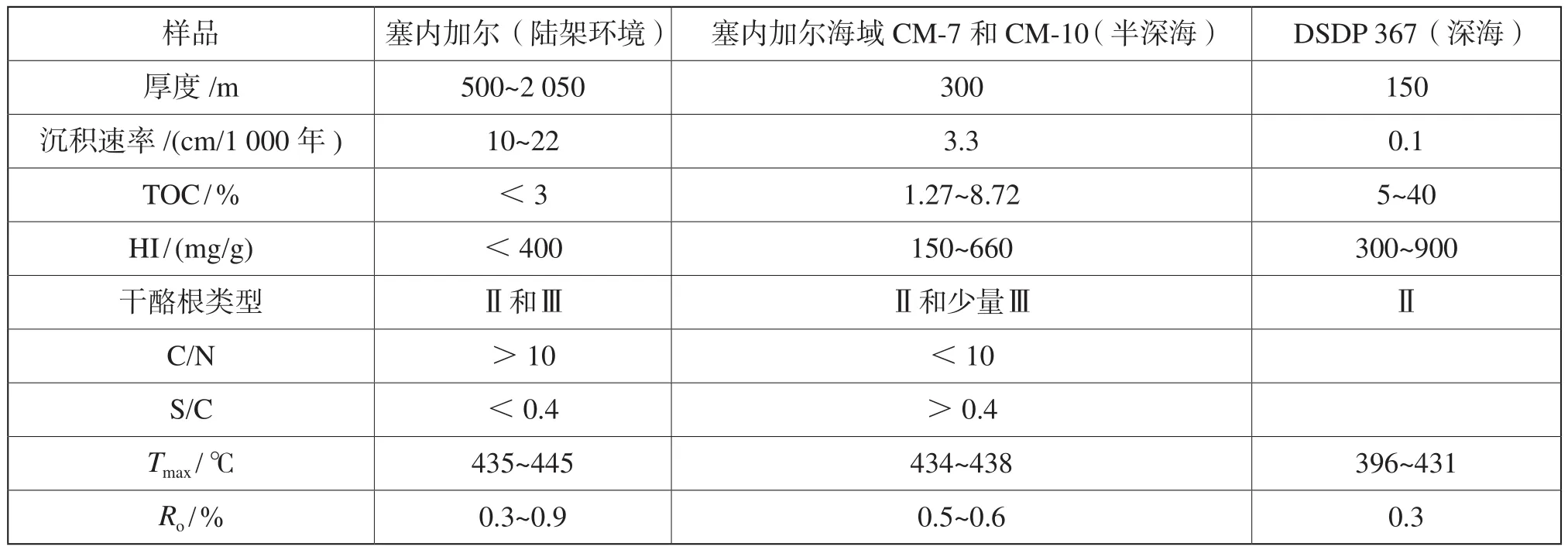
表1 赛诺曼—土伦阶地球化学数据
有机质中C/N值也受控于沉积环境,陆架沉积环境的样品C/N值通常大于10,而半深海沉积环境的样品小于10(图3C),高的C/N值通常指示陆源有机质贡献,因此陆架沉积环境的样品指示了较高的陆源有机质输入。有机质中S/C值可以反映有机质沉积时的氧化还原条件。陆架沉积环境的样品S/C值通常小于0.4(图3A),可能指示了中等氧化条件的沉积环境[16]。半深海环境沉积的样品S/C值基本上大于0.4(图3A),可能暗示了缺氧还原的沉积环境。陆架沉积样品的成熟度处于生油窗阶段,而半深海沉积样品处于低成熟阶段(图3B)。整体而言,所研究样品的成熟度处于未成熟-成熟阶段,这样的成熟度对有机质的影响是有限的,仅当样品处于高成熟-过成熟阶段,才会对有机质丰度、类型产生较大的影响。
2.2 分子地球化学有机质母源特征
热解产物主要化合物的色谱图如图4a所示,主要有烷基苯、正构烷烃以及烷基萘,其中烷基苯略占优势。乙苯是占主导地位的烷基苯化合物,其是主要的链状的脂质体(比如脂肪酸和脂肪醇)环化和进一步芳构化的结果。
陆架及半深海环境沉积样品的正构烷烃分布见图4b。正构烷烃的分布比较有规律,即陆架环境沉积的样品表现为双峰分布样式,前峰为nC13,后峰为nC23,C23–C29无明显的奇偶优势,反映了陆源和海相有机质的混合特征(图4 b1),与上述有机质中C/N值的结果相一致。相反,半深海和深海环境的样品表现为单峰分布,碳数集中在C12~C17 之间(图4 b2,b3),表明有机质主要为海相水生生物来源。
除了烷基苯之外,芳烃馏分中也有烷基萘(图4 a3)和少量烷基菲。陆架环境沉积样品中这些化合物的存在,进一步证实了陆源有机质的输入。半深海样品中鉴定出藿烷类化合物(C27~C31),可能反映了沉积环境中厌氧细菌的发育[17]。
3 晚白垩世赛诺曼—土伦阶古生产力与有机质保存条件分析

图4 热解气相色谱图
古生产力的重建首先要考虑营养源的问题,一般认为海洋表层透光带营养源来自河流的输入或者上升流。前者可以通过沉积物的沉积速率来估算[18]。深海沉积(DSDP 367)C–T黑色页岩的平均沉积速率约为0.1 cm/ky(表1),而陆架环境的沉积速率为10~22 cm/ky(表1),深海沉积速率低而有机质丰度高,陆架环境的沉积速率高而有机质丰度低,因此,这种机制不能解释塞内加尔盆地深海区高有机质丰度优质烃源岩发育的主要成因关系。上升流也可以导致海洋生物生产力的提高。根据Kruijs 和 Barron(1990)[19]的模拟的结果,晚白垩纪赛诺曼—土伦时期塞内加尔海岸的上升流大于20 cm/day(图5)。假定保存因子为2%,该时期北大西洋的初级生产力约为50 g C/(m2·y),远远低于现今上升流发育的地区,其初级生产力高达200~360 g C/(m2·y)[20]。尽管相比与侏罗纪末Kimmeridgian 期(约 20 g C/(m2·y)),生产力得到了一定幅度的提高。由上述讨论可知,晚白垩纪赛诺曼-土伦时期的古生产力条件,不足以形成半深海-深海页岩富含有机质烃源岩,接下来讨论有机质保存条件。
海洋的初级生产力是“供碳”的过程,而塞内加尔盆地晚白垩纪赛诺曼—土伦阶富有机质页岩是“储碳”的结果。供碳的量很大,但是储碳的量却是极小的[21],因此,缺氧条件是有机碳富集的重要因素,甚至是决定性因素。在晚白垩纪赛诺曼—土伦时期,与周围开阔大洋相比,特提斯洋和大西洋显示了缓慢的、迟滞的洋流循环,原因是和周围开阔大洋以窄而浅的水道相连通(图5)。这种条件有助于缺氧底水环境的形成与发展[22],通常这个时期,海洋氧含量最小层带(OMZ)甚至可以扩展至陆架,Thurow和 Kuhnt(1986)[23]估计在北大西洋边缘局部缺氧水体深度可能大于3 000 m。塞内加尔盆地位于北大西洋中段(图1),其所处的地理位置显示了缓慢、迟滞的洋流特征,客观上有助于缺氧水体的形成与发展。
来源于芳基类胡萝卜素的分子化石可以确切地指示古海洋透光带沉积环境。芳基类异戊二烯化合物是光合绿硫细菌成岩作用产物,该细菌需要光和硫化物,因此可以指示静水盆地上部为透光带,下部为缺氧环境。绿硫细菌通过倒三羧酸循环来固定CO2,它们的类胡萝卜素相比于固定CO2轻微亏损13C,因此,它们的分子化石典型的富集13C,相比于藻类来源的有机质,其碳同位素正偏10‰~15‰[24]。DSDP 367黑色页岩鉴定出了芳基类异戊二烯和含硫芳基类异戊二烯化合物(图6)[25],这些化合物来自光合绿硫细菌,这也被同位素所证实,该碳同位素值相比于藻类和蓝藻化石分子正偏10‰~15‰(图6)。塞内加尔盆地晚白垩纪赛诺曼—土伦时期海洋发育静水缺氧环境,芳基类异戊二烯化合物的存在为是一个有力直接的证据。因为缺氧环境的发育,有机质保存条件较好,进而导致有效“储碳”,形成有机质碳富集的优质烃源岩的发育。
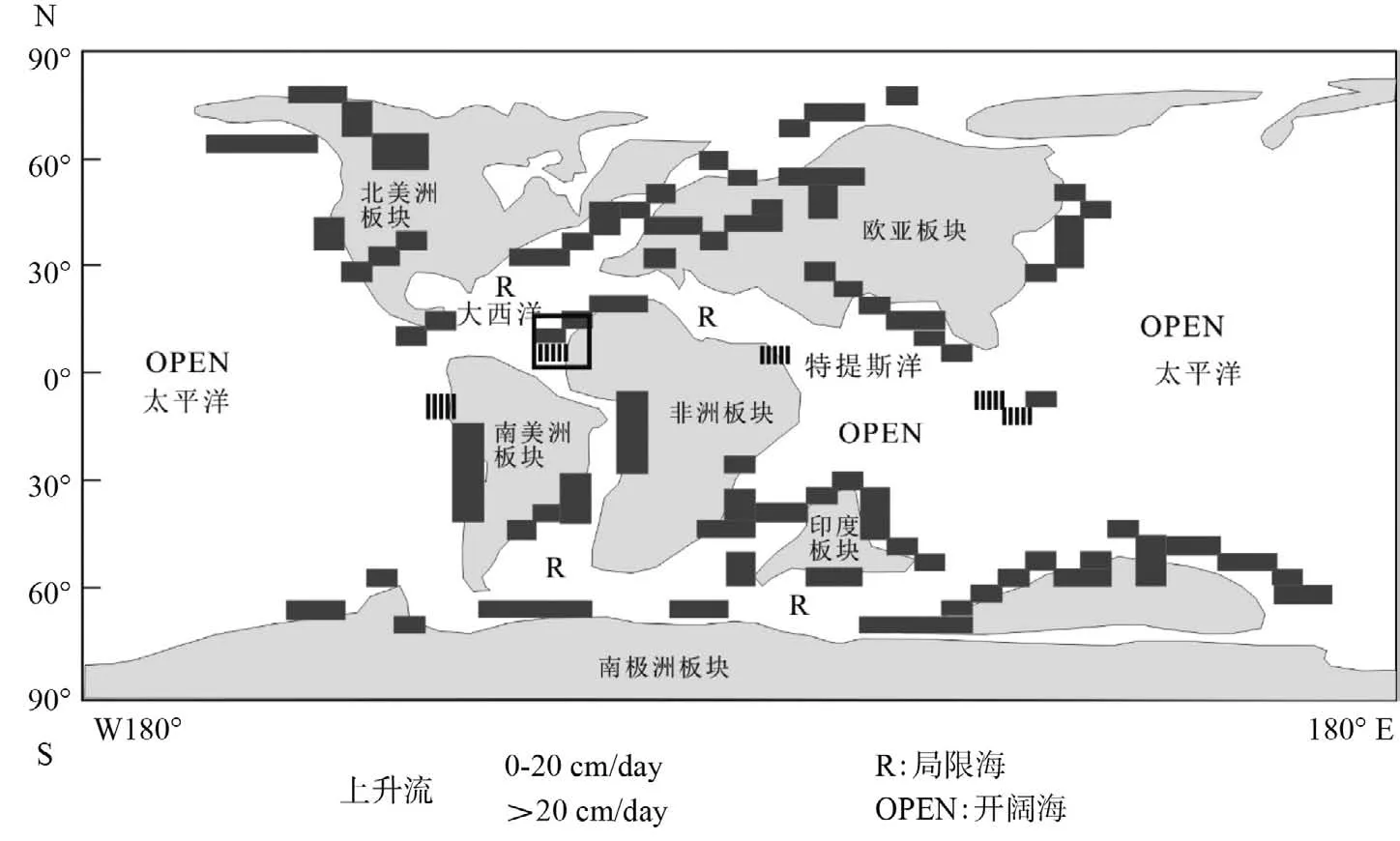
图5 赛诺曼—土伦期上升流模拟 [19]
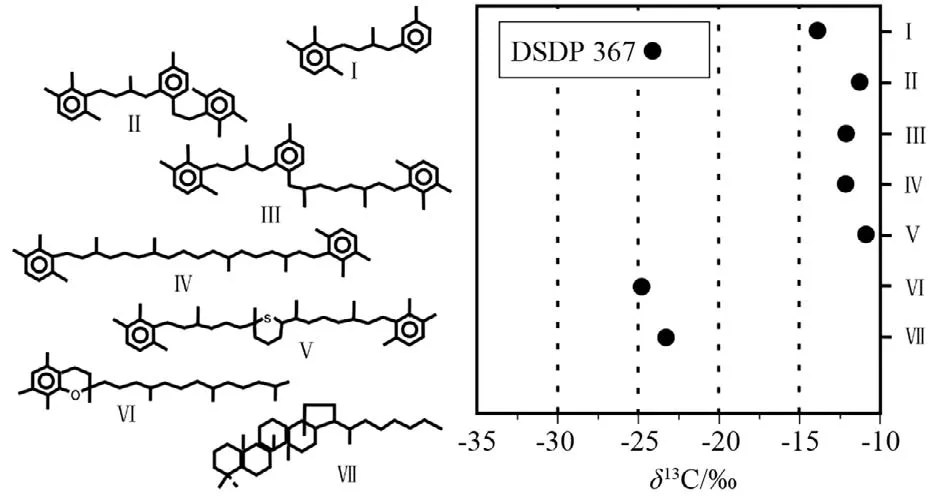
图6 DSDP367黑色页岩芳基类异戊二稀化合物的结构和稳定碳同位素[25]
4 晚白垩世赛诺曼—土伦阶古海洋沉积模式与主控因素
综合上述二类页岩地化特征、生标特征、古生产力和缺氧环境的讨论,建立了塞内加尔盆地晚白垩世赛诺曼—土伦阶时期的沉积和有机相模式(图7)。该模式合理地解释了塞内加尔盆地富有机质页岩的控制因素、分布范围。陆架环境晚白垩纪赛诺曼—土伦阶有机质相对贫瘠,其主要原因是该区的沉积速率较高、水体含氧量较高,有机质保存效率相对较低,储碳结果欠佳,而深海、半深海环境,沉积速率低,海洋氧含量最小层带(OMZ)发育,有机质保存条件好,储碳效率高。因此,缺氧保存条件是海相优质烃源岩的主要控制因素。
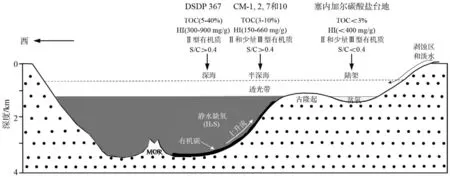
图7 塞内加尔盆地晚白垩世赛诺曼-土伦期古海洋沉积-有机相模式示意图
5 认识与结论
(1)塞内加尔盆地发育两类沉积—有机相:1)陆架浅水碳酸盐台地—较高丰度混合型母质有机相,有机碳含量中等—较高。有机质主要为海生生物和陆源高等植物的混合类型。2)陆坡至深海—高丰度海洋生物母质有机相,具有高的有机质丰度,高的氢指数,为海相优质烃源岩。
(2)塞内加尔盆地晚白垩世赛诺曼—土伦阶海相优质烃源岩的主控因素为缺氧保存条件,其被分子标志物的热解化合物色谱图和芳基类异戊二稀稳定碳同位素所直接证实。同时,该时期相比与晚侏罗世,生产力也得到了一定的提高,这些背景均有利于有机质富集。
[1]童晓光, 关增淼. 世界石油勘探开发图集(非洲地区分册)[M].北京: 石油工业出版社, 2002.
[2]Schlanger S O, Jenkyns H C. Cretaceous Oceanic Anoxic Events: Causes and Consequences[J]. Geologie en Mijnbouw,1976, 55(3-4): 179-184.
[3]Jenkyns H C. Cretaceous Anoxic Events: from Continents to Oceans[J]. Journal of the Geological Society, 1980, 137(2):171-188.
[4]Leckie R M, Bralower T, Cashman R. Oceanic Anoxic Events and Plankton Evolution: Biotic Response to Tectonic Forcing the Mid-Cretaceous[J]. Paleoceanography, 2002, 17(3): 13-1-13-29.
[5]徐汉梁, 单玄龙, 高璇, 等. 西非塞内加尔盆地重油成藏条件及富集规律 [J]. 科学技术与工程, 2014, 14(16): 211-215,220.
[6]Abdelrahman E M, Tealeb A, Hassan R M. A Least-Squares Derivative Analysis of Gravity Data Applied in Senegal, West Africa[J]. Journal of African Earth Sciences, 2001, 33(1):165-175.
[7]郑应钊. 西非海岸盆地带油气地质特质与勘探潜力分析[D].北京: 中国地质大学(北京), 2012.
[8]刘延莉. 塞内加尔盆地油气地质特征及勘探潜力[J]. 地质与资源, 2014, 23(2): 197-201, 196.
[9]熊利平, 刘延莉, 霍红. 西非海岸南、北两段主要含油气盆地油气成藏特征对比[J]. 石油与天然气地质, 2010, 31(4):410-419.
[10]徐志诚, 吕福亮, 范国章, 等. 西非海岸盆地深水区油气地质特征和勘探前景 [J]. 油气地质与采收率, 2012, 19(5): 1-5, 11.
[11]冯国良, 徐志诚, 靳久强, 等. 西非海岸盆地群形成演化及深水油气田发育特征[J]. 海相油气地质, 2012, 17(1): 23-28.
[12]Nzoussi-Mbassania P, Khamlib N, Disnara J R, et al.Cenomanian–Turonian Organic Sedimentation in North-West Africa: A Comparison between the Tarfaya( Morocco) and Senegal Basins[J]. Sedimentary Geology, 2005, 177(3-4):271-295.
[13]Shipboard Scientific Party, Site 367 : Cape Verde Basin, Init.Rep. DSDP 41, 163-232.
[14]Espitalie' J, Le Laporte J, Madec M, et al. Me' thode rapide de caracte' risation des roches me`res de leur potentiel pe'trolier et de leur degre' d’e'volution[J]. Revue de l'Institut Francais du Petrole, 1977, 32 : 23-42.
[15]Tissot B P, Welte D H. Petroleum Formation and Occurrence[M].Berlin : Springer-Verlag, 1984.
[16]Morse W J, Berner A R. What Determines Sedimentary C/S Ratio[J]. Geochimica et Cosmochimica Acta, 1995, 59(6):1073-1077.
[17]Peters E K, Moldowan J M. The Biomarker Guide: Interpreting Molecular Fossils in Petroleum and Ancient Sediment[M].Englewood Cliffs, NJ: Prentice Hall, 1993.
[18]Waples D W. Reappraisal of Anoxia and Organic Richness, with Emphasis on Cretaceous of North Atlantic[J]. AAPG Bulletin,1983, 67(6): 963-978.
[19]Kruijs E, Barron E. Climate model prediction of paleoproductivity and potential source-rock distribution[C]//Huc A Y, Ed.Deposition of Organic Facies. Tulsa, OK : AAPG, 1990, 30 :195-216.
[20]Pedersen T F, Calvert S E. Anoxia vs. Productivity : What Controls the Formation of Organic-Carbon-Rich Sediments and Sedimentary Rocks?[J]. AAPG Bulletin, 1990, 74(4): 454-466.
[21]焦念志. 海洋固碳与储碳—并论微型生物在其中的重要作用[J]. 中国科学 : 地球科学, 2012, 42(10): 1473-1486.
[22]Philip J, Babinot J F, Tronchetti G, et al. Late Cenomanian[C]//Dercourt J, Ricou L E, Vrielynck B, Eds. Atlas Peri-Tethys,Palaeogeographical Maps. Paris: CCGM/CGMW, 2000.
[23]Thurow J, Kuhnt W. Mid-Cretaceous of the Gibraltar Arch Area[C]//Summerhayes C P, Shackleton N J, eds. North American Palaeoceanography. Geological Society, London,Special Publications, 1986, 21 : 423-445.
[24]Koopmans M P, Köster J, Van Kaam-Peters H M E, et al.Diagenetic and Catagenetic Products of Isorenieratene:Molecular Indicators for Photic Zone Anoxia[J]. Geochimica et Cosmochimica Acta, 1996, 60(22): 4467-4496.
[25]Damsté J S S, Köster J. A Euxinic Southern North Atlantic Ocean During the Cenomanian/Turonian Oceanic Anoxic Event[J]. Earth and Planetary Science Letters, 1998, 158(3-4): 165-173.
