黄粉虫虫粉对球孢白僵菌孢子粉产量、质量及其毒力的影响
2018-01-06陶淑霞薛丹肖悦田径
陶淑霞+薛丹+肖悦+田径


摘要: 球孢白僵菌(Beauveria bassiana)在营养贫乏的培养基质上,菌株易发生退化,为保持高毒力优良菌株的稳定性,采用固液双相法生产球孢白僵菌分生孢子粉,在大米固态培养基中加入不同剂量的黄粉虫虫粉,研究添加虫粉对孢子粉产量、质量及其对大豆食心虫(Leguminivora glycinivorella)毒力的影響。结果表明,添加虫粉对球孢白僵菌的产粉量具有明显的促进作用,其中5.0+处理组的产粉量达到46.88 mg/g;添加虫粉对孢子粉活孢率无显著影响,但各处理组间的萌发速率不同,其中5.0+处理组的孢子萌发率达到50%、90%所需的时间分别为13.86、20.77 h,明显短于对照组。5.0+处理组的孢子粉对大豆食心虫毒力最高,接种后第10天的校正死亡率达到98.27%,侵染率为96.67%,致死中时(lethal time of 50%,简称LT50)为5.58 d。说明添加虫粉可以明显提高大米固态发酵生产球孢白僵菌的孢子粉产量和质量,有利于保持高毒力优良菌株的稳定性。
关键词: 球孢白僵菌;黄粉虫虫粉;孢子粉;毒力;大豆食心虫;微生物防治
中图分类号: S182 文献标志码: A
文章编号:1002-1302(2017)22-0114-03
球孢白僵菌(Beauveria bassiana)作为一种重要的生物防治介体,由于它对环境无污染、对人和动物无毒害、杀虫谱广、致病性强,从而成为目前国内外应用最广泛的昆虫病原真菌。球孢白僵菌是害虫微生物防治中的一种经典丝孢类昆虫病原真菌,可侵染15个目149个科的700多种昆虫和若干种螨类。在自然界中,它具有分布范围广、感染力强、易于扩散流行的特点,因此被认为是具有开发潜力的生防真菌 。
随着球孢白僵菌的产业化发展,人们对高质量、高纯度球孢白僵菌分生孢子的需求量越来越大。生产上面临着由菌株退化而导致的防效不稳、高毒力菌株多代移植后毒力退化的问题[3]。因此在实际生产中控制菌株退化是十分必要的。长期在营养贫乏的培养基质上培养球孢白僵菌容易导致菌株产生退化现象[4]。因此,根据目前球孢白僵菌生产和应用的现状,亟待解决的问题之一是如何保持高毒力优良菌株的稳定性。
黄粉虫是一种高蛋白、高脂肪、氨基酸含量较全面的资源昆虫。本试验选用经毒力测定对大豆食心虫具有高毒力的球孢白僵菌Bb02作为供试菌株,采用固液双相法生产球孢白僵菌分生孢子粉,在大米固态培养基中加入不同剂量的黄粉虫虫粉,研究添加虫粉对孢子粉含孢量、活孢率、孢子萌发速率及其对大豆食心虫毒力的影响,以求证添加虫粉对孢子粉质量、产量及对大豆食心虫毒力的影响,为今后球孢白僵菌孢子粉的工业化生产提供必要参考。
1 材料与方法
1.1 供试菌株
试验中所选用的球孢白僵菌为分离自大豆食心虫越冬幼虫且经过筛选对其具有高毒力的Bb02菌株。
1.2 供试昆虫
大豆食心虫采自吉林省长春市吉林农业大学植物保护基地大豆田,自然脱荚的越冬老熟幼虫。选择健康、整齐一致的幼虫用于试验。黄粉虫由吉林农业大学昆虫实验室提供,取新鲜健康的黄粉虫蛹,微波烘干,研磨成粉备用。
1.3 球孢白僵菌孢子粉生产
1.3.1 液体培养
采用固液双相法生产球孢白僵菌分生孢子粉,应用SDY培养基(成分:葡萄糖为40 g,酵母浸出粉为10 g,蛋白胨为10 g,水为1 000 mL)进行母液和子液培养。将筛选出的高毒力菌株Bb02的孢子粉接入20 mL的SDY培养基中,在25 ℃,130 r/min摇床中培养48 h后,取1 mL母液加入到灭菌后的100 mL SDY培养基中,在25 ℃,130 r/min摇床中进行子液培养24 h。
1.3.2 固体培养
每个保鲜袋盛装100 g大米,拌入0.5 mL豆油,分别添加虫粉1.0、2.5、5.0 g,替代相应等量的大米,以不添加虫粉为对照,各处理分别用1.0+、2.5+、5.0+、CK表示,每个处理重复3次。经湿热灭菌至适当熟度,取子液接入其中并进行下一步培养,接种量为13%(体积/质量),充分混合后铺平,将袋口边折起。在温度为25 ℃、相对湿度为75%的条件下培养14 d。将发酵物置于超净工作台上自然晾干后,用200目标准筛收集孢子粉。对收集的孢子粉进行质量检测。
1.4 球孢白僵菌产粉量和孢子粉质量的测定
产粉量:每袋大米培养物上收获的孢子粉质量除以大米质量即为产粉量(mg/g)。
含孢量:称取孢子粉1 mg加入到50 mL 0.05%吐温-80溶液中,经磁力搅拌器充分搅拌(500 r/min)后镜检单孢率为100%时,用血球计数板测定孢子悬浮液浓度(个/mL),计算1 g孢子粉所含的孢子个数,即为含孢量(个/g)。
含水量:将称量瓶烘干称质量,加入约100 mg孢子粉,将称量瓶和孢子粉在100 ℃下干燥2 h后取出称质量,计算含水量(%)。
活孢率:取适量孢子粉加入至50 mL SDY培养基中,配成浓度为1.0×106个/mL的孢子悬浮液,在25 ℃条件下,130 r/min培养24 h,镜检萌发率,即为孢子粉的活孢率(%)。孢子萌发的标准为芽管长度等于或大于孢子直径[5]。
1.5 球孢白僵菌孢子萌发速率测定
分别取1.0+、2.5+、5.0+、CK组孢子粉适量,加入 50 mL SDY培养基中,配成浓度为1.0×106个/mL的孢子悬浮液,在25 ℃,130 r/min摇床中培养。分别在培养6、12、18、24、30 h时,用微量移液器吸取20 μL菌液滴在血球计数板上,然后置于显微镜下进行观察,测定孢子的萌发率,每个处理重复3次。
1.6 球孢白僵菌芽生孢子浓度的测定
将上述用于测定孢子萌发率的菌液继续培养至48 h时观察芽生孢子,分别在培养48、72、96、168 h时,用微量移液器吸取20 μL 培养液,加入到380 μL无菌水中进行稀释。然后用微量移液器吸取20 μL滴在血球计数板上,然后置于显微镜下进行观察,測定芽生孢子浓度(个/mL),每个菌株3次重复。
1.7 球孢白僵菌孢子粉毒力测定
分别称取各处理组孢子粉与灭菌土(过50目筛)充分混匀,使1 g菌土中含有1.0×107个分生孢子。将含水量为20%的灭菌土壤盛于塑料杯(8 cm×12 cm)中,然后在表面均匀洒上配制的菌土,每个杯子接入20头健康的大豆食心虫老熟幼虫,使 之在菌土上自由爬行,待幼虫钻入土壤后,将杯口用保鲜膜封好,放于25 ℃培养箱中,每个处理重复3次,分别在接种后第4、6、8、10天调查幼虫死亡情况,同时捡出死虫将其保湿培养。观察球孢白僵菌侵染情况,统计幼虫死亡率、校正死亡率和侵染率,并计算致死中时(lethal time of 50%,简称LT50)。
1.8 数据分析
采用SPSS 19.0软件进行数据分析;单因素方差分析(One-Way ANOVA)检验各虫粉添加组孢子粉的产量、质量及其对大豆食心虫毒力的差异显著性(P<0.05); 用回归分析中曲线估计拟合孢子粉的萌发率随培养时间变化的过程;用回归分析中Probit分析各处理组孢子粉对大豆食心虫的LT50。
2 结果与分析
2.1 添加虫粉对球孢白僵菌孢子粉产量和质量的影响
由表1可知,各处理组间的球孢白僵菌产粉量具有显著差异(F3,11=175.299,P<0.000 1),添加虫粉对球孢白僵菌产粉量具有明显的促进作用,其中5.0+组的球孢白僵菌产粉量高达46.88 mg/g,对照组的球孢白僵菌产粉量只有1862 mg/g。各处理组的含水量(F3,11=0.03,P=0.993)和含孢量(F3,11=0.277,P=0.841)均无显著差异。虫粉添加组[CM(25]的活孢率随着虫粉添加量的增加而增加,但未达到显著差
异水平(F3,11=1.512,P=0.284)。
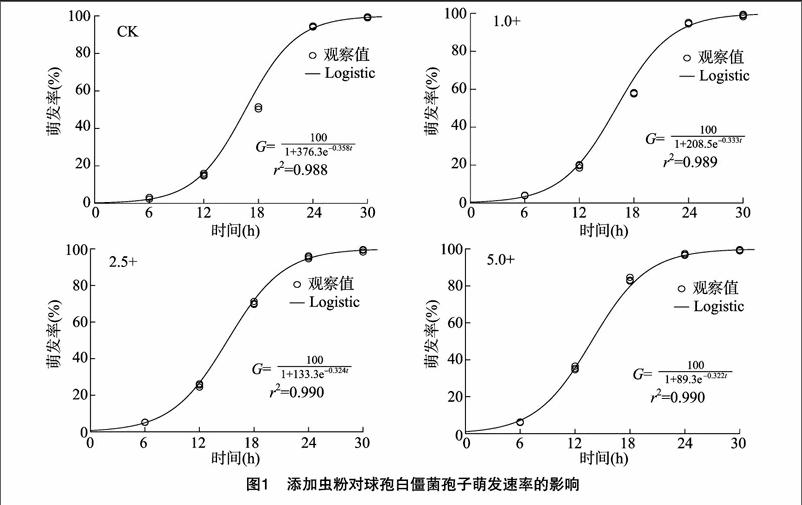
2.2 添加虫粉对球孢白僵菌孢子萌发速率的影响
各处理组的球孢白僵菌孢子萌发率观察值如图1所示,添加组和对照组球孢白僵菌孢子粉在第24 h的萌发率差异并不显著(F3,11=1.512,P=0.284)。用Logistic方程 A为最大观测值的渐近值;B为常数;k为增长率)拟合球孢白僵菌孢子粉的萌发率(G)随培养时间(t)变化的过程,拟合结果较好[6]。根据拟合模型计算球孢白僵菌孢子萌发率达到50%、90%所需的时间,5.0+组分别为13.86、20.77 h,2.5+组别为15.10、21.88 h,1.0+组分别为16.04、22.63 h,对照组分别为16.57、22.70 h。虽然添加组和对照组球孢白僵菌孢子粉的最终萌发率是一致的,但5.0+组的孢子萌发速度最快,对照组孢子的萌发速度最慢,表明添加虫粉对球孢白僵菌孢子的萌发速率具有促进作用。
2.3 添加虫粉对球孢白僵菌芽生孢子浓度的影响
由表2可知,培养48 h后5.0+组的球孢白僵菌芽生孢子浓度为 1.24×106个/mL,显著高于其他组(F3,11=11.786,P=0.03),而2.5+、1.0+组与对照组差异不显著。在培养72 h(F3,11=74.115,P<0.000 1)、96 h(F3,11=39.195,P<0.000 1)、168 h(F3,11=125.061,P<0.000 1)时,各处理组间球孢白僵菌芽生孢子浓度存在显著差异,芽生孢子浓度随着虫粉添加量的增加呈现显著的上升趋势,表明添加虫粉对Bb02菌株液体发酵芽生孢子浓度有明显的促进作用。
2.4 添加虫粉对球孢白僵菌孢子粉毒力的影响
由表3可知,与对照组相比,5.0+组的球孢白僵菌孢子粉对大豆食心虫的校正死亡率(P=0.022)、侵染率(P=0046)有显著影响。5.0+组对大豆食心虫毒力最高,接种后第10天的校正死亡率达到98.27%,侵染率为96.67%,LT50为 5.58 d,而对照组接种后第10天的校正死亡率为9138%,侵染率为8833%,LT50为6.64 d,可见添加虫粉能明显提高球孢白僵菌对大豆食心虫老熟幼虫的毒力。
3 讨论
虫生真菌在继代培养过程中会发生高频率的退化现象,表现为菌落角变、气生菌丝化、分生孢子产生能力下降或丧失、重要次生代谢产物合成能力下降或丧失,往往给虫生真菌的工业化生产及应用带来重大损失[7-8]。球孢白僵菌的高度变异可引起生产菌种的退化,从而导致毒力降低,严重影响防治效果。因此在生产和应用中,选育高毒力菌株的同时,应采取有效措施控制变异。在营养贫乏的培养基质上,球孢白僵菌菌株易发生变异,为保持高毒力优良菌株的稳定性,应选择适宜的培养基质[9]。
王福祥等在培养球孢白僵菌的PDA培养基中加入黄粉虫幼虫发现,与对照相比,添加幼虫的处理组在营养生长量、孢子萌发率、产孢量和酶活性等方面均有所提高[10],但在培养基上的产孢状况和应用中的规模化生产还存在着很大差异[11],所以还不能充分反映生产中的实际产孢情况。因此,本试验将黄粉虫虫粉添加到大米固态培养基中,采用固液双相法对供试菌株进行适当规模的扩大生产。结果表明,添加虫粉可以显著提高大米固态发酵生产球孢白僵菌的孢子粉产量和质量,有利于保持高毒力优良菌株的稳定性。
目前,球孢白僵菌孢子粉的活孢率以24 h的萌发率表示[12]。然而,活孢率相同的孢子粉并不意味着其孢子的萌发进程一致[13]。本研究在培养基质中添加黄粉虫虫粉虽未影响球孢白僵菌孢子粉的活孢率,但与对照相比,球孢白僵菌孢子萌发率达到50%、90%所需的时间却明显缩短,表明添加黄粉虫虫粉可能影响球孢白僵菌孢子恢复活力的进程或吸水萌发的速度,与对照相比,对大豆食心虫的毒力也有所增加,这种差异正好从孢子萌发速度差异的比较中得到合理解释,即萌发速度越快的孢子毒力越强。由此印证,任何影响孢子萌发的因素都将影响到孢子毒力[14]。
稳定并提高虫生真菌的毒力对发展有效的商品制剂至关重要。虫生真菌入侵寄主的过程与其对体壁结构物质分解酶的活性有关,几丁质酶和蛋白降解酶在虫生真菌入侵寄主体壁的过程中起重要作用,其活性高低是决定真菌侵染能力的重要因素[15-16]。虫生真菌的几丁质酶和蛋白降解酶都具有诱导性。研究表明,不同的真菌培养基也影响真菌毒力。Milner发现,粉擬青霉菌株对麦长管蚜(Sitobion avenae)的毒力在蚜虫或含蚜虫表皮的琼脂板上传代3代后LT50从11.1 d下降到 5 d[17]。Pinto等发现,黄绿绿僵菌(Metarhizium flavoviride)蛋白降解酶的活性在该菌的培养液中存在,但当有昆虫表皮存在时能高度表达[18]。本研究在培养基质中加入虫粉显著提高球孢白僵菌对大豆食心虫的毒力,也进一步表明加入昆虫物质可诱导虫生真菌毒力的提高,说明毒力因子易受培养底物的诱导,为提高真菌毒力提供了有价值的途径。
参考文献:
[1] 林乃铨. 害虫生物防治[M]. 北京:科学出版社,2010,238.
[2]Feng M G,Poprawski T J,Khachatourians G G. Production,formulation and application of the entomopathogenic fungus Beauveria bassiana for insect control:current status[J]. Biocontrol Science and Technology,1994,4(1):3-34.
[3] 唐晓庆,樊美珍,李增志. 球孢白僵菌继代培养中菌落局变现象及环境影响因素的研究[J]. 真菌学报,1996,15(3):188-196.
[4]蔡国贵,林庆源,徐耀昌,等. 白僵菌菌株退化与培养条件关系及其控制技术[J]. 福建林学院学报,2001,21(1):76-79.
[5]Hong T D,Gunn J,Ellis R H,et al. The effete of storage environment on the longevity of conidia of Beauveria bassiana[J]. Mycological Researeh,2001,105(5):597-602.
[6]董江水. 应用SPSS软件拟合Logistic曲线研究[J]. 金陵科技学院学报,2007,23(1):21-24.
[7]张正坤,孙召朋,张语迟,等. 继代培养对球孢白僵菌毒素产生水平的影响[J]. 中国农学通报,2012,28(24):243-249
[8]王成树,吕丁丁,李 琳. 虫生真菌致病与退化机理研究[C]//中国菌物学会第四届会员代表大会暨全国第七届菌物学学术讨论会论文集. 湖北武汉,2008:10-17.
[9]樊美珍,李增智,唐晓庆. 白僵菌菌种退化及其控制[J]. 安徽农业大学学报,1996,23(3):239-245.
[10] 王福祥,温晓蕾,王长青,等. 黄粉虫虫粉对白僵菌生长的影响[J]. 安徽农业科学,2010,38(18):9575-9576,9596.
[11]Quesada-Moraga E,Maranhao E A A,Valverde-Garcia P,et al. Selection of Beauveria bassiana isolates for conrol of the whiteflies Bemisia tabaci and Trialeurodes vaporariorum on the basis of their virulence,thermal requirements,and toxicogenic activity[J]. Biological Control,2006,36(3):274-287.
[12]殷凤鸣,潘务耀,李增智. 白僵菌生产企业标准[J]. 安徽农业大学学报,1996,23(3):321-325.
[13]应胜华,冯明光. 真空干燥球孢白僵菌纯孢粉的活孢率、毒力与贮存期[J]. 微生物学通报,2002,29(5):42-47.
[14]Anderson T E,Roberts D W. Compatibility of Beauveria bassiana isolates with insecticide formulations used in colorado potato beetle (Coleoptera:Chrysomelidae) control[J]. Journal Economic Entomology,1983,76(6):1437-1441.
[15]Wang C S,Hu G,st Leger R J. Differential gene expression by Metarhizium anisopliae growing in root exudate and host(Manduca sexta)cuticle or hemolymph reveals mechanisms of physiological adaptation[J]. Fungal Geneties and Biology,2005,42(8):704-718.
[16]Robert A,Messing-AI-Aidfroos K. Acid production by Metarhizium anisoplia:effects on virulence against mosquitoes and on detection of invitro amylase,protease and lipase activity[J]. Journal of Invertebrate Pathology,1985,45(1):9-15.
[17]Milner R J. Prospects for biopesticides for aphid control[J]. Entomophaga,1997,42(1/2):227-239.
[18]Pinto F G S,Fungaro M H P,Ferreira J M,et al. Genetic variation in the cuticle-degrading protease activity of the entomopathogen Metarhizium flavoviride[J]. Genetics and Molecular Biology,2002,25(2):231-234.
