软体动物腹足纲分类学研究进展——从近海到深海*
2018-01-10张素萍张树乾
张素萍 张树乾
软体动物腹足纲分类学研究进展——从近海到深海*
张素萍①张树乾
(中国科学院海洋研究所 海洋生物分类与系统演化实验室, 青岛 266071)
随着科学技术的不断进步和研究的不断深入, 我国关于海洋的研究已由浅海逐步向深海发展, 不断取得新的发现和认知。自2014年4月起, 中国科学院海洋研究所依托先进的“科学”号海洋科学综合考察船, 搭载“发现”号水下机器人(remotely operated vehicle, ROV), 先后对冲绳海槽热液区、南海冷泉区、雅浦海山区以及马努斯热液区等深海区域进行了综合性科学考查, 采集到了大量深海腹足纲(Gastropoda)标本, 其中有一些为未经发现和描述的新种, 极大地推动了我国关于深海软体动物物种多样性研究, 实现了深海环境和软体动物资源的新认知。本文对近年来从深海采集到的腹足类标本进行了整理分类, 对其中的11属13种进行描述, 并对深海特殊环境中腹足纲物种多样性进行了总结, 对中国软体动物腹足纲从近海至深海的分类现状进行研究论述。
腹足类; 热液区; 冷泉区; 海山区; 展望; 从近海到深海
新中国成立以来, 我国对海洋腹足纲(Gastropoda)的研究主要集中在近海浅水区域。自20世纪50年代初开始, 在张玺和齐钟彦教授的倡导下, 我国贝类工作者对中国海洋软体动物进行了全面、系统的区系调查和分类学研究, 开展了数次大规模的海洋科学调查, 包括全国海洋综合调查(1958~1960年)、中越北部湾联合调查(1959~1960年, 1962年)、东海大陆架调查(1975~1981年)、中国科学院海洋研究所西沙群岛生物调查(1975~1980年)、中国科学院南沙群岛生物调查(1985年, 1987~2000年)以及近年开展的黄海冷水底栖生物群落结构和多样性特点的研究(2007~ 2012年)、海南岛及西沙群岛海岸带生物资源考察(2007~2010年)等, 获取了大量的贝类标本和信息资料, 基本摸清了我国近海贝类重要经济类群的物种数量与区系特点, 陆续发现了一些新物种和中国新记录种。中国科学院海洋研究所是我国海洋贝类分类学研究的主导力量, 对海洋贝类学研究已有半个多世纪的科学积累。据统计, 迄今我国沿海已发现报道软体动物4000余种, 其中腹足类约3000种(张素萍, 2008), 双壳类约1200种(徐凤山, 2008); 先后发表和出版了数百篇在国内外有广泛影响力的贝类学研究论文以及数十部专著。这些研究成果, 为中国近海海洋生物多样性研究与资源保护和利用提供了宝贵的参考文献和基础资料。
深海作为海洋系统的重要组成部分, 不仅孕育着独特的生态系统和生命过程, 还可能对上层海洋热量耗散产生影响。因此, 深海研究在整个地球科学和全球变化研究中都处于十分重要的地位, 开展相关深海研究符合我国科学发展及国家战略的需求(李超伦和李富超, 2016)。当前, 国际上关于深海的研究主要集中在深海海山区、热液区、冷泉区等特殊环境区域。这些独特的生态系统对于研究生命起源与演化、深海生物与极端环境相互作用、深海相关资源的开发利用等方面都具有十分重要的科学意义。过去几十年, 受限于深海探测装备的落后, 我国对于深海软体动物多样性研究几乎处于空白阶段, 仅在南海和东海的冲绳海槽附近进行过几次深海拖网作业, 但收获并不大。进入21世纪以来, 随着我国科学技术的不断发展, 一些深海探测器如“蛟龙号”载人深潜器的研制成功, 标志着我国在深海探测技术方面的突破。特别是近年来, 在中国科学院A类战略性先导科技专项“热带西太平洋海洋系统物质能量交换及其影响”的资助下, 中国科学院海洋研究所依靠先进的“科学”号海洋综合考察船以及搭载的“发现”号水下机器人(remotely operated vehicle, ROV)、电视抓斗等设备分别对冲绳海槽热液区和南海冷泉区(2014年4月和2016年8月)、雅浦海山区(2014年12月)、马努斯热液区(2015年6月)以及雅浦-马里亚纳海山区(2016年3月)进行了数次综合调查, 采集到了大量深海生物标本, 其中包括许多腹足类标本, 为我国进行深海腹足纲物种多样性研究提供了重要材料。通过深入细致的整理和分类, 本文在我国首次报道了针对这些深海贝类标本的研究结果, 并围绕发现的种类对当前世界范围内关于深海腹足纲的分类现状进行了研究论述。
1 热液区和冷泉区腹足纲生物概述
海底热液(hydrothermal vent)俗称“黑烟囱”, 是指在大陆板块与海洋板块之间的火山口, 在这里海水沿着地壳缝隙深入地壳内部(深度可达2~3km), 并经岩浆加热喷涌而出。被加热后的海水富含矿物质, 温度可高达600°C, 喷口处的热液温度亦可达到400°C左右(冯军等, 2005)。而冷泉(cold seep)是指在大陆边缘海底, 富含天然气水合物(甲烷和石油)的流体组分在海底表面的溢出, 其温度与周围海水温度相差不大。
自1977年发现第一个热液口之后, 1984年第一个冷泉口也在墨西哥湾被发现。迄今, 在世界范围内共发现500余个热液口和冷泉口。其中, 关于腹足类研究的热点区域主要包括大西洋中嵴(Mid-Atlantic)、印度洋中嵴(Central Indian Ridge)、胡安·德富卡海岭(Juan de Fuca ridge)、东太平洋隆起(East Pacific Rise)、加拉帕戈斯群岛(Galapagos Rift)、马里亚纳盆地(Mariana Basin)、马努斯盆地(Manus Basin)、劳盆地(Lau Basin)、冲绳海槽(Okinawa Though)等区域。被发现和报道的热液区和冷泉区腹足纲种类约为231种(Sasaki et al., 2010; Johnson et al., 2014; Chen et al., 2015, 2016; Zhang et al., 2016b; Zhang and Zhang, 2017), 是这类区域中仅次于甲壳动物的第二大门类。其中, 在热液区发现的腹足纲种类为153种, 超过95%物种为热液区特有种; 冷泉区特有种比例稍低, 在发现的83种中有55种为冷泉区特有种。热液区和冷泉区共有种为5种, 占总数量的2%。在报道的104个属中, 热液区和冷泉区的共有属为17个, 约占总属数的16%。热液区和冷泉区有19个共有科, 约占总科数的46%。
腹足纲动物除营寄生生活的种类外, 通常都具有发达而锐利的齿舌。生活在热液区和冷泉区的腹足类可利用齿舌刮取岩石表面沉积的细菌或直接捕食热液口其他生物, 也可通过鳃滤食周围水体中的悬浮颗粒, 从而获得食物。除此之外, 绝大多数种类可与化能自养菌共生。化能自养菌利用寄主获取的H2S和O2制造有机物, 部分用来滋养寄主。腹足类与化能自养菌的共生关系可分为两大类, 即外共生(ectosymbiosis)和内共生(endosymbiosis)。外共生是指自养菌共生在腹足类体表的一些特殊结构上, 内共生是指自养菌栖息在腹足类鳃上皮组织或体内。
对深海海底热液区和冷泉区生态系统的研究是进入21世纪以来的深海研究重点领域。中国科学院海洋研究所分别于2014年4月和2016年8月两次对冲绳海槽热液区和南海冷泉区进行了调查, 2015年6月对马努斯热液区进行了科学考查, 先后采集了一批腹足类标本, 通过分类整理, 共鉴定出20余种。本文对其中已确立分类地位的9属11种的形态特征和生态习性等进行了描述。
1.1 热液区和冷泉区物种描述
(1) 探花深海笠贝Sasaki, Okutani & Fujikura, 2003 (图1a, 2e)
Sasaki, Okutani & Fujikura, 2003: 190, figs. 3B, 4, 5.
标本采集地 冲绳海槽热液区。
观察标本 20余个标本。
形态特征 贝壳呈笠状, 椭圆或卵圆形, 壳长 8.7mm, 宽6.2mm, 高3.6mm, 壳顶位于中央近前方, 顶部常被腐蚀。壳面为白色, 雕刻有细密的环行螺肋和极细弱的辐射状生长纹。壳口内乳白色, 有光泽, 边缘薄。触角细长, 无眼点。头部圆, 无附属结构。齿舌齿式为0·1·0·1·0。
习性及分布 生活于996~1000m深的热液区; 目前仅发现于冲绳海槽。
(2) 乳白深海笠贝Zhang, Zhang & Zhang, 2016b (图1b, 2f)
Zhang, Zhang & Zhang, 2016b: 2, figs. 1~9.
标本采集地 南海冷泉区。
观察标本 5个标本。
形态特征 贝壳呈卵圆形, 壳长19.6mm, 宽17.0mm, 高8.5mm, 壳质厚。壳面乳白色, 较光滑, 上具排列细密的环行螺纹和细弱的辐射状生长纹。壳口边缘增厚, 具齿状突起。触角粗短, 无眼点。头部圆, 无附属结构。齿舌齿式为0·1·0·1·0。
习性及分布 生活在1000m以上的深海底; 目前仅发现于南海冷泉区。
讨论 该属种类仅分布于西太平洋海域, 全部生活在热液区或冷泉区。乳白深海笠贝是我国在南海冷泉区发现的新种, 它除齿舌形态与同属其他种存在明显不同外, 其外壳也明显比其他种厚, 最厚处可达1.5 mm。
(3) 琉球马蹄螺Okutani, Sasaki & Tsuchida, 2000 (图1c, 2g)
Okutani, Sasaki & Tsuchida, 2000: 268, figs. 2~5.
标本采集地 冲绳海槽热液区。
观察标本 1000余个标本。
形态特征 贝壳小, 壳长5.5mm, 宽4.8mm, 近圆形; 缝合线深, 螺层膨圆。贝壳的宽度大于高度, 螺旋部极低小, 体螺层宽大。壳面光滑, 呈白色, 被有浅绿色的薄壳皮, 生长纹极细密。壳口大, 圆形。厣角质, 淡黄色, 半透明, 多旋, 核位于中央。脐孔狭小。齿舌为扇状型(rhipidoglossate), 齿式为n·11·1·11·n。
习性及分布 研究用的标本采自于996~ 1242m深的海底; 目前仅发现于冲绳海槽。
讨论 本研究所观察标本与模式标本形态特征一致, 但不同之处在于模式标本的齿舌具12枚侧齿, 而本研究观察标本具11枚侧齿。作者认为该差异应属于种内变异。
(4) 坚果鳞笠贝(Okutani, Fujikura & Sasaki, 1993) (图1d)
Okutani, Fujikura & Sasaki, 1993: 132, figs. 22~27.
标本采集地 冲绳海槽热液区。
观察标本 2000余个标本。
形态特征 贝壳呈帽贝状, 壳形椭圆, 壳长11.0mm, 宽8.0mm, 高3.5mm, 前端稍窄, 壳顶位于后端, 弯曲呈鸟喙状。自顶部向周缘环生有粗糙的生长纹, 壳表被有一层厚的绿色壳皮, 延伸于整个壳口边缘。壳口特大, 卵圆形, 边缘厚, 内灰白色。触角粗短, 眼点缺。足呈椭圆状, 小。外套膜边缘具细小的乳突。颈部左侧具一乳突, 无雄性交接器。
习性及分布 标本采自于996~1242m深的热液区; 目前仅发现于冲绳海槽。
讨论 本种壳皮颜色有变化, 模式标本的壳皮为深棕色(Okutani et al., 1993), 而作者所观察标本的壳皮均为绿色。壳皮颜色的变化可能是由于不同个体处于不同生态环境所导致, 具深棕色壳皮的个体可能离热液口更近一些。
(5) 斯氏鳞笠贝Beck, 1993 (图1e, 1f, 2h)
Beck, 1993: 171, pl. 1~4, figs. 2~3.
标本采集地 马努斯弧后盆地。
观察标本 650个标本。
形态特征 贝壳小, 壳形椭圆, 两端窄, 壳长5.5mm, 宽3.8mm, 高2.3mm, 壳顶位于后端。贝壳表面被有一层橄榄绿色或深棕色的壳皮。壳表光滑, 除细密的生长纹外, 无其他雕刻。壳口边缘平直或不在同一平面上。贝壳内缘呈白色。触角粗短, 眼点缺。足呈椭圆状, 小。外套膜边缘具细小的乳突。颈部左侧具一乳突, 无雄性交接器。齿舌齿式为n·5·1·5·n。
习性及分布 生活于1450~2050m深的热液区; 目前仅发现于马努斯弧后盆地。
讨论 本种除壳皮颜色有变化外(橄榄绿或深棕色), 其贝壳高度及壳口形状也常有变化, 这可能是不同的个体为了适应不同形态的底质所致。
(6) 光滑普氏螺Okutani, Tsuchida & Fujikura, 1992 (图1g)
Okutani, Tsuchida & Fujikura, 1992: 143, figs. 17~21.
标本采集地 冲绳海槽热液区。
观察标本 2000个标本。
形态特征 贝壳小型, 壳长10.0mm, 宽6.5mm, 呈长卵圆形; 螺旋部圆锥形, 体螺层高大。壳顶无(由于栖息于深海与水压的关系, 在拖出水面时, 所有标本的壳顶全部破损)。外被有一层橄榄绿或黄褐色的壳皮。壳口大, 卵圆形, 外唇外边缘薄; 内唇弧形, 有一个极弱的前水管沟。
习性及分布 栖息于996~1242m深的热液区和冷泉区; 发现于冲绳海槽和日本的相模湾。
讨论 本种与Sasaki, Ogura, Watanabe & Fujikura, 2016最为相似, 但本种的壳表更为光滑、无雕刻, 且壳形较瘦。
(7) 深海灰蝾螺(Okutani & Fujikura, 1990)(图1h)
Okutani & Fujikura, 1990: 85, figs. 2~6.
标本采集地 冲绳海槽热液区。
观察标本 3个标本。
形态特征 贝壳呈低圆锥形, 壳长15.2mm, 宽16.0mm。螺旋部低, 壳顶常破损, 体螺层宽大。壳面雕刻有稀疏而发达的螺旋肋, 两肋间有明显斜行生长纹, 贝壳底部形成一个平面, 其上具有2~3条明显的细螺肋。壳面呈灰白色。壳口大, 近圆形, 内唇滑层稍厚, 向上翻卷, 遮盖脐部, 仅留一小的空隙; 外唇薄, 弧形。
习性及分布 生活水深为700~720m; 目前仅发现于冲绳海槽。
讨论 本属种类在普通深海海底、热液区和冷泉区均有分布。迄今, 在热液区和冷泉区发现的种类共4种, 冲绳海槽(Okutani and Fujikura, 1990; Okutani, 2001)、路易斯安那(Warén et al., 1993)以及智利(Sasaki et al., 2010)均有分布。
(8) 斯坦拟蛾螺Okutani & Ohta 1993 (图1i)
Okutani & Ohta, 1993: 220, figs. 6~7.
标本采集地 马努斯弧后盆地热液区。
观察标本 2个标本。
形态特征 贝壳中等大小, 壳长28.0mm, 宽18.1mm, 呈纺锤形。壳面为白色。壳表雕刻具粗细不均的螺肋和细密的生长纹。缝合线明显。螺旋部小, 体螺层膨大, 约占整个壳长的3/4。壳口呈椭圆状, 外唇边缘呈锯齿状; 内唇直, 滑层弱。前水沟宽短。
习性及分布 生活于1740~2750m深的热液区; 发现于北斐济盆地和马努斯弧后盆地。
讨论 本种的外壳厚度有变化。生活在马努斯弧后盆地的个体通常比生活在北斐济盆地的个体外壳更厚。这可能是不同的个体为适应不同的深度所采取的适应策略。
(9) 纳氏热液螺Bouchet &Warén, 1991 (图1j)
Bouchet & Warén, 1991
Beck, 1991: 282.
标本采集地 马努斯弧后盆地热液区。
观察标本 20个标本。
形态特征 贝壳大, 壳长可达95.0mm。幼体壳皮呈棕色, 成体壳皮呈黑色, 贝壳表面常覆盖一层厚的棕色矿物质。胚壳常腐蚀, 螺层膨圆。缝合线深。各螺层在近缝合线处具一螺旋列, 其由月牙状的生长疤组成。幼壳表面雕刻明显, 由粗细不均的念珠状螺肋组成; 成壳表面雕刻不明显, 仅具纵向生长纹。壳口近圆形。具脐孔。厣角质, 呈椭圆形。
习性及分布 生活在1740m深左右的热液区; 目前发现于马努斯弧后盆地、北斐济盆地(North Fiji Basin)以及劳盆地(Lau Basin)。
讨论 该种为热液区优势种, 常呈大面积密集分布。加厚的壳皮、厣等特征均是对深海热液特殊环境的适应。
(10) 克氏球刺螺Johnson, Warén, Tunnicliffe, Van Dover, Wheat, Schultz & Vrijenhoek, 2014 (图2a)
Johnson, Warén, Tunnicliffe, Van Dover, Wheat, Schultz & Vrijenhoek, 2014: 288, fig. 2.2.
标本采集地 马努斯弧后盆地热液区。
观察标本 20个标本。
形态特征 贝壳大, 呈球形, 壳长可达85.0mm。壳表颜色为黄色或棕色。螺旋部小, 体螺层膨大, 占整个壳长的绝大部分。壳表被有分布规则的长鬃毛, 其顶部常弯曲。壳口呈耳状, 外唇薄; 轴唇弯曲, 具明显的滑层。前水沟宽短。厣角质, 棕色, 呈椭圆形, 核在前端。
习性及分布 栖息于1480~2700m深的热液区; 目前发现于马努斯弧后盆地、斐济盆地以及劳盆地。
讨论 该属之前被认为只有1种, 即Okutani & Ohta, 1988, 广泛分布于西太平洋以及印度洋的热液区。近来通过分子生物学研究表明, 它们属于5个不同的种(Johnson et al., 2014), 其外部形态的相似可能是趋同进化的结果。

图1 热液区及冷泉区种类(a. 探花深海笠贝, 8.7mm; b. 乳白深海笠贝, 19.6mm; c. 琉球马蹄螺, 5.5mm; d. 坚果鳞笠贝, 11.0mm; e、f. 斯氏鳞笠贝, 5.2mm; g. 光滑普氏螺, 10.0mm; h. 深海灰蝾螺, 15.2mm; i. 斯坦拟蛾螺, 28.0mm; j. 纳氏热液螺, 60.0mm)
(11) 马努斯片脐螺Zhang & Zhang, 2017 (图2b, 2i)
Zhang & Zhang, 2017: 78, figs. 1~32.
标本采集地 马努斯弧后盆地热液区。
观察标本 130个标本。
形态特征 贝壳小, 壳长8.8mm, 宽5.7mm, 高2.9mm, 呈帽贝状, 但早期螺层呈螺旋状。贝壳表面被有橄榄绿色的壳皮。螺旋部小, 位于贝壳的右后部。胚壳圆, 具网状雕刻。成壳表面具发达的螺肋和纵肋, 两者相交呈格子状。雌性左右触角大小相等; 雄性的左触角膨大, 特化为雄性生殖器, 为右触角的3~4倍。足近圆形, 后端具5~7对上足触角。鳃大, 为双栉鳃。齿舌为扇状齿舌, 齿式为n·4·1·4·n。厣非常薄, 位于足的后端。
习性及分布 生活在1740m深的热液区黑色岩石上; 目前仅发现于马努斯弧后盆地。
讨论 脐螺科(Neomphalidae)种类是深海热液区的特有类群, 主要分布在东太平洋隆起、加拉帕戈斯、劳盆地以及马里亚纳弧后盆地, 目前全世界已发现6属9种, 其贝壳呈帽贝状或螺旋状。而马努斯片脐螺的贝壳形状如帽贝, 但早期壳层仍保留着螺旋状, 后足上具退化的厣, 这些特征均表明该种类为脐螺科动物由螺旋状向帽贝状演化的中间过渡类型。
2 海山区腹足纲生物概述
海山(seamounts)一般指大洋深海中高度大于1000m、顶部不突出海平面的隆起地形, 广义上的海山也包括高度小于1000m的海丘(hills或knolls)。海山通常由海底的火山运动所形成, 高度通常在1000~4000m, 其顶部在海平面以下几百米至数千米, 在深海中呈链状分布或以细长形集群出现。目前, 世界范围内已发现的海山近25000座, 其中约60%集中于太平洋。由于其特殊的地质结构, 孕育和支持着独特的生态群落, 海山通常被认为在海洋物种扩散和进化中扮演着重要的中间角色。近几十年关于海山的生物区系研究, 衍生出了一系列假说, 如海山特有种假说、物种汇源假说、孤岛假说、绿洲假说等, 但由于调查采样的不足, 这些假说均缺乏说服力, 且存在着一定的争议(张均龙和徐奎栋, 2013)。海山上栖息着几乎所有门类的海洋动物, 由于各国对海山的定义及对海山动物区系的认知存在差异, 因此对海山区分布种类的统计存在着很大难度。迄今, 对海山腹足类的研究也仅见于零星报道, 如Beck(1996)对西太平洋Edison海山的腹足类进行了调查, 描述了4个新种; Gofas(2007)对东北大西洋的Lusitanian海山上分布的麂眼螺科(Rissoidae)种类进行了记述。据Seamounts Online统计, 目前分布于海山的软体动物共计约2500余个记录数, 其中大部分为腹足类。
中国科学院海洋研究所于2014年12月起对位于西太平洋海域的雅浦海山区进行了综合调查, 由于海山顶部常凸凹不平, 给拖网带来了很大的难度, 因此拖出水面的标本很有限, 在两次深海海山调查中采获的软体动物标本明显比热液和冷泉区少。目前, 共鉴定出腹足纲标本6种(包括未定种), 本文对其中2种的形态特征和生态习性等进行研究论述。
2.1 海山区物种描述
(1) 精致翁戎螺Zhang, Zhang & Wei, 2016a (图2c, 2j)
Zhang, Zhang & Wei, 2016a: 254, Figs. 1, 2A~C, 3, 4.
标本采集地 雅浦海山区。
观察标本 2个标本。
形态特征 贝壳中小型, 壳形低矮, 壳长47.8mm, 宽62.1mm, 壳质薄。胚壳约1层, 光滑。成壳约7层, 壳层平扁。前4层上螺肋和纵肋较为发达, 两者相交使壳表形成方形的雕刻; 后3层雕刻精细。贝壳底部稍凸, 具许多精细的生长纹和螺线。贝壳表面呈橘色, 有光泽。壳口椭圆形, 内具彩虹色, 裂缝短, 上缘长约30.0mm, 下缘长约24.0mm。脐孔闭合。厣棕色, 近圆形, 核在中央。
习性及分布 生活于距水面255~289m深的山顶处。目前仅知分布于雅浦海山区。
讨论 翁戎螺属于非常古老的类群, 起源于古生代寒武纪时期, 距今约5亿年, 被人们誉为贝类的“活化石”, 迄今已发现化石种1000余种, 但现生种仅32种。精致翁戎螺与寺町翁戎螺在壳形上较为相似, 但其贝壳较小, 壳表雕刻更为精细。
(2) 史氏东方蛾螺(Schepman, 1911) (图2d)
()Schepman, 1911: 54, pl. 19, fig. 6, pl. 23, fig. 11.
标本采集地 雅浦海山区。
观察标本 6个标本。
形态特征 贝壳大, 呈纺锤形, 壳长39.0mm, 宽20.1mm。贝壳表面为白色, 被有浅黄色的壳皮。成壳螺层稍凸, 约5层。螺旋部低, 呈锥形, 体螺层膨大, 约占整个壳长的3/4。缝合线明显。成壳螺层上具发达的低平螺肋, 螺肋间具细沟。无纵肋, 具精细的生长纹。壳口卵圆形, 外唇薄, 内缘光滑; 内唇具弱的滑层。前水管沟宽短。厣角质, 棕色。
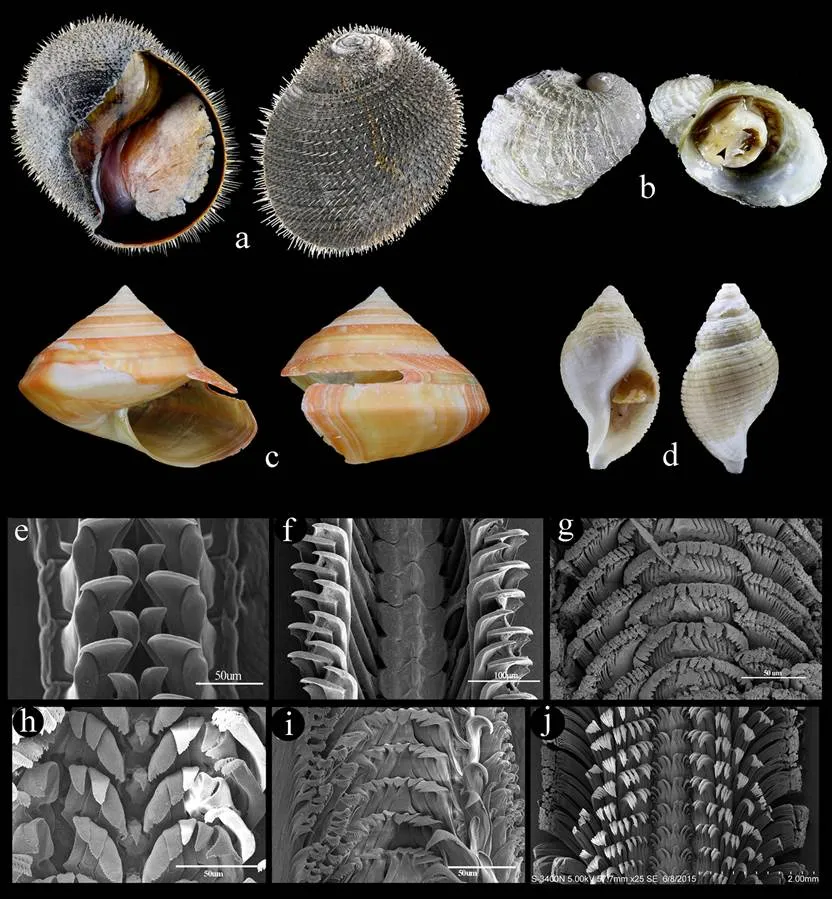
图2 海山区种类(a. 克氏球刺螺, 40mm; b. 马努斯片脐螺, 8.8mm; c. 精致翁戎螺, 47.8mm; d. 史氏东方蛾螺, 39.0mm; e. 探花深海笠贝的齿舌; f. 乳白深海笠贝的齿舌; g. 琉球马蹄螺的齿舌; h. 斯氏帽贝的齿舌; i. 马努斯片脐螺的齿舌; j. 精致翁戎螺的齿舌)
习性及分布 通常栖息于529~1600m深的海底, 本研究所用标本采集于水深1119m的海山区, 底质为有孔虫泥。广泛分布于印度-西太平洋。
讨论 本种不仅分布范围广, 而且贝壳大小变化大, 模式标本壳长为20.4mm, 但Bouchet和Warén (1985)所观察的标本长度达53.4mm。
3 展望
自2014年4月起, 我国已对深海海山、热液区、冷泉区进行了数次综合调查, 搜集了一些珍贵的深海腹足类标本和相关信息, 为开展深海贝类学研究提供了基础资料。通过分类研究发现了一些未经描述和报道的新种, 这些成果表明我国已经拥有国际一流的深海综合调查能力和研究条件。然而, 总体来讲, 深海底栖生物多样性研究在我国尚处于起步阶段, 对目前存在的一些问题仍需完善和加强。探讨深海生命的起源与进化、深海物种对极端生态环境的适应以及深海种类组成等正越来越受到人们的关注, 相信通过我们不懈努力和深入细致的观察研究, 将有更多的新种、新属、甚至新科等被发现和报道。
3.1 运用多手段对深海腹足纲种类进行分类鉴定
深海腹足类样品采集难度大, 有的物种标本数量往往较少。因此, 对其进行准确的鉴定显得尤为重要, 同时也存在一定的难度。由于深海环境复杂, 深海腹足纲种类常存在趋同或趋异进化现象, 所以简单地依靠贝壳的外部形态对深海腹足类进行鉴定容易受到主观因素的干扰和影响, 难以得到正确、可信的结果, 甚至会导致分类上的混乱, 阻碍相关研究的开展。因此, 亟需一套准确、有效的手段来解决这一问题。在对深海腹足类进行分类时, 我们要在观察外部形态的基础上, 利用光学显微镜和扫描电子显微镜对软体部分的解剖特征以及齿舌的细微结构进行观察和描述, 以期达到准确的鉴定。同时, 还要引入分子生物学研究等新的方法与技术手段对深海底栖腹足纲进行分类学和系统学研究, 确立分类地位, 探讨系统演化关系。
3.2 重视多学科交叉与融合
深海特殊的地理地貌和物理、化学环境孕育了独特的生态环境。很多深海区域, 尤其是海山区、热液区和冷泉区具有很多腹足类特有种。目前, 国际上关于深海腹足纲的研究主要集中在物种多样性上, 但这些特有种是如何形成的?在深海中又是如何进化和演变的?是如何适应深海极端环境的?直到目前仍知之甚少。回答这些问题必需从海洋系统的角度综合考虑分析, 通过多学科多领域的交叉融合, 从不同角度阐述深海腹足纲物种多样性及其生命起源过程。此外, 深海特殊功能基因研究、深海生命起源与进化研究、深海海底生物资源开发与利用等研究也是重要的研究发展方向。
致谢 感谢“科学”号全体工作人员在标本采集过程中给予的热情帮助; 感谢参加深海热液区、冷泉区以及海山区综合科学考查的科考人员, 为本研究提供了深海贝类标本和信息资料。
冯 军, 李江海, 牛向龙, 2005. 现代海底热液微生物群落及其地质意义. 地球科学进展, 20(7): 732—739
张均龙, 徐奎栋, 2013. 海山生物多样性研究进展与展望. 地球科学进展, 28(11): 1209—1216
张素萍, 2008. 软体动物——腹足纲. 见: 刘瑞玉. 中国海洋生物名录. 北京: 科学出版社, 462
李超伦, 李富超, 2016. 深海极端环境与生命过程研究现状与对策. 中国科学院院刊, 31(12): 1302—1307
徐凤山, 2008. 软体动物——双壳纲. 见: 刘瑞玉. 中国海洋生物名录. 北京: 科学出版社, 548
Beck L A, 1991.n. gen. et n. sp.-a new mesogastropod (Gastropoda: Prosobranchia) from hydrothermal vents in the Manus Back-Arc Basin (Bismarck Sea, Papua New Guinea). Annalen des Naturhistorischen Museums in Wien, 92: 277—286
Beck L A, 1993 Morphological and anatomical studies on a new lepetodrilacean limpet (Gastropoda, Prosobranchia) from hydrothermal vents at the Manus Back-Arc Basin (Bismarck Sea, Papua New Guinea). Annalen des Naturhistorischen Museums in Wien, 94—95: 167—179
Beck L A, 1996. Morphology and anatomy of new species of neolepetopsid, acmaeid, fissurellid and pyropeltid limpets from Edison Seamount off Lihir Islands (West Pacific). Arch Molluskenkd, 125(1—2): 87—103
Bouchet P, Warén A, 1985. Mollusca Gastropoda: taxonomical notes on tropical deep water Buccinidae with descriptions of new taxa. Memoires du Museum National d'Histoire Naturelle Serie A Zoologie 133: 457—499
Bouchet P, Warén A, 1991., a new gastropod from hydrothermal vents, probably associated with symbiotic bacteria. Comptes Rendus de l’Académié des Sciences Série III, 312(10): 495—501
Chen C, Linse K, Copley J T et al., 2015. The ‘scaly-foot gastropod’: a new genus and species of hydrothermal vent-endemic gastropod (Neomphalina: Peltospiridae) from the Indian Ocean. Journal of Molluscan Studies, 83(3): 322—334
Chen C, Ogura T, Hirayama H et al., 2016. First seep- dwelling(Gastropoda: Abyssochrysoidea) species discovered from a serpentinite-hosted seep in the Southeastern Mariana Forearc. Molluscan Research, 36(4): 277—284
Gofas S, 2007. Rissoidae (Mollusca: Gastropoda) from northeast Atlantic seamounts. Journal of Natural History, 41(13—16): 779—885
Johnson S B, Warén A, Tunnicliffe V et al., 2014. Molecular taxonomy and naming of five cryptic species ofsnails (Gastropoda: Abyssochrysoidea) from hydrothermal vents. Systematics and Biodiversity, 13(3): 278—295
Okutani T, Fujikura K, 1990. A new turbinid gastropod collected from the warm seep site in the Minami- Ensei Knoll, west of the Amami-Oshima Island, Japan. Venus, 49(2): 83—91
Okutani T, Tsuchida E, Fujiakura K, 1992. Five bathyal gastropods living within or near the- community of the Hatsushima Islet, Sagami Bay. Venus, 51(3): 137—148
Okutani T, Ohta S, 1993. New buccinid and turrid gastropods from North Fiji and Lau Basins. Venus, 52(3): 217—221
Okutani T, Fujikura K, Sasaki T, 1993. New taxa and new distribution records of deepsea gastropods collected from or near the chemosynthetic communities in the Japanese waters. Bulletin of the National Science Museum. Series A, Zoology, 19(4): 123—143
Okutani T, Sasaki T, Tsuchida S, 2000. Two additional new species to gastropod fauna of chemosynthetic site on north Knoll of Iheya Ridge, Okinawa Trough. Venus, 59(4): 267—275
Okutani T, 2001. Six new bathyal and shelf Trochoidean species in Japan. Venus, 60(3): 121—127
Sasaki T, Okutani T, Fujikura K, 2003. New taxa and new records of patelliform gastropods associated with chemoautosynthesis-based communities in Japanese waters. The Veliger, 46: 189—210
Sasaki T, Warén A, Kano Y et al., 2010. Gastropods from recent hot vents and cold seeps: systematics, diversity and life strategies the vent and seep biota. The Vent and Seep Biota, 33: 169—254
Sasaki T, Ogura T, Watanabe H K et al., 2016. Four New Species of(Gastropoda: Provannidae) from vents and a seep off Nansei-shoto area, southwestern Japan. Venus, 74(1—2): 1—17
Schepman M M, 1911. Prosobranchia of the Siboga Expedition. Part 4: Rachiglossa. Siboga Expeditie, 49: 247—363
Warèn A, Bouchet P, 1993. New records, species, genera, and a new family of gastropods from hydrothermal vents and hydrocarbon seeps. Zoologica Scripta, 22(1): 1—90
Zhang S P, Zhang S Q, Wei P, 2016a., a new species of pleurotomariid from Yap Seamount, Western Pacific, near Palau (Gastropoda: Pleurotomariidae). Zootaxa, 4161(2): 252—260
Zhang S Q, Zhang J L, Zhang S P, 2016b. A new species of(Gastropoda: Pectinodontidae) from a methane seep area in the South China Sea. The Nautilus, 130(1): 1—4
Zhang S Q, Zhang S P, 2017. A new genus and species of Neomphalidae from a hydrothermal vent of the Manus Back-Arc Basin, western Pacific (Gastropoda: Neomphalina). The Nautilus, 131: 76—86
Status of Gastropod Taxonomy Study in China —from Shallow Water to Deep Sea
ZHANG Su-Ping*, ZHANG Shu-Qian
(Department of Marine Organism Taxonomy and Phylogeny, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China)
With the development of science and technology, marine sciences research in China has been progressed from the exploration of shallow waters to the deep sea. This has led to new discoveries and in turn, new perceptions. Since April 2014, several deep-sea areas were investigated by Institute of Oceanology, Chinese Academy of Sciences (IOCAS). These areas include the hydrothermal vents in Okinawa Trough and Manus Back-Arc Basin, the cold seep in South China Sea, and Yap Seamount in the western Pacific. During these surveys, numerous gastropod specimens were sampled by the mechanical arm of ROV FAXIAN (Mother ship: KEXUE),and many of them were classified as new taxa. In this paper, 13 species belonging to 11 genera from the samples collected during these surveys are described and illustrated. A review on gastropod biodiversity research in the hydrothermal vent, cold seep, and seamount are presented. The results can provide important basic data for studying the biodiversity and zoogeography of deep-sea organisms.
Gastropoda; hydrothermal vent; cold seep; seamount; prospect; from shallow water to deep sea
Q959
10.12036/hykxjk20170710001
中国科学院战略性先导科技专项A类(XDA11030201, XDA11030401); 国家自然科学基金(41636167, 41606162)
张素萍, 女, 研究员, 主要从事海洋贝类分类学研究, Email: museum@qdio.ac.cn
2017-07-10,
2017-08-01
