不同品种丰花月季根系抗寒性比较
2018-01-12焦美玲弓瑞娟范少然崔睿航武东霞
焦美玲, 张 钢, 孟 昱, 弓瑞娟, 范少然, 崔睿航, 武东霞
(1.河北农业大学园艺学院,河北保定 071001; 2.河北农业大学园林与旅游学院,河北保定 071001)
木本植物在其发育的年生长周期期间,根系暴露于许多胁迫因子下。在我国北方地区霜冻是常见的逆境之一,最冷月空气温度可能低于-40 ℃,土壤冻结,为了生存,根系需要进行低温锻炼。近年来,电阻抗图谱(electrical impedance spectroscopy,EIS)技术被广泛用于研究不同器官对非生物胁迫的生理反应,包括冷驯化[1-2]、重金属胁迫[3]、涝渍胁迫[4]、盐分敏感度[5]等方面,具有广泛的应用前景。本研究对根系进行人工冷冻处理,研究电阻抗图谱参数结合Logistic方程评价不同月季品种根系的抗寒性,并与传统的电导法进行比较,结合相对含水量、可溶性糖含量、淀粉含量指标综合分析、比较品种间的抗寒性差异,以期为月季品种的抗性育种、区域化栽培、推广应用提供理论基础。
1 材料与方法
1.1 试验材料
供试材料为河北农业大学标本园(115°26′E,38°50′N)内多年生丰花月季品种红帽、 金玛丽、柔情似水、仙境,栽培条件一致,常规养护管理。分别于2015年12月、2016年2月、2016年3月取样,每个品种选取生长状况良好的月季6株(每小区2株),全株挖出,与土壤分离,带回。剪下的根用自来水充分淋洗以除去土壤和根表死皮,并剪除须根[6],选取粗细均匀、无病虫害、无机械损伤的粗度为2.0~5.0 mm的根段,再用去离子水清洗3遍,吸水纸吸干表面水分备用。
1.2 指标测定(处理方法)
将根系进行人工冷冻处理,以4 ℃为对照,设置6个不同的冷冻温度,每次取样处理温度均为4(对照)、-2、-4、-6、-8、-12、-16 ℃,降温速率为4 ℃/h,达到指定温度保持 4 h,在处理完毕的枝条再升温到4 ℃(升温速率同降温速率),完全解冻后2 h后即可用于电阻抗及相关生理指标的测定。
1.2.1 电阻抗图谱 每个处理中随机选取6段15 mm长的根系样本用作电阻抗测定。参考前人方法[7],用测厚仪(Mitutoyo No.7331,Japan)测定直径,精确到0.01 mm,用阻抗仪(Agilent E4980A,USA)测定样本在42个频率(80 Hz~1 MHz)下的电阻值和容抗值,作出不同频率下电阻和容抗的变化曲线,即EIS。月季根系的EIS为单弧,采用分布模型中的单-DCE模型。拟合的参数为电阻R(Ω)和R1(Ω)、弛豫时间τ、弛豫时间分布系数ψ;需计算的为胞外电阻率re(Ω·m)、胞内电阻率ri(Ω·m)、低频电阻率r1(Ω·m)、高频电阻率r(Ω·m)。
1.2.2 相对电解质渗出率 参照前人方法[8],将每个处理温度下根系切取10 mm放入试管,加12 mL去离子水,用保鲜膜将试管口封好,试验设4次重复。试管放入摇床中振荡 24 h。用BANTE-950型电导仪(上海般特仪器有限公司)测定初电导值C1和空白对照C空白1,并记录;C1测定完毕后,再用保鲜膜封口,将试管置于沸水中水浴20 min,自然冷却后再放入摇床中振荡24 h,测终电导值C2和C空白2。通过公式(1)计算相对电导率(REL):

(1)
1.2.3 干物质含量 与EIS测定同步进行。每个品种对照组随机选取6段15 mm长的根段,然后放入烘箱105 ℃杀青30 min,置于80 ℃下烘干至恒质量,计算相对干物质含量(relative dry matter content,RDMC):
RDMC= (干质量/鲜质量)×100%。
(2)
1.2.4 可溶性糖含量和淀粉含量的测定 采用蒽酮比色法[9]将根系置于80 ℃烘箱48 h烘干,取出后放于干燥皿里待用。每个月季品种设3次重复。
取磨成粉末的根系样品0.2 g置于10 mL离心管中,用80%乙醇80 ℃水浴浸提,分设3次离心提取,将3次提取的上清液合并于25 mL容量瓶,用80%乙醇定容。准确吸取0.8 mL溶液于10 mL离心管水浴蒸干,加入5 mL水充分搅拌,使糖完全溶解,离心备用。
向沉淀中加水2 mL,搅拌均匀,在沸水浴中糊化15 min。冷却后,冰浴中加入9.2 mol/L高氯酸2 mL,搅拌15 min后加水 2 mL 混匀,离心10 min,上清液倾入50 mL容量瓶,再向沉淀中加入4.6 mol/L高氯酸2 mL,搅拌15 min后加水3 mL混匀,离心10 min,然后用水洗(4 mL)沉淀1次,合并各次离心的上清液于上述50 mL容量瓶中,用蒸馏水定容至刻度,备用。
向干净的试管中加入1 mL水,再加入1 mL上述提取液(调零对照加入2 mL水),冷水中沿壁缓慢加入5 mL硫酸蒽酮混匀,80 ℃水浴中显色10 min,于620 nm下测定吸光度D值。计算可溶性糖含量和淀粉含量。
1.3 统计分析
用Microsoft Excel 2003计算出REL,并将REL所得数据随温度变化用Microsoft Excel 2003作图,参照Logistic方程,见式(3),用SPSS 17.0软件计算半致死温度,表征月季根系的抗寒性,给出与Logistic方程的决定系数r2和线性相关系数r。通过LEVM 8.06软件[10]拟合得出EIS参数值。同样EIS参数与温度参照方程(3)拟合得到半致死温度,统计EIS各参数(胞外电阻率re、胞内电阻率ri、弛豫时间τ、弛豫时间分布系数ψ)及生理指标(相对干物质含量、可溶性糖含量及淀粉含量)的变化,做相关分析,EIS各参数的解释参见文献[11]。
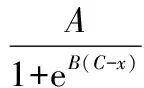
(3)
式中:x为冷冻处理温度(℃);y为各EIS参数(re、ri、τ、ψ)或相对电导率(%);A+D表示未受冻害时EIS各参数的基础值或受冻害时REL的最大值;B为EIS各参数或REL随冷冻温度变化的斜率(%/℃);C为曲线拐点对应温度(℃),即样本的抗寒性,以LT50表示;D表示受冻害后EIS各参数的最小值或未受冻害时REL的基础值。
2 结果与分析
2.1 土壤霜冻期(2月)EIS的变化
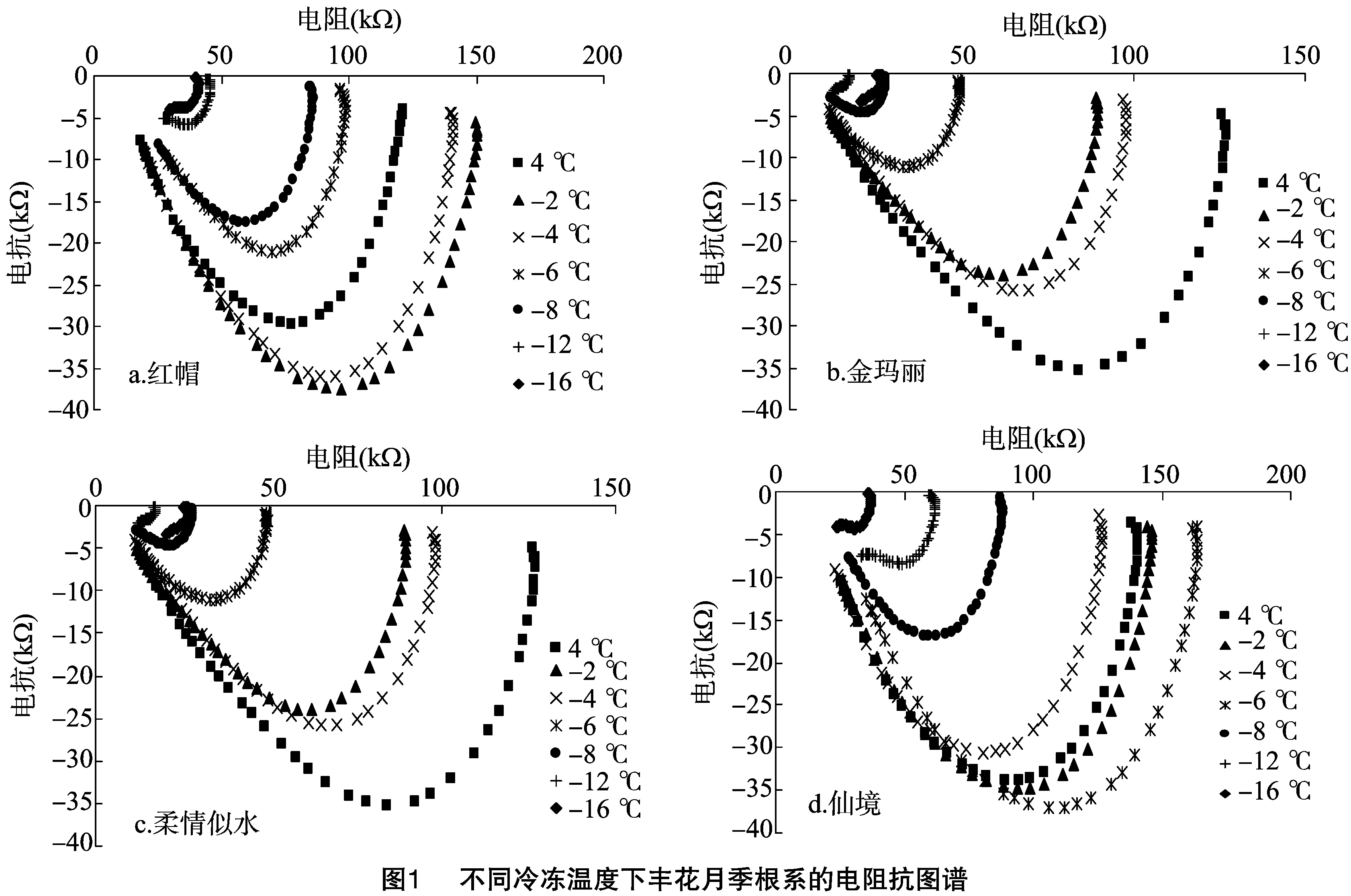
研究发现,4种丰花月季根系的电阻抗图谱均为单弧,当冷冻处理温度降至足够低时,单弧明显变小,且有变形(变散)的趋势。其中,金玛丽和柔情似水变化趋势较为一致,随着胁迫温度的降低,弧度不断变小,而红帽和仙境则分别是 -2、-4 ℃弧较对照(4 ℃)大,而后又变小。当弧顶值达到 -15 kΩ 左右时,4种月季的EIS弧度明显小于各自对照的弧度,可以判定月季根系组织受害,将要或已经达到半致死温度(图1)。
2.2 电导法确定半致死温度
研究发现,4个品种丰花月季根系的低温致死曲线都可以和Logistic方程得到很好的拟合,拟合优度r2均达到0.80以上,且土壤霜冻期及土壤结冻前均为仙境根系抗寒性最强,最强时可以达到-10.66 ℃,根系抗寒性顺序为仙境>红帽>柔情似水>金玛丽。而初春土壤解冻后,各品种之间的抗寒性差异不大,但顺序有所变化,红帽最强,而柔情似水最弱(表1)。
2.3 EIS参数计算半致死温度
研究发现,几乎4个EIS参数(红帽3月ψ未估计出)都可以估计出月季根系的抗寒性。re与REL所得抗寒性结果最为接近,基本都是(红帽3月抗寒性较强)土壤霜冻状态下抗寒性最强,2种方法所得抗寒性值相差1~3 ℃。而ri较REL估计的抗寒性普遍偏高,与REL所得抗寒性同样很接近,但抗寒性结果有所不同,金玛丽2月抗寒性最弱(表2)。4个品种τ求得的抗寒性与其他方法相差较大。各参数与REL法测定月季根系抗寒性的相关分析结果显示出较高的相关性,其中各个品种re法与REL法相关性r2均达到0.62以上,柔情似水的4个参数的r2均达到0.9以上,显示出明显的线性相关性(表3)。
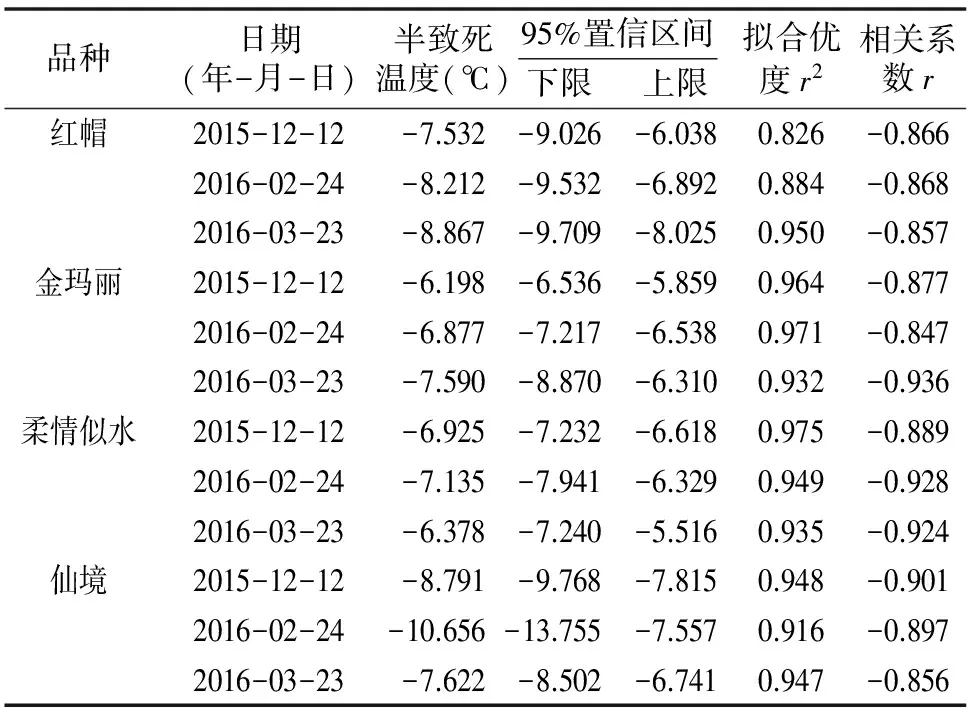
表1 4个丰花月季品种根系半致死温度变化

表2 4个丰花月季品种EIS各参数计算出的半致死温度
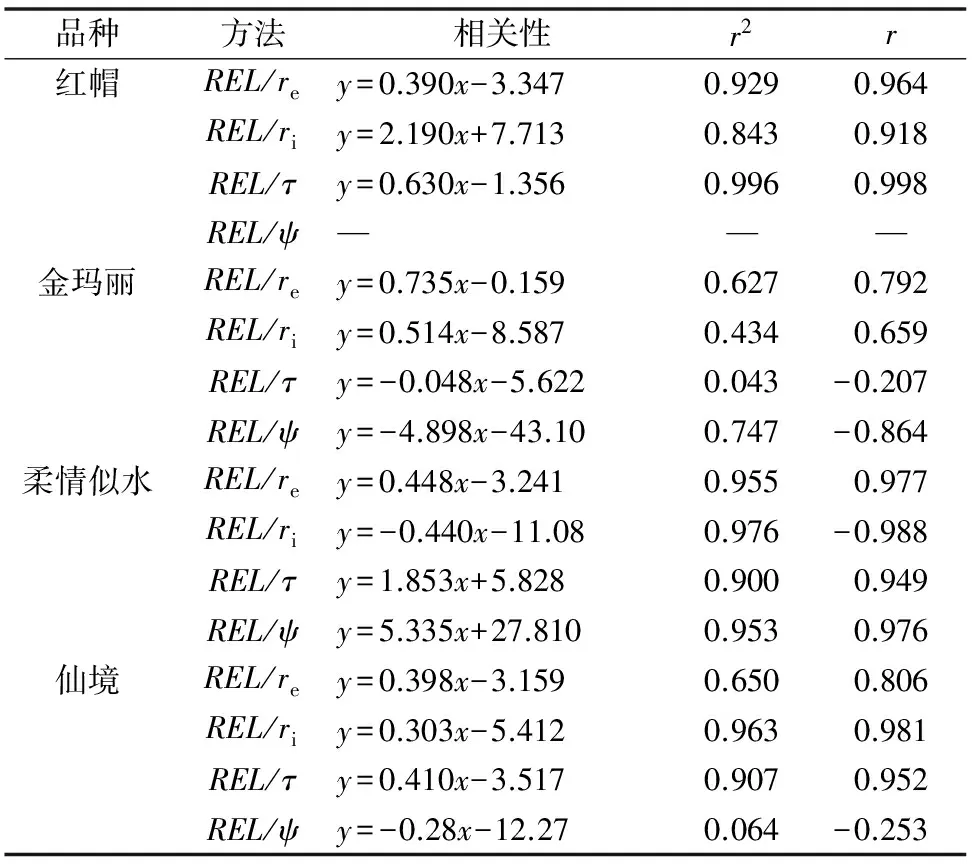
表3 REL与EIS各参数测定的丰花月季根系抗寒性的相关性
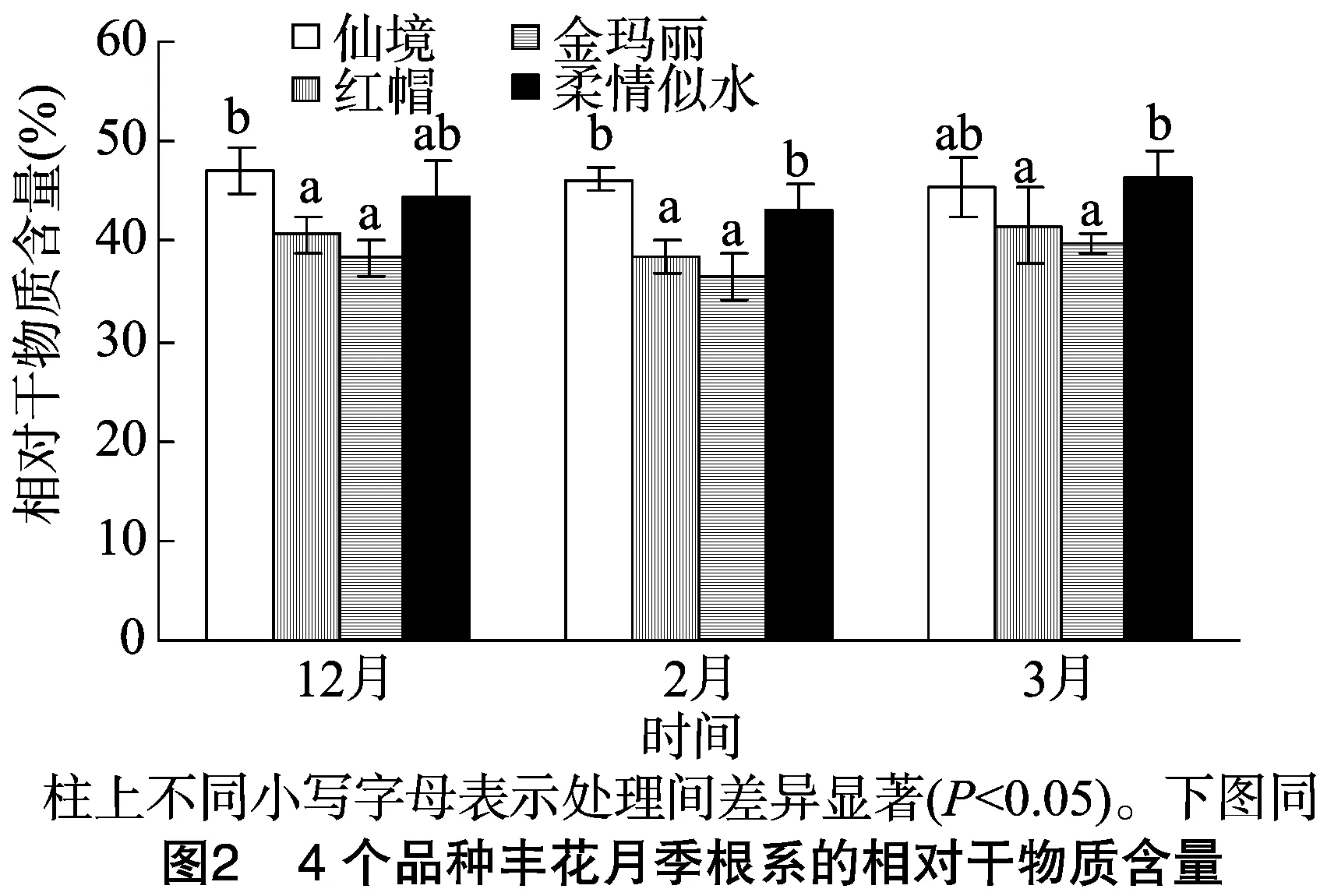
2.4 相对干物质含量
研究发现,金玛丽和红帽根系的相对干物质含量(RDMC)明显低于柔情似水和仙境,不同时期抗寒性强的仙境的RDMC最高值出现在12月,高达46.94%,而金玛丽最低出现在2月,为36.58%,抗寒性最弱。红帽和柔情似水的RDMC与其抗寒性结果不一致,但柔情似水的RDMC与其抗寒性有着良好的线性相关性,r2高达0.98(其他品种r2<0.2)(图2)。
2.5 可溶性糖含量和淀粉含量
研究发现,4个月季品种根系的可溶性糖含量都是在2月最高,而土壤结冻前的12月和土壤解冻后的3月,其含量明显低于抗寒性较强的2月,其中2月金玛丽根系可溶性糖含量显著低于其他3个品种月季(P<0.05),而仙境、红帽和柔情似水之间差异不显著,但仙境的可溶性糖含量最高,与其抗寒性结果一致。淀粉含量浮动在1%~2.5%,各品种之间没有明显差异,但总体上看,土壤霜冻时期淀粉含量要低于其他2个时期(图3)。
3 讨论
朱根海等认为可以用Logistic方程中拐点温度来确定植物组织受低温胁迫的半致死温度(LT50)[12]。低温胁迫可以使生物膜发生由液晶相向凝胶相的变化,膜流动性降低,通透性增加,细胞膜透性的改变可以用来表征其结构和功能的受损程度[13]。本研究结果表明,相对电导率及部分EIS参数可以和Logistic方程得到很好的拟合,re、τ、ψ与温度呈现线性正相关,所得的半致死温度在-5.111~-13.911 ℃,而REL和ri同样遵循“S”形曲线变化,与温度呈负相关关系。其中re所求半致死温度估计的抗寒性更是与传统的相对电导率所得的抗寒性结果完全一致,但是比REL法低0.723~3.054 ℃(2月),根系抗寒性最强的仙境,二者所得抗寒性相差最大,这与Repo等对欧洲赤松(PinussylvestrisL.)、李亚青等[14],董胜豪等对白皮松(P.bungeanaZucc.)在抗寒性较强时所得结果[15-16]一致。说明用EIS参数来判定丰花月季根系的抗寒性相对可靠。
几个月季根系的电阻抗图谱均为单弧,与孟昱等在白桦(BetulaplatyphyllaSuk.)根系上的研究一致,EIS弧的大小变化表明,根系组织的结构及生理生化状态在温度低于某一数值时发生改变,遭受冻害以后,弧顶值(绝对值)呈减小趋势(图1),温度很低时,电阻值和容抗值(绝对值)变得很小且图谱变散[4]。

植物的含水量与其物质代谢有着密切的关系[17-18],细胞水分含量是影响抗寒性的重要因素之一,金研铭等研究得出牡丹(PaeoniasuffruticosaAndr.)束缚水自由水比值大,抗寒性强的结论[19]。束缚水含量高而自由水含量相对较低即相对干物质含量高,则抗寒性强。本研究中,月季根系相对干物质含量与半致死温度有一定的线性相关关系,抗寒性强的品种仙境干物质含量高于其他品种。
糖是植物抗寒的主要保护物质,植物体内糖含量高,可以提高原生质浓度,降低细胞液冰点,亦可缓和细胞质过度脱水,保护细胞质胶体不致遇冷凝固[20]。根系组织中积累可溶性糖,维持膜在低温下的正常功能,而淀粉含量与植物抗寒性同样有密切关系,但不直接参与渗透调节,主要通过降解成可溶性糖等物质来间接影响抗寒性。申晓慧等对紫花苜蓿(MedicagosativaL.)的研究表明,抗寒性强的品种可溶性糖含量在越冬最冷月较高[21]。刘慧民等对绣线菊(SpiraeasalicifoliaL.)的研究也表明可溶性糖含量在众多指标中与抗寒性关联度最大[22]。本研究中月季根系的可溶性糖含量与抗寒性有正相关关系,土壤霜冻期可溶性糖含量高而淀粉含量低。
丰花月季根系处于土壤霜冻状态的外环境时,通过调节自身水分含量维持低温状态下细胞正常的代谢,同时通过淀粉、蛋白质等大分子物质降解为可溶性糖等物质来调节细胞渗透压,从而提高根系的抗寒性。当初春气温回升,月季抗寒性相应的减弱,细根明显增多,可溶性糖、淀粉、水含量等相应的动态调整,将要或已经为地上部生长做准备。
[1]张海旺,陈海江,张学英,等. 桃树脱锻炼期间电阻抗参数与抗寒性关系研究[J]. 北方园艺,2014(6):11-14.
[2]邸 葆,孟 昱,张 钢,等. 基于电阻抗图谱估测金丝楸茎和叶可溶性糖含量的模型[J]. 生物物理学报,2014,30(6):443-453.
[3]向地英,邸 葆,张 钢,等. 镉胁迫对黄栌叶片电阻抗参数及相对电导率的影响[J]. 河北农业大学学报,2014,37(2):58-64.
[4]孟 昱,弓瑞娟,邸 葆,等. 涝胁迫下白桦叶和茎含水量的电阻抗图谱估测[J]. 植物学报,2014,49(1):105-113.
[5]姚 琳,张 钢,吕 薇,等. 盐碱地区不同观赏树种电阻抗图谱参数和电导率的变化[J]. 中国农学通报,2011,27(6):54-59.
[6]袁军伟,郭紫娟,刘长江,等. 11个葡萄品种根系抗寒性的综合评价[J]. 中外葡萄与葡萄酒,2015(2):21-25.
[7]Zhang G,Ryyppö A,Repo T. The electrical impedance spectroscopy of Scots pine needles during cold acclimation[J]. Physiologia Plantarum,2002,115(3):385-392.
[8]王刘环,弓瑞娟,张 钢. 抗寒锻炼期间丰花月季的电阻抗参数与抗寒性的关系[J]. 河北农业大学学报,2014,38(1):48-52.
[9]高俊凤. 植物生理学实验指导[M]. 北京:高等教育出版社,2006:144-147.
[10]Repo T,Zhang M,Ryyppö A,et al. Effects of freeze-thaw injury on parameters of distributed electrical circuits of stems and needles of Scots pine seedlings at different stages of acclimation[J]. Journal of Experimental Botany,1994,45(275):823-833.
[11]Repo T,Lappi J. Estimation of standard error of impedance-estimated frost resistance[J]. Scandinavian Journal of Forest Research,1989,4(1/2/3/4):67-74.
[12]朱根海,刘祖祺,朱培仁. 应用Logistic方程确定植物组织低温半致死温度的研究[J]. 南京农业大学学报,1986(3):11-16.
[13]Lyons J M,Wheaton T A,Pratt H K. Relationship between the physical nature of mitochondrial membranes and chilling sensitivity in plants[J]. Plant Physiology,1964,39(2):262-268.
[14]Repo T,Zhang G,Ryyppö A,et al. The relation between growth cessation and frost hardening in Scots pines of different origins[J]. Trees,2000,14(8):456-464.
[15]李亚青,张 钢,卻书鹏,等. 白皮松茎和针叶的电阻抗参数与抗寒性的相关性[J]. 林业科学,2008,44(4):28-34.
[16]董胜豪,张 钢,卻书鹏,等. 脱锻炼期间白皮松针叶的电阻抗图谱参数和生理指标的相关性[J]. 园艺学报,2009,36(6):891-897.
[17]艾育芳,陈观水,钟陈菲,等. 干旱胁迫对巨菌草幼苗水分状况和渗透调节物质的影响[J]. 江苏农业科学,2015,43(5):344-347.
[18]佘波.旱柳花期枝条内养分含量的动态变化[J]. 江苏农业科学,2015,43(5):190-192.
[19]金研铭,徐惠风,李亚东,等. 牡丹引种及其抗寒性的研究[J]. 吉林农业大学学报,1999,21(2):37-39.
[20]潘瑞炽,董愚得. 植物生理学[M]. 北京:高等教育出版社,1958:322-328.
[21]申晓慧,姜 成,冯 鹏,等. 6种紫花苜蓿越冬前后几个抗寒生理指标变化研究[J]. 农学学报,2015,5(12):94-98.
[22]刘慧民,仉 茜,苏 青,等. 18种绣线菊苗期抗寒性评价与筛选[J]. 园艺学报,2014,41(12):2427-2436.
