不同林龄马尾松林下植物多样性与环境特征
2018-01-06陈小红赵安玖
陈小红,赵安玖,张 健,曾 洪
(四川农业大学林学院,四川温江 611130)
不同林龄马尾松林下植物多样性与环境特征
陈小红,赵安玖,张 健*,曾 洪
(四川农业大学林学院,四川温江 611130)
【目的】分析不同林龄马尾松(Pinus massoniana)林下植物多样性变化及其与环境特征的关系,探求影响物种更替的环境因子,以期为马尾松人工林的生态系统管理提供参考。【方法】采用空间代替时间的方法,对不同林龄(幼龄林、中龄林和成熟林)的马尾松林下植物多样性特征进行了研究,环境因子与植物物种的相关性采用典范对应分析(Canonical Correspondence Analysis,CCA)来实现。【结果】早期阶段(幼龄林和中龄林)的马尾松林下物种丰富,到后期(成熟林)林下仅被少量主要物种占据;随着演替的进行,物种多样性指数E、D、H明显降低,草本层表现尤为明显,木本层不显著;林分结构(林分密度、胸径、树高)对草本层植物多样性影响明显,而对木本层的影响相对较弱;土壤容重与幼龄林、成熟林的物种多样性指数显著相关;土壤全氮、全钾、有机质对初期植物多样性的影响显著,特别是草本植物。【结论】随着林龄的延长,马尾松林下植物多样性指数明显降低。林分结构和土壤质量是马尾松林下植物多样性变化的主要影响因子。
林分结构;土壤养分;物种多样性;年龄序列;马尾松人工林
植物群落是植物与环境相互作用的产物,群落演替是物种扩散和环境筛综合作用的过程[1]。随着植物群落演替进行,群落结构、物种组成和群落环境都表现出分异格局,同时又相互依存。林下植被通过资源和位置的占有及其生命活动,不断改变着林下微环境,从而对整个森林生态系统的稳定、演替发展和植物多样性起着重要作用[2-4]。同时,不同的林分结构形成了不同的生态条件以适应不同的物种生存,从而影响着植物多样性的变化[1-2]。在外界环境因子作用下形成土壤养分格局后,土壤对植物的生长势、胸径、株高、冠幅、覆盖度等造成直接影响,影响着植物群落的组成结构和演替进程[5-7]。因此,研究植物多样性与环境特征,能够有效地反映各物种在群落中的分布情况及其对环境因子的适应程度,有助于进一步认清群落结构和演替趋势。
植物多样性与环境特征是人工林生态系统稳定性的基础,是人工林经营管理的重要目标[8]。以单一树种为主的人工林生态系统中,乔木层植物种类单一,结构简单,养分分解归还速率慢,林下植被的作用就更为重要,其林分结构、土壤肥力与林下植物多样性的关系引起了众多学者的广泛关注[9-12],加强人工林合理经营与调控已成为实现人工林可持续经营亟待解决的科学问题。马尾松(Pinus massoniana)是我国亚热带地区特有的乡土树种,也是主要造林树种中分布最广、亚热带东南部湿润地区典型的针叶代表种,在我国生态环境建设和林业产业中具有十分重要的作用[13]。
本研究以不同林龄阶段的马尾松人工林为研究对象,采用空间代替时间的方法,分析不同林龄马尾松林下植物多样性变化及其与环境特征的关系,探求影响物种更替的环境因子,以期为马尾松人工林的生产力维护及生态系统管理提供一定参考,推动人工林经营的可持续发展。
1 材料和方法
1.1 研究区概况
研究区位于宜宾市高县来复镇国营林场的马尾松林区,地理坐标为 104°33′37″~104°34′22″E,28°35′47″~28°36′19″N,海拔高度为 417~538 m,地处四川盆地南缘,属中亚热带湿润季风性气候,年平均气温18.1℃,降水量1 021 mm,无霜期360 d。
马尾松林为单一的马尾松人工纯林,造林时均为2 m×2 m株行,距林分年龄为2~39 a。选取地形地貌、海拔、母岩、土壤类型等相同或相近的不同林龄马尾松人工林为研究样地。马尾松人工林林下植被主要为油樟(Cinnamomum longepaniculatum)、梨叶悬钩子(Rubus pirifolius)、巴豆(Croton tiglium)、野桐(Mallotus tenuifolius)、展毛野牡丹(Melastoma normale)、马唐(Digitaria sanguinalis)、芒(Miscanthus sinensis)、蕨(Pteridium aquilinum var.latiusculum)、芒萁(Dicranopteris pedata)和荩草(Arthraxon hispidus)等。各样地基本情况见表1。
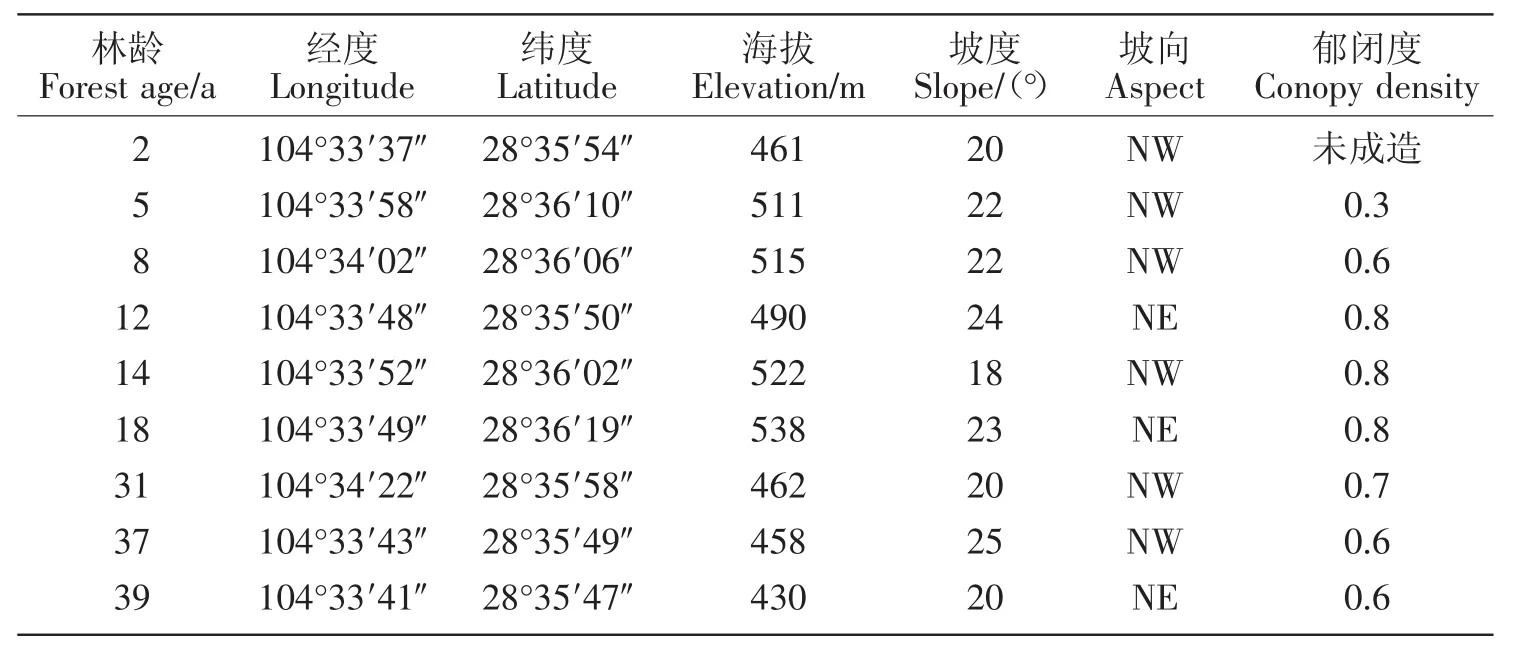
表1 样地设置及林分状况Table 1 Sample plot and forest condition
1.2 研究方法
选择立地条件相近、人为干扰较小的不同林龄马尾松林进行调查,每个年龄林分设置3个20 m×20 m的乔木样方调查乔木树种,3个5 m×5 m和5个1 m×1 m的小样方分别调查灌木和草本植物。在每个样方中记录物种种类、盖度、数量、高度、乔木的胸径和冠幅等,同时记录样方的地理坐标、海拔、坡度、坡向、坡位、土壤类型、林分郁闭度等立地因子。在每个样方的四角、中央5个点挖取0~20 cm的土壤,混匀后带回实验室风干,测定土壤容重、含水量、pH、有机质、全氮、速效氮、全磷、速效磷、全钾和速效钾[14]。
通过野外调查数据,根据马克平所评述的植物群落多样性测度方法[15],测度指标是分层各物种的重要值(important value,IV),按照下列公式计算各α多样性指数:

式中,S为物种数,即物种丰富度;Pi=Ni/N,Ni为第i个物种的重要值,N为各层所有物种的重要值之和。
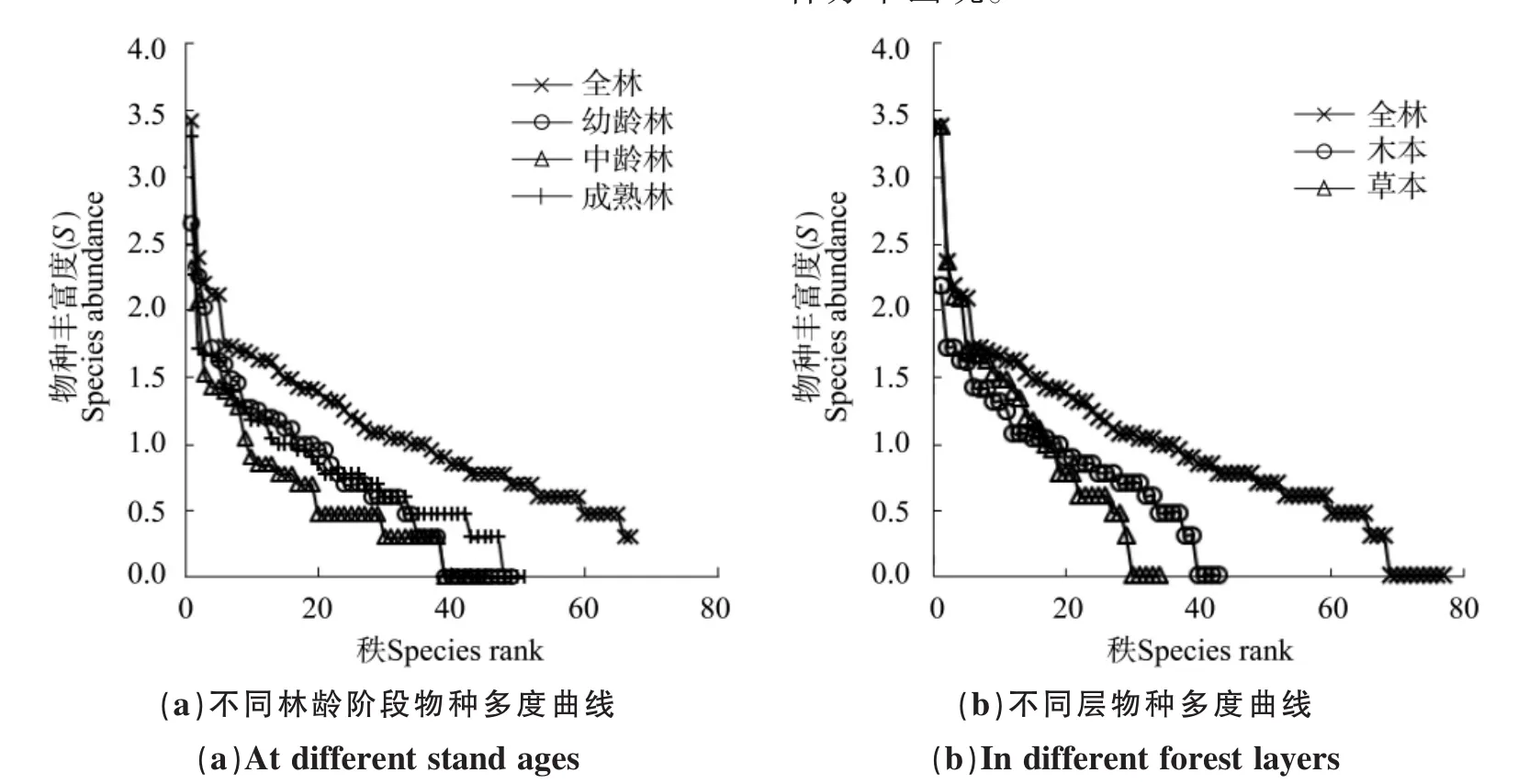
图1 物种秩-多度曲线Figure 1 Relationship of the abundance of species (logarithmic) and species rank
环境因子与植物物种的相关性采用PC-ORD 5.0中的典范对应分析(CanonicalCorrespondenceAnalysis,CCA)来实现,其他相关数据处理通过SPSS 19.0软件完成。
2 结果与分析
2.1 不同林龄马尾松林下物种组成
在所有样方中,马尾松人工林下出现的植物种类为43科70属77种,其中木本植物25科38属43种;草本植物20科32属34种。山茶科(Theaceae)、大戟科(Euphorbiaceae)、豆科(Leguminosae)、蔷薇科(Rosaceae)和樟科(Lauraceae)植物分布较多,芒、芒萁、裂叶榕(Ficus laceratifolia)、野桐、铁仔(Myrsine africana)在每个样地中均有出现。
物种秩-多度曲线表明(图1),成熟林的曲线变化较平缓,其林下植物均匀度相对高些,以梨叶悬钩子、展毛野牡丹、茶(Camelia sinensis)和蕨类植物(Pteridophyta)为主。幼龄林、中幼龄林的均匀度相对低些,中龄林曲线变化更加急剧,中龄林下植物除枹栎(Quercus serrata)、野桐占有绝对优势(IV>25%)以上外,其他植物的重要值大小分布较均衡,彼此差异不大。木本层相对草本层有较高的均匀度,其物种秩-多度曲线变化平缓,在不同林龄马尾松林下梨叶悬钩子、展毛野牡丹、野桐、黄荆(Vitex negundo)、楤木(Araliachinensis)、枹栎和盐肤木(Rhus chinensis)等木本植物占有较大的优势,而草本植物种类较多,且分布不均匀,有些植物如白酒草(Conyza japonica)、紫花地丁(Viola philippica)、梁子菜(Erechthites hieracifolia)、狗脊蕨(Woodwardia japonica)等只在个别林分中出现。
2.2 不同林龄马尾松林下植物多样性
由表2和图2可知,不同林龄马尾松林下草本层的多样性指数E、D和H总体差异明显(P<0.01),并随年龄的增加而降低,成熟林的林下植物种类较单一,且分布不均,其多样性指数明显低于幼龄林和中龄林。不同林龄阶段全林和木本层的多样性指数差异均不明显(P>0.01)。
幼龄林中木本层物种多样性指数E与草本层E、D和H呈显著负相关,并在林分年龄增加的过程中,相关性越来越低。随着林分年龄增加,木本层多样性指数H与草本层E、D和H的相关性随之发生变化,到成熟林时表现为显著正相关(表3)。
2.3 环境因素对马尾松林下物种多样性的影响
不同林龄阶段的植被群落,其环境条件(林分结构与土壤营养)也发生着变化。由表2可知,不同林龄马尾松林之间的林分密度、树高、胸径差异性显著(P<0.01)。土壤养分速效钾、全磷之间亦呈现显著差异(P<0.01),从幼龄林到成熟林土壤养分含量呈增加趋势,土壤肥力得以提高。
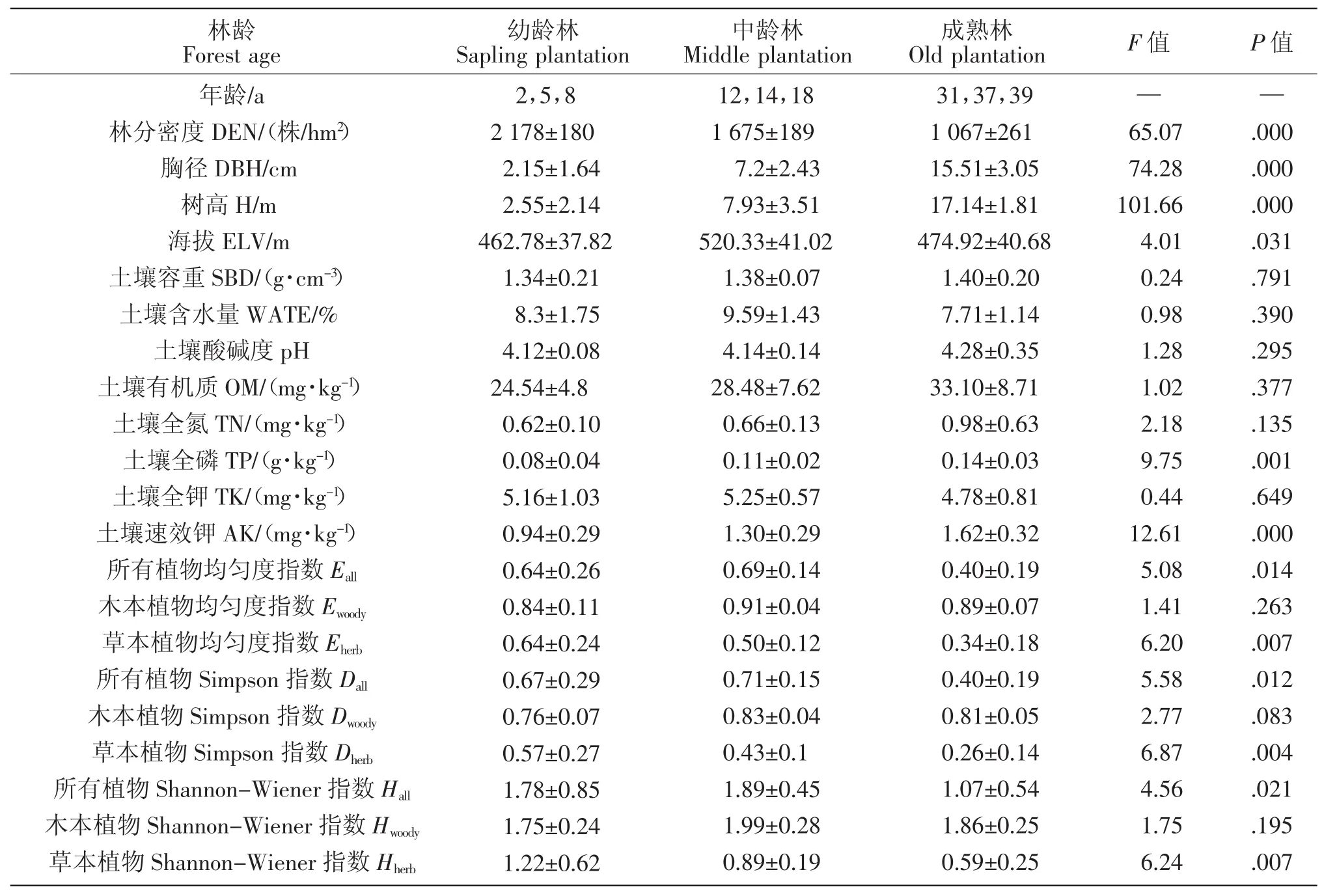
表2 马尾松林下植物多样性与环境因子特征(平均值±标准误)Table 2 Description of plant diversity and environmental factors(mean value±standard error)
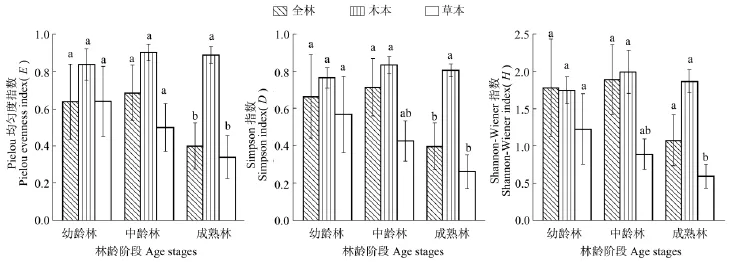
图2 不同林龄阶段物种多样性指数E、D和H分布图Figure 2 Changes of species diversity index E,D and H
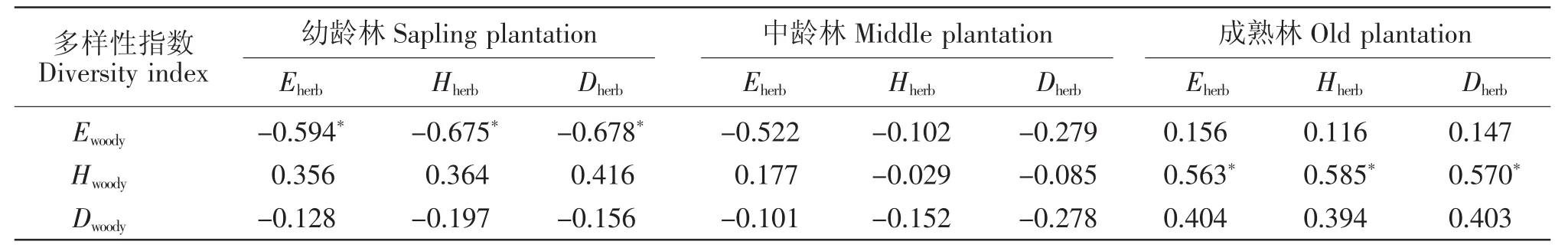
表3 不同林龄阶段木本层多样性与草本层多样性之间的相关性(Sperman相关系数)Table 3 Correlations between the diversities of herb layer and woody layer(Sperman's correlation coefficients)
随着环境条件的改善,林下植物种也发生着变化。由表4、表5和图3可看出,环境因子对物种的影响由CCA的第一轴(Axis1)决定,林分环境对各层物种组成的解释变量分别达23%、20.3%、23.7%,环境因子对植物多样性的影响十分显著,影响着植物种在各林龄下的分布和重要值,各物种在CCA排序图上分异显著,占据着不同的生态位。林分密度与物种组成呈正相关,与树高、胸径、海拔及土壤速效钾、全磷呈负相关,由此看出林分结构(密度、树高、胸径)、土壤特性(土壤速效钾、全磷)等环境因子是物种组成的重要影响因素。
就所有马尾松林分而言,林分结构(密度、树高、胸径)和土壤容重对植物多样性的影响极显著(P<0.01),对草本层的影响甚为明显,木本层植物多样性仅受土壤速效钾含量的影响。幼龄林阶段土壤全氮、全钾含量与植物多样性指数高度相关(0.01<P<0.05),有机质含量、海拔高度与多样性指数E、D和H之间的相关性达到了极显著(P<0.01)。中龄林阶段土壤全氮对草本层多样性的影响较大(0.01<P<0.05),而到成熟林阶段土壤容重与草本层多样性指数的关系密切(P<0.01),土壤速效钾、全磷含量对木本层植物多样性影响显著(P<0.01)。
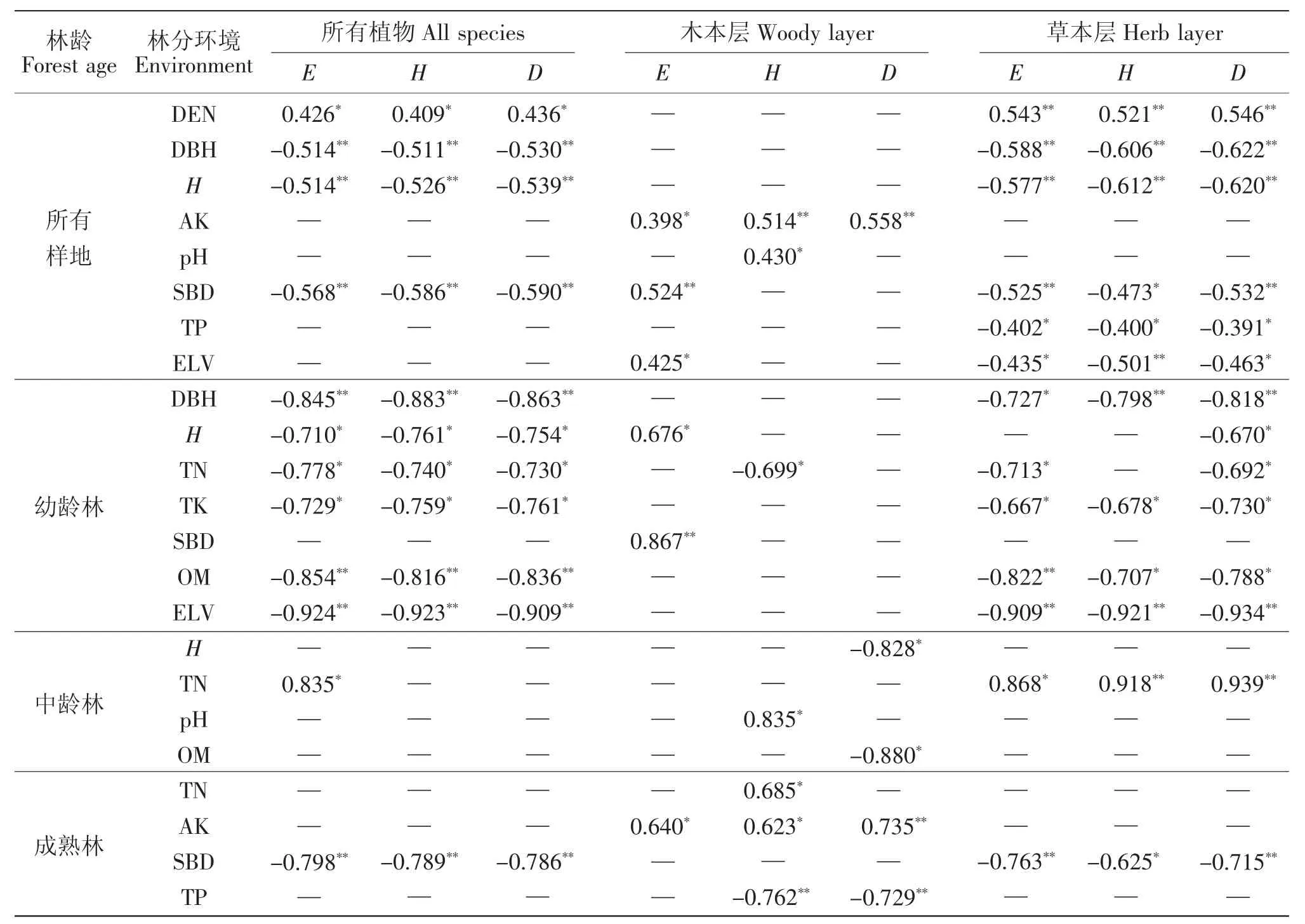
表4 植物多样性指数与环境变量的相关分析(Sperman相关系数)Table 4 Sperman's correlation coefficients between environmental parameters and diversity indexes
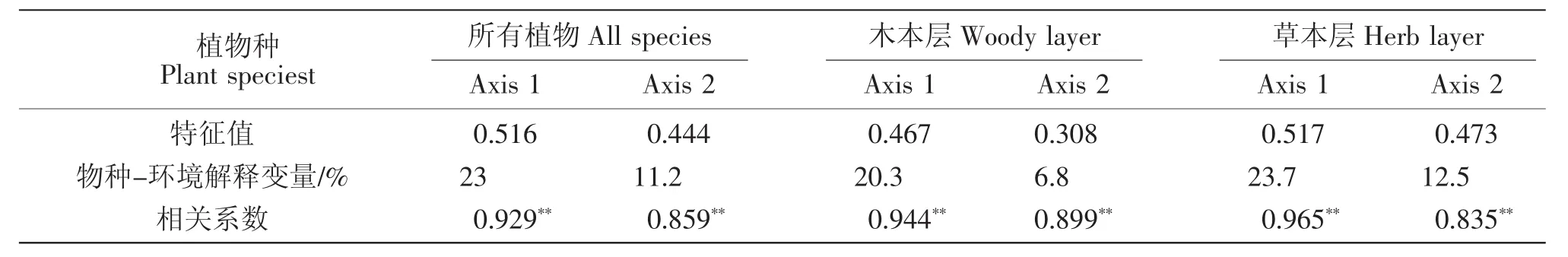
表5 植物种与环境因子的CCA分析特征Table 5 Summary of the CCA analysis of all species,woody-layer and herb-layer vegetation
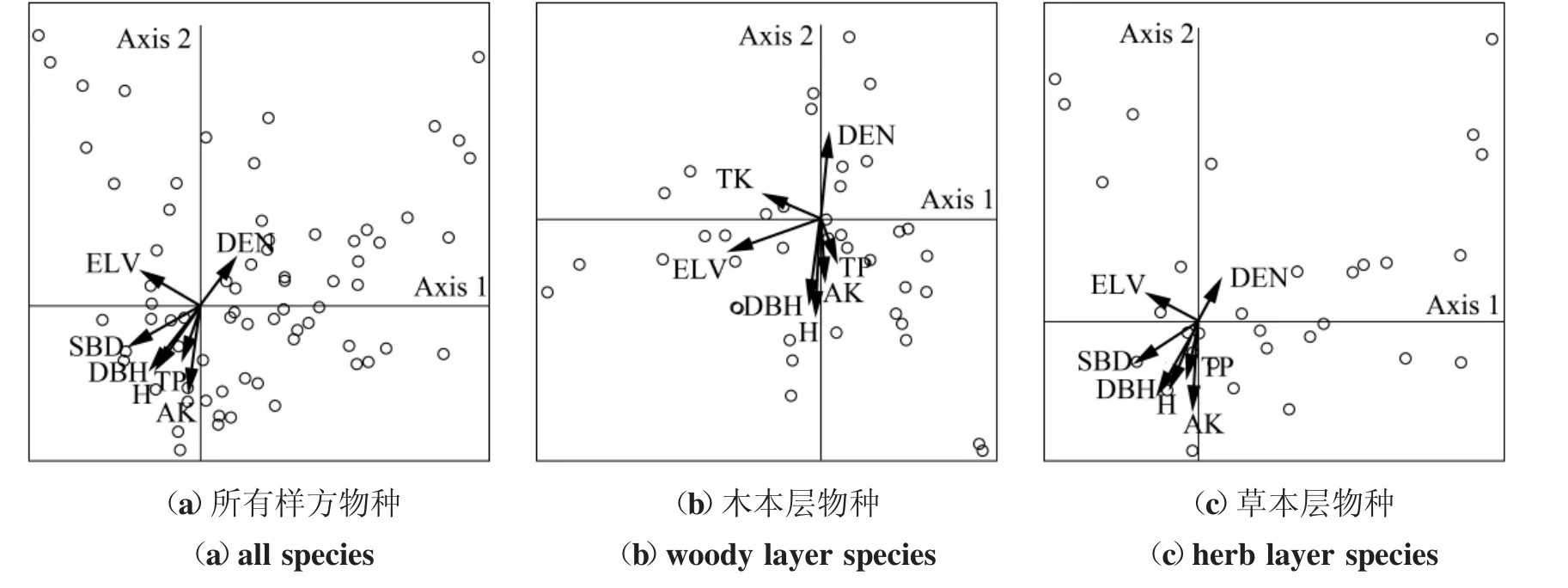
图3 植物种与环境因子的CCA关系图Figure 3 CCA graph of plant species and environmental variables
3 讨论
3.1 马尾松人工林林下植物多样性
马尾松人工林在发育过程中造成栖息地的异质性,不同阶段的植物多样性表现不同。物种变化主要发生在早期演替阶段,幼龄林(2~8 a)林下物种数表现为先下降后逐渐升高的趋势,中龄林后随着时间的推移,群落中草本层的多样性指数基本上逐渐降低,木本层的变化规律不明显,成熟林(31~39 a)林下草本层中芒和芒萁占有绝对优势,其他植物种类明显减少。不同林龄的桉树[16-17]、杉木[18]等人工林亦发现林下植物多样性存在明显差异和波动性,在演替阶段后期,物种多样性有降低的趋势,原因在于植物多样性除了受海拔、土壤、坡位、坡向、坡度、地形等环境因子影响外,也与林分郁闭度、林龄和群落结构等密切相关,而与人工林造林树种无关[18]。因此,在营林过程中应合理调整人工林的林分结构,以增加林下灌草的物种多样性。
木本层和草本层的多样性之间存在一定相关性,在幼龄林、成熟林表现更明显,表明除优势树种马尾松外,灌木层在一定条件下形成生境条件的异质性,在整个林分中有助于维持一个恒定水平草本层的多样性[19-22]。在幼龄林阶段,常见灌木为悬钩子、野桐、枹栎等,灌木层生境特征表现为一个具有更高生物活性的凋落物,尤其是在马尾松林树冠还没完全展开时,大量侵入的光源、水分和较高的土壤温度,增加了凋落物分解速率,使得草本植物得以及时出现。在成熟林阶段,灌木层的生境已形成,一定程度上对草本层生境产生影响,林下出现了一些耐荫植物,其他不能适应遮荫环境的物种则被淘汰。
3.2 林分环境对林下植物多样性的影响
林分结构对林下植物多样性的影响十分明显,CCA分析表明,林分密度与物种多样性指数E、H、D呈正相关,而与胸径、树高呈负相关。这表明林分结构(林分密度、胸径、树高)能改变局部环境,同时作为功能群的重要过滤器,能很好地适应局部立地条件,对灌木、草本植被结构塑造具有重要性[23-24]。在马尾松林演替阶段中后期,林分结构变化大,林冠郁闭度较早期增加,光照较难进入林下,同时芒、芒萁等植物在酸性土上有着极强生存力和竞争力,导致后期植物多样性有所降低。本研究中林分密度、胸径与物种多样性变化息息相关,是较林分年龄更能反映演替进程中物种多样性变化的理想指数,这与大多数研究结果一致[25-28]。
植物多样性变化不仅取决于林分结构,土壤质量同样是影响不同演替阶段物种组成的主要环境因子[1]。植物群落中土壤限制植物种群的生长,土壤营养与物种多样性密切相关[12],但不同研究对象其研究结论各异,有的研究认为生物多样性随着土壤营养的增加而降低[29];有的则发现生物多样性起初随着土壤营养的增加而提高,然后降低[30-31];还有的则发现土壤营养对生物多样性没有影响[32]。本研究中土壤容重在成熟林阶段与多样性指数呈高度负相关(P<0.01),而与土壤含水量的相关性较差(P>0.05),原因可能在于研究区降雨充沛,水分不是主要限制因子。物种多样性与演替早期阶段的土壤有机质、全氮、全钾相关性显著(P<0.05),随着演替进行,由于凋落物的回归,土壤养分逐渐增加并趋于稳定,对物种多样性的影响相对减小,表明物种多样性的改变与群落最初的土壤营养条件有关,这与T.A.Theresa等[33]的研究结果一致。因此,应增加马尾松人工林的土壤肥力,适当疏伐改善林分结构,促进演替过程中的植物多样性恢复,维护生态系统的稳定性。
本研究在一定程度上揭示了林分结构和土壤因子对马尾松林下物种多样性演替的重要性,但植物群落多样性的维持机制仍然不很清楚,今后还应从林分光环境、林分微气候、土壤中各元素的生物化学循环等方面进行深入细致研究。
[1]王世雄,王孝安,李国庆,等.陕西子午岭植物群落演替过程中物种多样性变化与环境解释[J].生态学报,2010,30(6):1638-1647.
[2]龙成,吴庭天,杨小波,等.热带次生林演替过程中环境因子对物种多样性的影响[J].东北林业大学学报,2016,44(1):37-43.
[3]CARREARA A L.,MAZZARINO M J,BERTILLER M B,et al.Plant impacts on nitrogen and carbon cycling in the Monte Phytogeographical Province[J].Argentina.J.Arid.Environ.,2009(73):192-201.
[4]HOOPER D U,VITOUSEK P M.Effects of plant composition and diversity on nutrient cycling[J].Ecol.Monogr.,1998(68):121-149.
[5]OELMANN Y,WILCKE W,TEMPERTON V M,et al.Soil and plant nitrogen pools as related to plant diversity in an experimental grassland[J].Soil Sci.Soc.Am.J.,2007(71):720-729.
[6]NAGAIKE T,KAMITANI T,NAKASHIZUKA T.Plant species diversity in abandoned coppice forest in a temperate deciduous forest area of central Japan[J].Plant Ecology,2003,166(1):63-74.
[7]SITZIA T,TRENTANOYI G,DAINESEA M,et al.Stand structure and plant species diversity in managed and abandoned silver fir mature woodlands[J].Forest Ecology and Management,2012(270):232-238.
[8]温远光,元昌安,李信贤,等.大明山中山植被恢复过程植物物种多样性的变化[J].植物生态学报,1998,22(1):33-40.
[9]沈国舫.中国林业可持续发展及其关键科学问题[J].地球科学进展,2000,15(l):l0-18.
[10]盛炜彤,范少辉.人工林长期生产力保持机制研究的背景、现状和趋势[J].林业科学研究,2004,17(1):106-115.
[11]康冰,刘世荣,蔡道雄,等.马尾松人工林林分密度对林下植被及土壤性质的影响[J].应用生态学报,2009,20(10):2323-2331.
[12]叶绍明,温远光,杨梅,等.连栽桉树人工林植物多样性与土壤理化性质的关联分析[J].水土保持学报,2010,24(4):246-250.
[13]洪伟,吴承祯.马尾松人工林经营模式及其应用[M].北京:中国林业出版社,1999:5-9.
[14]中国科学院南京土壤研究所.土壤理化分析[M].上海科学技术出版社,1977:78-139.
[15]马克平.生物群落多样性的测度方法Ⅰ∶α多样性的测度方法(上)[J].生物多样性,1994,2(3):162-168.
[16]余雪标,钟罗生,杨伟东,等.桉树人工林林下植被结构的研究[J].热带作物学报,1999,2(1):66-72.
[17]温远光,刘世荣,陈放等.桉树工业人工林植物物种多样性及动态研究[J].北京林业大学学报,2005,27(4):17-22.
[18]林开敏,张文富,谢国阳,等.老龄杉木林下天然更新阔叶植被物种多样性研究[J].福建林学院学报,1997,17(4):313-317.
[19]COMELISSEN J H C,PEREZ-HARGUINDEGUY N,DIAZ S,et al.Leaf structure and defence control litter decomposition rate across species and life forms in regional floras on two continents[J].New Phytol.,1999,143:191-200.
[20]GALLARDO A,MERINO J.Leaf decomposition in 2 Mediterranean ecosystems of Southwest Spain e influence of substrate quality[J].Ecology,1993(74):152-161.
[21]BARBIER S,GOSSELIN F,BALANDIER P.Influence of tree species on understory vegetation diversity and mechanisms involved a critical review for temperate and boreal forests[J].For.Ecol.Manage.,2008(254):1-15.
[22]HOBBIE S E,REICH P B,OLEKSYUN J,et al.Tree species effects on decomposition and forest floor dynamics in a common garden[J].Ecology,2006(87):2288-2297.
[23]LEBRIJA-TREJOS E,PEREZ-GARCIA E,Lebrija-Trejos A,et al.Functional traits and environmental filtering drive community assembly in a species-rich tropical system[J].Ecology,2010(91):386-398.
[24]LEBRIJA-TREJOS E,PEREZ-GARCIA E,Lebrija-Trejos A,et al.Environmental changes during secondary succession in a tropical dry forest in Mexico[J].Journal of Tropical Ecology,2011(27):1-13.
[25]BREUGEL M,BONGERS F,MARTINEZ-RAMOS M.Species dynamics during early secondary forest succession:recruitment,mortalityandspeciesturnove[rJ].Biotropica,2007(39):610-619.
[26]CHAZDON R L,LETCHER S G,BREAGUL M,et al.Rates of change in tree communities of secondary neotropical forests following major disturbances[J].Philosophical Transactions of the Royal Society B:Biological Sciences ,2007(362):273-289.
[27]LOHBECK M,POOTER L,PAZ H,et al.Functional diversity changes during tropical forest succession[J].Perspectives in Plant Ecology,Evolution and Systematics,2012(14):89-96.
[28]汤孟平,周国模,施拥军,等.不同地形条件下群落物种多样性与胸高断面积的差异分析[J].林业科学,2007,43(6):27-31.
[29]WANG C T,LONG R J,WANG Q J,e t al.Effects of altitude on plant-species diversity and productivity in an alpine meadow,Qinghai Tibetan plateau[J].Australian Journal of Botany,2007(55):110-117.
[30]BOBBINK R.Effects of nutrient enrichment in Dutch chalk grassland[J].Journal of Applied Ecology,1991(28):28-41.
[31]LAURA G,GRACE J B.Herbivore effects on plan t species density at varying productivity levels[J].Ecology,1998,79(5):1586-1594.
[32]CARSON W P,PICKETT S T A.Role of resources and disturbance in the organization of an old field plant community[J].Ecology,1990,71(1):226-238.
[33]THERESA T A,BOWMAB W D.Nutrient availability,plant abundance,and species diversity in two Alpine Tundra communities[J].Ecology,1997,78( 6):1861-1872.
Understory Plant Diversity and Environmental Characteristics of Different Age Pinus massoniana Plantations
CHEN Xiao-hong,ZHAO An-jiu,ZHANG Jian*,ZENG Hong
(College of Forestry,Sichuan Agricultural University,Wenjiang 611130,Sichuan,China)
【Objective】The aim of the study was to explore the understory biodersity and its correlation with environmental factors in different age P.massoniana plantations.This study could also provide some references for ecosystem management of P.massoniana plantations.【Method】Understory plant diversity in different age P.massoniana plantations(young,middle-age and mature forest stands)was investigated and the reationships between plant diversity and environmental factors analyzed by canonical aorrespondence analysis(CCA).【Results】Species abundance was relatively highin the early succession stage but low in the late succession stage.The herb-layer species diversity indexes,such as evenness index(E),Simpson index(D),Shannon-Wiener index(H)significantly declined with the successtion advancement.however,no significant changes was observed in woody-layer.Stand structure variables(stand density,diameter at breast height and tree height)strongly impactd plant diversity in the herb-layer but not affect woodylayer.Soil bulk density showed a significant relationship with species diversity indexes in the young and mature stages.Soil variables,including total nitrogen,total phosphorus,soil organic matter,had significant effects on plant diversities in the early successional stage,especially for the herb layer.【Conclusion】The plant diversity index of P.massoniana plantation decreased obviously with increasing forest age.Both stand structure and soil quality were two key factors affecting plant diversity.
forest stand structure;soil nutrient;species diversity;chronosequence;Pinus massoniana plantation
S718;S791.248;S758.5+3 文献标志码:A 文章编号:1000-2650(2017)02-0186-07
10.16036/j.issn.1000-2650.2017.02.008
2017-01-25
国家重点科技攻关项目(2011BAC09B05)。
陈小红,博士,讲师,主要从事植物生理生态研究,Email:xiaohong_chen@sicau.edu.cn。*责任作者:张健,教授,主要从事森林培育研究,E-mai:sicauzhangjian@163.com。
(本文审稿:庞学勇;责任编辑:巩艳红;英文编辑:徐振锋)
