新老玉米品种花粒期干物质及氮素积累转运机制的差异分析
2018-01-12张中东郭正宇陈永欣
张中东,郭正宇,宫 帅,陈永欣
(山西省农业科学院 玉米研究所,山西 忻州 034000)
玉米花后灌浆期是籽粒建成、充实的关键产量形成时期[1-7],玉米花后的灌浆过程伴随着植株自身的干物质积累和氮素吸收,以及各营养器官干物质和氮素向籽粒的转运[1-2],且这2个过程存在明显的相互制衡作用[1-4]。国内外很多学者从玉米地上部各器官、地下部根系以及花前花后氮素和干物质积累、分配和再转运过程等角度,对玉米植株和籽粒氮素营养及物质积累生理特征与机制进行了系统解析[5-16]。在多年来多次的主栽品种更替和发展过程中,玉米品种自身的干物质及氮素营养策略和生理特性是否已经随之发生变化,如何变化,尚需进一步探明和系统解析。针对新老玉米的氮素积累生理机制差异研究,前人已做了很多工作。Ciampitti等[9]研究认为,花后更高的氮素吸收是现代品种具有更高干物质积累和籽粒所含氮素的主要原因,其中,新品种籽粒所含的氮素中56%来自花后氮素吸收,明显大于老品种(老品种约50%),而二者氮收获指数没有差异;Gallais等[14]研究认为,新品种有更多的花前氮素积累量,以为灌浆前期的高强度氮需求提供保障;Below等[15]研究认为,较老的品种氮素转运启动开始约在吐丝期开始到吐丝后7 d,早于较新品种;Ciampitti等[12]研究认为,相比老品种,新品种氮素转运的启动时间延后到灌浆中期,延后的氮素转运有利于植株光合能力的高保有量,更能获得高产。Tsai等[17]在低氮条件下研究籽粒氮素来源,结果认为,来自转运氮素的贡献高于吸收。而Fonzo等[18-20]研究则认为,在低氮条件下籽粒氮素组成、植株转运氮素和花后吸收氮素的贡献大约相等。Pan等[21-25]和Ciampitti等[10]研究认为,在高密度条件下转运氮素对于籽粒氮素积累的贡献要比花后氮素吸收高很多。上述研究分析了新老品种在花前花后转运氮素和吸收氮素对籽粒氮素的贡献,以及从玉米营养器官氮素转运的启动时间、低氮、高密度条件方面分别比对新老品种氮素营养机制的差异,但具体到玉米各个器官花前干物质和氮素积累分别对花后这些器官干物质和氮素转运分配影响的生理机制尚不清楚,且在不同氮素和密度条件下,新老玉米氮素营养策略和调整机制差异也尚无明确的结论。
本研究以典型现代品种先玉335、郑单958和典型老品种中单2号为材料,在不同氮素和密度水平条件下系统分析新老品种各器官干物质及氮素的积累和分配差异,明确新老玉米品种在氮素营养策略和调整机制方面的差异,旨在为玉米高产氮高效栽培调控和育种方向利用提供理论依据。
1 材料和方法
1.1 试验地概况
试验于2014-2015年在国家玉米产业技术体系忻州综合试验站(38°42′N,112°67′E)进行。前茬作物为玉米,土壤类型为浅褐土,质地为壤性,0~20 cm土层有机质含量8.22 g/kg,速效氮41 mg/kg,速效钾137.2 mg/kg,速效磷11.4 mg/kg,pH值8.14。2014,2015年玉米全生育期降雨量分别为445,432 mm,≥10 ℃土壤活动积温分别为3 450,3 522 ℃(图1)。
1.2 试验材料
以我国20世纪70年代主推杂交品种中单2号(ZD2)和21世纪前10年的主推杂交品种先玉335 (XY335)和郑单958(ZD958)为材料。
1.3 试验设计
试验设2个氮处理,追氮100 kg/hm2(100N) 和追氮200 kg/hm2(200N),于拔节期追施;设3个密度处理,5.25万(D1),7.50万(D2),10.50万株/ hm2(D3),底肥统一施用105 kg/hm2P2O5(磷酸二铵)和45 kg/hm2K2O(硫酸钾),整地时均匀旋耕入土。试验采用随机区组设计,3次重复,行距0.6 m。2014年播种期为5月1日,收获期为10月2日;2015年播种期为4月29日,收获期为10月1日。
1.4 测定指标及方法
以小区内50%植株达到某生育时期为准,准确记录各试验小区生育期到达时间。于吐丝期和生理成熟期选择长势整齐一致的连续3株,测定玉米植株及各器官干物质积累量(DM),吐丝期将植株分为叶片(Leaf)、茎秆(茎、叶鞘、雄穗)(Stem)和雌穗(Ear-shoot),生理成熟期将植株分为叶片(Leaf)、茎秆(茎、叶鞘、苞叶)(Stem)、穗轴(Cob)和籽粒(Grain),将植株切成5~7 cm样段装袋,于65 ℃烘箱内烘干至恒重,称量并记录干质量。将烘干后的样品粉碎过1 mm筛后,采用凯氏定氮法测定各器官含氮量。于生理成熟期,选无缺苗断垄且长势整齐的2行,记录总株数、双穗数、空秆数以及丝黑穗病情况;实收2行所有果穗,待果穗风干后考种,逐穗测定穗粒数后全部脱粒,测定含水量、千粒质量、收获指数,并计算产量。

图1 2014-2015年玉米生育期内日均降雨量、累积降雨量和日均温度Fig.1 Total daily precipitation, cumulative precipitation and daily average temperature during the whole growth period of maize in 2014-2015
1.5 数据分析
采用Microsoft Excel 2010软件进行数据处理,采用SPSS 17.0软件进行方差分析,采用Sigmaplot 10.0软件进行作图。
籽粒氮素含量=籽粒氮素积累量/籽粒干物质积累量×100%;叶片氮素含量=叶片氮素积累量/叶片干物质积累量×100%;营养器官氮素转移量=吐丝期营养器官氮积累量(NUs)-成熟期营养器官氮积累量;收获指数= 成熟期籽粒干物质积累量/成熟期植株干物质积累量;氮收获指数= 成熟期籽粒氮积累量/成熟期植株氮积累量;花后干物质积累量=成熟期干物质量-吐丝期干物质量;花后氮素积累量=成熟期氮素积累量-吐丝期氮素积累量。
2 结果与分析
2.1 新老品种花前干物质及氮素积累差异
由表1-4可知,在2年的试验中,新老品种干物质积累及氮积累指标,在各处理之间双因素和三因素互作并不显著(仅2014年成熟期穗轴干物质量(Cob DM)和吐丝期茎干物质量(Stem DM)D×V互作效应显著,由此可知,各干物质积累及氮积累指标均稳定的受到来自品种、氮肥和密度处理的独立影响,互作影响并不明显。
由表1,2可知,新品种(XY335和ZD958)的吐丝期全株干物质积累量大于老品种,其中,2014,2015年XY335较ZD2分别高0.6,0.7 t/hm2,而ZD958较ZD2在2014年高0.3 t/hm2,但在2015年仅高0.1 t/hm2,且二者差异不显著(表2)。从不同器官角度看,新品种吐丝期叶片干物质积累量总体高于老品种,2015年XY335和ZD958吐丝期叶片干物质积累量均显著高于ZD2,平均高0.11 t/hm2,但在2014年新老品种之间叶片干物质积累量的差异不显著(表1,2)。同时,新品种吐丝期茎秆干物质积累量总体较老品种低,具体表现为2014年XY335和ZD958较ZD2平均低1.18 t/hm2,2015年ZD2较ZD958高0.48 t/hm2,而XY335与ZD2的茎秆干物质积累量的差异不显著(表1,2)。新品种花前干物质积累较老品种更具优势,综合2年数据,玉米新品种花前干物质较老品种平均高0.43 t/hm2,同时新品种吐丝期茎秆生物量与叶片生物量的比值比老品种显著小11.7%,说明这种优势源于叶片的干物质积累优势。综合2年的结果看,无论是新老品种,还是不同水平的氮素和密度处理下,吐丝期玉米茎秆干物质始终是叶片干物质(Leaf DMs)1.9~2.6倍。而随着N水平的提高,玉米叶片干物质积累量和茎秆干物质积累量均无明显的上升,从全株的干物质积累看,2014年高氮条件下(200N)较100N明显高0.72 t/hm2,而在2015年2种N水平下全株的干物质积累的差异并不显著(表1,2)。随着密度(D)水平的增加,2年结果均表现出成熟期玉米全株干物质积累呈显著上升趋势,2年平均数据成熟期全株干物质积累量在高密度(D3)下分别较中(D2)、低(D1)密度提高0.54,1.73 t/hm2。
由表3,4可知,新品种(XY335和ZD958)的吐丝期叶片氮素浓度显著小于老品种(ZD2),2014年ZD2较XY335和ZD958吐丝期叶片氮素浓度分别高1.3,0.5 mg/g,2015年分别高0.8,0.9 mg/g。但是从全株的吐丝期氮素积累看,新品种较老品种高,2年平均高8.6 kg/hm2,而且新老品种间,吐丝期茎秆和叶片的氮素积累情况并不相同,从茎秆角度看,2年均表现出老品种(ZD2)吐丝期茎秆氮素积累显著高于XY335和ZD958,2014,2015年分别平均高8.95,3.35 kg/hm2。从叶片角度看,2014年新老品种的叶片氮素积累差异不明显,但在2015年XY335和ZD958叶片氮素积累均显著高于ZD2,平均高出3.45 kg/hm2(表3,4)。新老品种花前氮积累策略不同,新品种更侧重于花前叶片氮积累,而老品种侧重花前茎秆氮积累。综合2年的结果看,无论是新老品种,还是不同水平的氮素和密度处理下,叶片的氮素积累量始终显著高于茎秆(表3,4),且随着N水平的增加,玉米品种吐丝期叶片氮素积累和茎秆氮素积累均显著增加,同时随着密度水平的增加,玉米品种吐丝期叶片氮素积累和茎秆氮素积累也均呈增加的趋势。
2.2 新老品种花后干物质及氮素积累和转运差异
2年的试验结果显示,新品种产量显著高于老品种(表1,2),其中,2014年XY335和ZD958较ZD2平均高2.28 t/hm2,2015年则平均高1.36 t/hm2。事实上,新品种在花后干物质积累量显著高于老品种,其中,XY335和ZD958较ZD2花后干物质积累2014年平均高0.66 t/hm2,2015年高1.61 t/hm2,而且新品种花后干物质积累量占成熟期全株干物质积累量的比例明显较高(表1,2)。2年结果显示,新品种成熟期全株干物质积累量中有51.3%来自花后的干物质积累,而老品种成熟期全株干物质积累量中有48.6% 来自花后。新品种产量和成熟期干物质积累量均高于老品种,且新品种的这种优势源于花后物质积累优势。随着氮水平和密度水平的上升,2年产量均随氮水平和密度的增加而逐步上升,但在2015年各密度处理间产量差异不显著(表1,2)。
由表3,4可知,成熟期新老品种全株氮素积累量存在显著差异,其中, XY335和ZD958较ZD2全株氮素积累量2014年平均高23.5 kg/hm2,2015年高19.15 kg/hm2。与此同时,新品种成熟期籽粒氮积累量也明显高于老品种,新品种2014,2015年平均较老品种分别高17.45,18.35 kg/hm2。
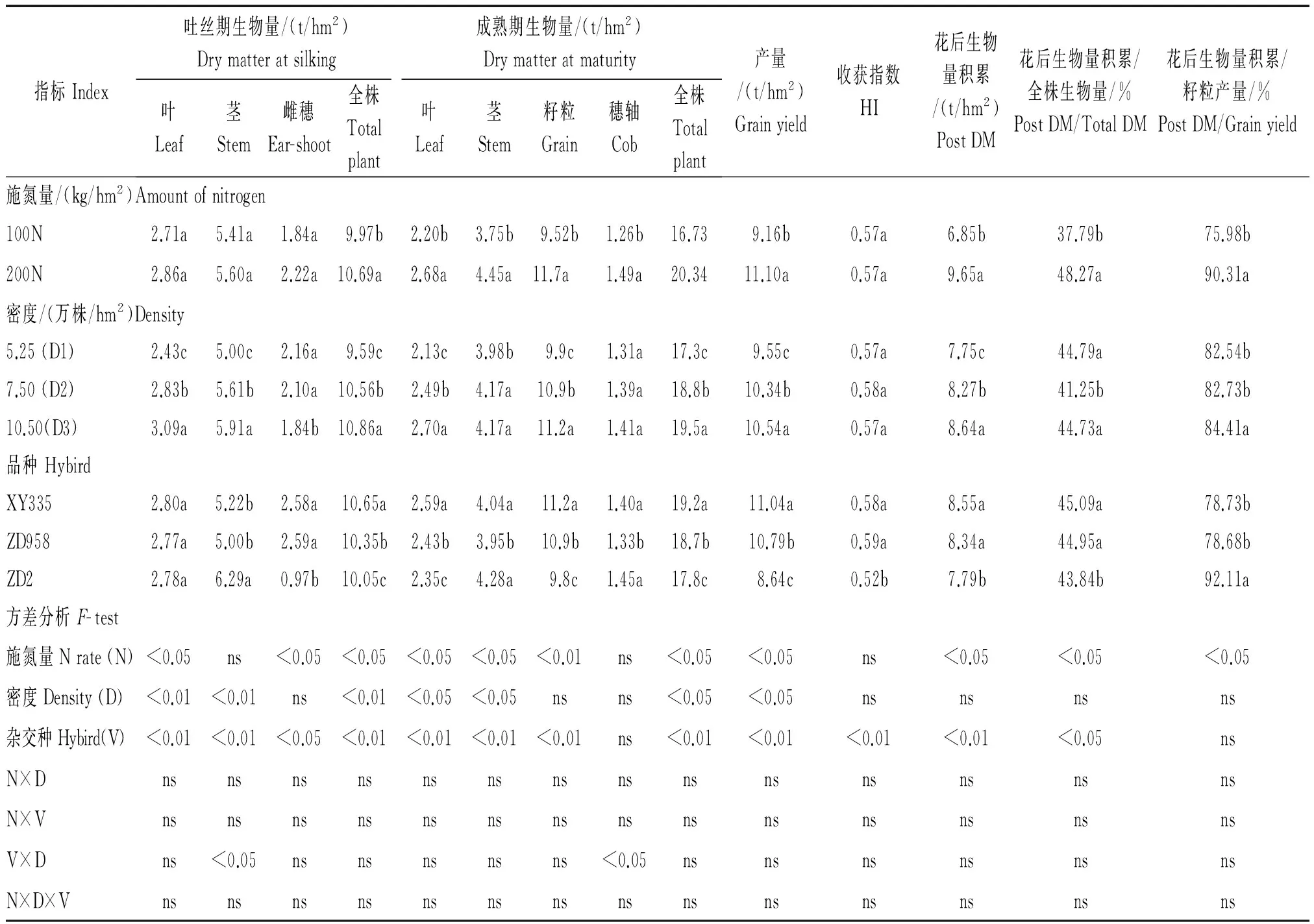
表1 2014年各处理和各时期玉米生物量及产量相关数据和方差分析Tab.1 Variance analysis of nitrogen use efficiency and its related parameters and biomass indexes and yield formation indexes of hybrids(F value)
注:DM.干物质积累量;Post N.花后氮素积累量;Rem N.花后氮素转运量;Post DM.花后干物质积累;Total DM.全株干物质积累;Total Nm.成熟期全株氮积累;Grain Nm.成熟期籽粒氮积累。不同字母表示在P=0.05水平上差异显著,ns为差异不显著。表2-6同。
Note:DM.Dry matter;Post N.Post nitrogen accumulation ;Rem N.Removable nitrogen accumulation;Post DM.Post dry matter;Total DM.Total plant dry matter;Total Nm.Total nitrogen accumulation at maturity;Grain Nm.Grain nitrogen accumulation at maturity.Values within the same year and the same row followed by different letters are significantly different according to ANOVA-protected LSD 0.05 test, ns meas no significantly different. The same as Tab.2-6 .

表2 2015年各处理和各时期玉米生物量及产量相关数据和方差分析Tab.2 Variance analysis of nitrogen use efficiency and its related parameters and biomass indexes and yield formation indexes of hybrids(F value)

表3 2014年各处理和各时期玉米氮素积累量及其相关数据和方差分析Tab.3 Variance analysis of nitrogen use efficiency and its related parameters and biomass indexes and yield formation indexes of hybrids(F value)

表4 2015年各处理和各时期玉米氮素积累量及其相关数据和方差分析Tab.4 Variance analysis of nitrogen use efficiency and its related parameters and biomass indexes and yield formation indexes of hybrids(F value)
对于成熟期籽粒氮素积累,计算方法有2种,其一,籽粒氮浓度与籽粒干物质的乘积。从这个角度看,2年籽粒氮浓度均表现为老品种高于新品种(表3,4),平均高0.73 mg/g,而新品种2年平均的籽粒干物质积累较老品种高(表1,2),新品种籽粒干物质积累较老品种在2014年高1.25 t/hm2,2015年高1.40 t/hm2。籽粒氮浓度的第2种计算方法为,成熟期籽粒氮素积累是花后转运氮素和花后吸收氮素之和。从2014年的结果看(表5),新品种花后转运氮素和花后积累氮素均明显高于老品种,平均分别高6.00,10.48 kg/hm2,2015年新品种花后转运氮素和花后积累氮素同样明显高于老品种,平均分别高4.70,14.59 kg/hm2。2年中新老品种花后转运氮素差异不显著,但是新品种花后氮素积累均显著高于老品种(表5)。由此可知,新品种成熟期籽粒氮积累量积累高于老品种可以解释为:综合所有氮水平和密度条件下,2年中新品种较老品种具有显著花后氮素吸收优势,且在较低籽粒氮浓度下具有更高的籽粒生产效率。相比老品种,新品种在气候适宜环境下(2015年)存在更高的籽粒氮积累需求,在其拉动下花后氮积累量明显升高,达到了64.58~68.55 kg/hm2;而在2014年,花前较少的降雨(图1),可能同步制约了新老品种的籽粒氮积累能力,使得花后氮积累量较2015年降低6.47~16.22 kg/hm2,但新品种仍高于老品种(表5)。2015年新老品种花后氮积累量占籽粒氮积累平均比例为43.86%,明显高于2014年的35.87%。综合2年数据,新品种成熟期产量、成熟期干物质积累量、成熟期全株氮积累量和籽粒氮积累量较老品种分别平均高1.69,1.35 t/hm2和21.3,17.9 kg/hm2,新品种较老品种优势明显。
由表3-5可知,2年试验中随着氮水平的增加,新老品种成熟期籽粒氮积累量、全株氮素积累和花后氮素积累量均呈上升趋势,高氮水平下玉米品种花后氮素积累量2年平均升高了30.1 kg/hm2,同时成熟期籽粒氮积累量、全株氮素积累分别平均上升了35.1,34.7 kg/hm2;综合品种和氮肥处理影响,随着密度的增加,2014年玉米花后氮素积累在密度之间差异不显著(表5),同时,花后氮积累占籽粒氮积累平均比例在密度之间的差异也不显著;而2015年则表现出在低密度(D1)下玉米花后氮素积累量显著高于其余2个密度(表5),同样地花后氮积累量占籽粒氮积累平均比例也表现出低密度(D1)条件下显著高于其余2个密度,即高密度压力可能抑制了花后的氮素吸收。
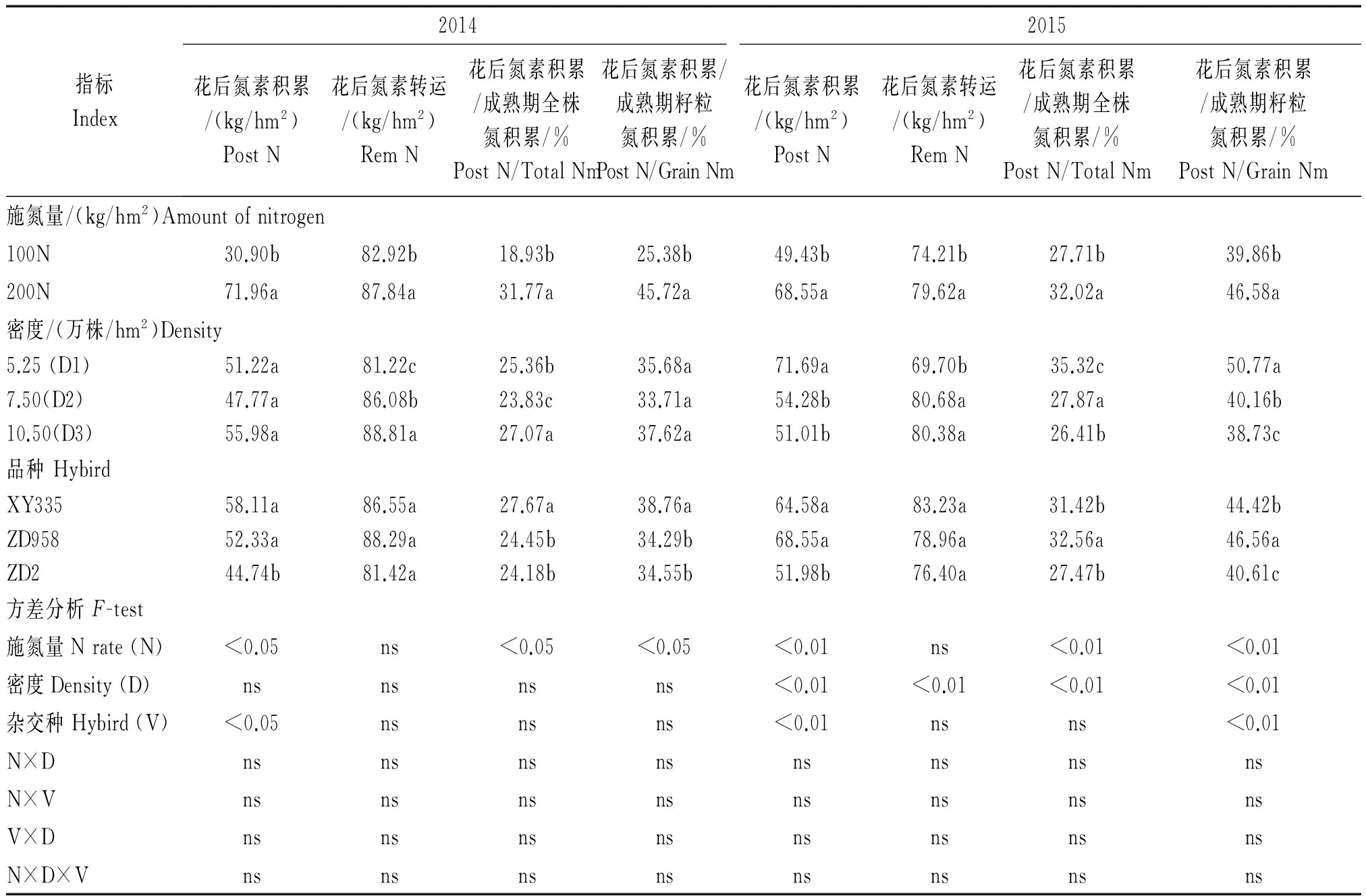
表5 2014-2015年各处理花后氮素积累转运及其相关数据和方差分析Tab.5 Variance analysis of nitrogen use efficiency and its related parameters and biomass indexes and yield formation indexes of hybrids(F value)

不同字母表示在P=0.05水平上差异显著,ns为差异不显著。Values within the same year and the same row followed by different letters are significantly different according to ANOVA-protected LSD 0.05 test, ns meas no significantly different.
由表6可知,玉米叶片的氮素转运量总体表现为新品种高于老品种,其中,XY335较ZD2显著高5.15 kg/hm2,ZD958较ZD2高2.28 kg/hm2,但并不显著。同时,与老品种相比,新品种叶片转运氮素占成熟期籽粒氮素积累的比例明显较高,新品种较老品种平均高出4.3%(图2),但是茎秆转运氮素占成熟期籽粒氮素的比例,新老品种之间并没有明显的差异(图2)。
由表6可知,新品种吐丝期茎秆生物量与叶片生物量的比值平均为2.53比老品种(2.82)显著小11.7%,也说明新品种花前干物质积累优势源于叶片的干物质积累优势。同时新品种吐丝期茎秆氮积累量与叶片氮及积累量的比值为0.76,也显著小于老品种(0.88)15.8%,也说明新品种花前干物质和氮素积累策略是更加侧重叶片干物质和氮素积累,老品种则是侧重茎秆花前干物质和氮素积累。随着氮肥水平的增加,玉米品种茎秆氮素转运显著增加,叶片氮素转运却显著下降;而随着密度的增加,玉米品种茎秆的氮素转运未出现明显的变化,但叶片的氮素转运却明显表现出中高密度(D2、D3)条件下大于低密度(D1),在中高密度(D2、D3)条件下,玉米叶片氮素转运较低密度下增大10.2 kg/hm2。综合考虑新老品种、氮肥、密度以及年份等各因素的影响,玉米叶片转运氮素占吐丝期叶片氮积累量的比例为60.26%~66.31%,均值保持在63%左右,而茎秆转运氮素占吐丝期茎秆氮积累量的比例为57.13%~62.64%,均值维持在60%左右(表6)。

表6 2014-2015年各处理玉米花后叶片和茎秆氮素积累转运积累及其相关数据和方差分析Tab.6 Variance analysis of nitrogen use efficiency and its related parameters and biomass indexes and yield formation indexes of hybrids(F value)
值得注意的是,新品种吐丝期茎秆生物量与叶片生物量的比值显著小于老品种(表6),XY335和ZD958 2年平均较ZD2分别小12.4%和11.0%,平均为11.7%。说明老品种花前更多的将干物质分配流向茎秆,新品种则更多的流向叶片。新品种吐丝期茎秆氮积累量与叶片氮及积累量的比值也显著小于老品种(表6),新品种平均为0.76,而XY335和ZD958的2年平均较ZD2分别小17.3%和14.3%,平均为15.8%,说明新品种花前的氮素积累侧重于叶片,而老品种侧重于茎秆。而且新老品种花前叶片和茎秆干物质积累量,在整个花后籽粒灌浆期,也会出现不同程度的外运。2年间,新品种平均有6.21%的花前叶片干物质在整个花后籽粒灌浆期向籽粒中转运,有24.7%的花前茎秆干物质向籽粒中转运;而老品种平均12.7%和29.1%的花前叶片及茎秆干物质在花后籽粒灌浆期向籽粒中转运。
3 讨论
Chen等[4]研究认为,玉米花粒期转运氮素和花后氮吸收存在相互制衡作用,且转运氮素和花后吸收氮素会受到氮肥和密度的明显影响。Ciampitti等[13]和Ning等[26]在2013年研究认为,美国20世纪60年代的老品种更倾向于在花前积累更多的氮素并在灌浆期将大部分氮素转运到籽粒中,而现代品种则更专注于花后的氮素吸收,转运氮素只能满足较高的籽粒氮需求的一部分,而Abe等[27-30]则研究认为,在胁迫条件下,玉米转运氮素仍然是籽粒氮需求的稳定供应来源。上述结果仍然未说明新老品种茎、叶干物质与氮素积累转运策略的差异。本研究结果表明,玉米新品种花前干物质较老品种高0.43 t/hm2,且新品种吐丝期茎秆生物量与叶片生物量的比值为2.53,比老品种显著小11.7%,说明老品种花前更多地将干物质分配流向茎秆,而新品种则更多地流向叶片。新品种吐丝期茎秆氮积累量与叶片氮积累量的比值为0.76,也显著小于老品种15.8%,说明新品种花前干物质和氮素积累策略是更加侧重叶片干物质和氮素积累,老品种则是侧重茎秆花前干物质和氮素积累。综合2年数据,新品种成熟期产量、成熟期干物质积累量、成熟期全株氮积累量和籽粒氮积累量较老品种分别高1.69,1.35 t/hm2和21.3,17.9 kg/hm2,且成熟期全株干物质积累量中有51.3%来自花后,高于老品种(48.6%)。同时新品种在花后氮素积累和花后氮素转运两方面较老品种均存在优势,其中新品种的花后氮素转运优势应源于叶片较高的氮素转运,新品种叶片氮素运移对籽粒氮积累的贡献较老品种平均高出4.3%。说明老品种较低的花后氮素吸收、花后氮素转运和干物质转运,是导致其成熟期籽粒氮浓度和产量较新品种低的主要原因。而具体到茎、叶角度看,2年间新品种平均6.21%的花前叶片干物质在整个花后籽粒灌浆期阶段外运,有24.7%的花前茎秆干物质外运;而老品种平均12.7%和29.1%的花前叶片及茎秆干物质在整个花后籽粒灌浆期外运。说明在不同的氮素和密度条件下,新品种花前干物质和氮素积累策略是更加侧重叶片干物质和氮素积累,老品种则是侧重茎秆,且在籽粒灌浆期新品种将茎秆作为玉米干物质及氮素的“库”,籽粒的需求优先拉动茎秆的养分运移,为叶片养分的过早运移提供缓冲,从而维持了灌浆期叶片较好的氮素营养和较高的光合能力,宏观上新品种表现出更高的花后氮素吸收、花后氮素转运和干物质转运,导致其更高的成熟期籽粒氮浓度和产量。
Tsai等[17]、Abe等[27-28]、张佳等[29]、申丽霞等[30]研究认为,玉米转运氮素和吸收氮素会受到土壤氮素和种植密度等管理模式的显著影响,且在低氮条件下转运氮素是籽粒氮的主要来源。Ciampitti等[10]研究认为,转运氮素对密度压力更加敏感,高密度条件下转运氮素较吸收氮素对吐丝期全株氮素积累和籽粒灌浆作用更加显著。本研究认为,氮素和密度可以同步地增加新老品种吐丝期茎叶的氮素积累量。高密度压力会抑制花后的氮素吸收,但是会提高叶片的氮素转运,以抵消玉米密度压力下而减弱的花后氮吸收。而且综合考虑新老品种、氮肥、密度以及年份等各因素影响,玉米叶片转运氮素占吐丝期叶片氮积累量的比例始终保持在62%左右,而茎秆始终维持在60%左右。在多年来品种更替过程中,新老品种叶片和茎秆的氮素转运极限是相近的,差别在于新品种具有更高效氮素积累和转运策略,以在不同栽培条件下获得更高的产量。
[1] 陈延玲.协调玉米高产与氮高效转运的机制[D].北京:中国农业大学, 2015.
[2] 吕 鹏,张吉旺.施氮量对超高产夏玉米产量及氮素吸收利用的影响[J].植物营养与肥料学报, 2011, 17(4): 852-860.
[3] 王永军.超高产夏玉米群体质量与个体生理功能研究[D].泰安:山东农业大学, 2008.
[4] Chen K, Kumudini S V, Tollenaar M, et al. Plant biomass and nitrogen partitioning changes between silking and maturity in newer versus older maize hybrids[J]. Field Crops Research, 2015, 183: 315-328.
[5] 钱春荣,于 洋,宫秀杰,等.黑龙江省不同年代玉米杂交种氮肥利用效率对种植密度和施氮水平的响应[J].作物学报, 2012, 38(11): 2069-2077.
[6] 张福锁,王激清,张卫峰,等.中国主要粮食作物肥料利用率现状与提高途径[J].土壤学报, 2008, 45(5): 915-924.
[7] 吕丽华,王 璞,鲁来清,等.不同冠层结构下夏玉米产量形成的源库关系[J].玉米科学, 2008, 16(4): 66-71.
[8] Ciampitti I A, Vyn T J. A comprehensive study of plant density consequences on nitrogen uptake dynamics of maize plants from vegetative to reproductive stages[J].Field Crops Res, 2011, 121(1): 2-18.
[9] Ciampitti I A, Vyn T J. Physiological perspectives of changes over time in maize yield dependency on nitrogen uptake and associated nitrogen efficiencies: a review[J]. Field Crops Res, 2012, 133: 48-67.
[10] Ciampitti I A, Murrell S, Camberato J, et al. Physiological dynamics of maize nitrogen uptake and partitioning in response to plant density and N stress factors: I. Vegetative phase[J]. Crop Sci, 2013, 53(5):2105-2119.
[11] Mi G H, Liu J A, Chen F J. Netrogen uptake and remobilization in maize hybirds differing in leaf senescence[J]. Journal of Plant Nutrition, 2003, 26(1):237-347.
[12] Ciampitti I A, Murrell S, Camberato J, et al. Physiological dynamics of maize nitrogen uptake and partitioningin response to plant density and nitrogen stress factors: II. Reproductive phase[J]. Crop Sci, 2013, 53(6):2588-2602.
[13] Ciampitti I A, Vyn T J. Grain nitrogen source changes over time in maize: a review[J]. Crop Science, 2013, 53(2): 366-377.
[14] Gallais A, Coque M, Gouis J, et al. Estimatingthe proportion of nitrogen remobilization and of post-silking nitrogen uptake allocated to maize kernels by nitrogen-15 labeling[J]. Crop Sci, 2007,47(2):685-693.
[15] Below F E, Christensen L E, Reed A J, et al. Availability of reduced N and carbohydrates for ear development of maize[J]. Plant Physiol, 1981, 68(5):1186-1190.
[16] 申丽霞,王 璞.不同基因型玉米氮素吸收利用效率研究进展[J].玉米科学, 2016, 24(1): 50-55.
[17] Tsai C, Huber D, Warren H, et al. Effects of cross-pollination on dry matter accumulation, nutrient partitioning and grain yield of maize hybrids grown under different levels of N fertility[J]. J Sci Food Agron, 1991, 57(2):163-174.
[18] Fonzo N, Motto M, Maggiore T, et al. N-uptake, translocation and relationships among N-related traits in maize as affected by genotype[J]. Agronomie,1982,2:789-796.
[19] 马 庆,李 猛.我国玉米育种与生产中189份自交系氮利用效率评价[J].玉米科学, 2015, 23(5): 6-11.
[20] Triboi E, Triboi-Blondel A. Productivity and grain or seed composition: anew approach to an old problem-invited paper[J]. Eur J Agron, 2002, 16(3):163-186.
[21] Pan W L, Camberato J J, Moll R H, et al.Altering source-sink relationships in prolific maize hybrids: consequences for nitrogen uptake and remobilization[J]. Crop Sci,1995,35:836-845.
[22] Rajcan I, Tollenaar. MSource: sink ratio and leaf senescence in maize: Ⅰ. Dry matter accumulation and partitioning during grain filling[J]. Field Crops Res, 1999, 60(3):245-253.
[23] Rajcan I, Tollenaar M. Source: sink ratio and leaf senescence in maize:Ⅱ. Nitrogen metabolism during grain filling[J]. Field Crops Research, 1999, 60(3): 255-265.
[24] 崔晓朋,郭家选,刘秀位,等.不同种植模式对夏玉米光能利用率和产量的影响[J].华北农学报, 2013, 28(5): 231-238.
[25] 齐 华,梁 熠,赵 明,等.栽培方式对玉米群体结构的调控效应[J].华北农学报, 2010, 25(3): 134-139.
[26] Ning P, Li S, Yu P, et al. Post-silking accumulation and partitioning of dry matter, nitrogen, phosphorus and potassium in maize varieties differing in leaf longevity[J]. Field Crops Res,2013, 144:19-27.
[27] Abe A, Adetimirin V, Menkir A, et al. Performance of tropical maize hybrids under conditions of low and optimum levels of nitrogen fertilizer application grain yield, biomass production and nitrogen accumulation[J]. Maydica,2013,58(2):141-150.
[28] Gallais A, Coque M. Genetic variation and selection for nitrogen use efficiency in maize:A synthesis[J]. Maydica, 2005, 50(3/4): 531-547.
[29] 张 佳,王永亮,郭彩霞,等.不同土壤肥力条件下高产玉米氮肥最佳用量研究[J].山西农业科学, 2013, 41(2): 155-159.
[30] 申丽霞,王 璞,孙西欢.不同种植密度下施氮对夏玉米物质生产及穗粒形成的影响[J].山西农业科学, 2008, 36(1): 41-44.
