我国60年来土壤养分循环微生物机制的研究历程—基于文献计量学和大数据可视化分析
2018-01-05孙波王晓玥吕新华
孙波,王晓玥#,吕新华
(1 土壤与农业可持续发展国家重点实验室/中国科学院南京土壤研究所,南京 210008;2 中国科学院武汉文献情报中心情报部,武汉 430071)
我国60年来土壤养分循环微生物机制的研究历程—基于文献计量学和大数据可视化分析
孙波1,王晓玥1#,吕新华2*
(1 土壤与农业可持续发展国家重点实验室/中国科学院南京土壤研究所,南京 210008;2 中国科学院武汉文献情报中心情报部,武汉 430071)
【目的】土壤养分循环是土壤肥力和作物生长的基础,土壤微生物是驱动养分循环的关键因子。挖掘微生物调控土壤–植物系统养分循环功能的潜力,已成为提高农田养分资源利用效率的发展趋势。通过梳理我国土壤养分循环和土壤微生物学交互研究的发展历史和脉络,审视与国际前沿研究比较的不足之处,指出未来的研究热点,促进我国在相关领域研究水平的提高。【方法】本文利用文献计量学及大数据可视化方法定量分析了60年来土壤养分循环和土壤微生物学交互研究领域的国内外的科学文献,研究了相关领域不同时期的发展与演变过程,对比了国内外土壤养分循环和土壤微生物学交互研究发展的异同点。【主要进展】分析表明国内研究主要分为起步期 (1981~1990年),发展期 (1991~2005年) 和定型期 (2006~2016年)。起步期的研究热点分散,主要研究了红壤、水稻土和紫色土中特定微生物和酶活性的肥力功能。发展期研究热点关联度加强,主要研究了红壤、黑土、水稻土中微生物量和酶活性的肥力和环境功能。定型期研究热点间的连接度不断增强,主要研究了土壤养分循环与微生物群落结构耦合的微生物学机制。通过与国际相关研究比较,国内学者追赶国际研究可以分为起步追赶期 (1990~2005年) 和快速追赶期 (2006~2016年)。在起步追赶期,国内学者追随国际上对碳氮转化与微生物交互研究的热点,但研究热点的关键词交互网络成熟程度低,侧重对红壤的研究,缺少对根际的研究。加速追赶期,国内学者从水稻土和根际方面强化了对养分转化微生物机制研究,开展了土壤微生物网络结构和功能的研究,但需要加强土壤生物网络结构和功能的实证研究。总体上,与国外相比,国内研究相对滞后,但发展迅猛,从简单关注酶活性,发展到微生物网络结构与土壤功能的交互。同时国内研究热点的关键词交互网络日益成熟,且与国际相比表现出趋同趋势,但仍需深入研究微生物群落结构演替对养分循环的驱动机制。【结论与展望】在未来研究中,应在建立养分利用率提升的区域微生物调控技术体系的同时,不断发展和完善土壤微生物结构和功能协同演变的理论体系。
养分循环;土壤微生物;群落结构;多样性;文献计量学;关键词交互网络
土壤养分循环是土壤肥力和作物生长的基础,而土壤微生物驱动了养分的循环过程,明确土壤养分循环的微生物学机制及其调控原理,是提高土壤肥力和养分资源利用效率,实现农业增产增效的理论基础。我国人均耕地资源缺乏、地力水平总体低下,根据《2015中国国土资源公报》,全国中等和低等耕地分别占52.9%和17.7%;耕地基础地力对粮食产量的平均贡献率为45.7%~60.2%[1],比欧美发达国家低20~30个百分点。同时,在追求高产目标下,我国化肥投入水平高,施用方式不尽合理,导致其利用率低下,2015年我国化肥总用量约6000万吨,单位耕地面积化肥用量是世界平均水平的3倍,但粮食作物的化肥利用率仅为35.2%,比欧美低10~30个百分点[2]。我国耕地土壤类型丰富,地力水平差异巨大,耕作轮作和施肥管理方式多样,不同农业主产区根据气候、土壤和生物资源特色,针对土壤培肥和养分增效建立了多样化的物理和化学调控措施;同时不断发展土壤微生物学调控方法,充分挖掘土壤微生物调控养分循环功能的潜力,通过建立土壤养分的协同管理措施,降低使用成本和环境风险,实现粮食安全和环境安全协调发展。然而,在土壤养分循环研究领域,与土壤物理和土壤化学研究相比,我国土壤微生物学研究起步较晚,一直受到微生物可培养性和高分辨率研究方法滞后的限制,在土壤微生物分布格局与影响因子、演替过程与驱动机制、微生物功能调控原理与技术等方面亟需系统深入的研究。因此,梳理我国土壤养分循环和土壤微生物学交互研究的发展历史和脉络,审视与国际相关学科前沿研究的不足之处,有利于促进我国在土壤养分循环的微生物机制和调控方法领域的研究水平。
在分析不同学科领域的研究进展方面,传统的文献综述方法通常基于研究者深厚的科研基础和创新思维,通过梳理和分析某一领域数十篇到百余篇尺度的重要文献,总结已取得的关键成果,提出未来的研究方向。随着土壤微生物学研究深度和广度的不断拓展,文献数量激增,利用传统的文献分析方法已经无法处理数千篇尺度的文献信息,难以精准描述研究内容的动态发展全貌。近年来,以大数据可视化为基础的文献计量学的发展,有助于研究者全面了解研究领域的发展概况,敏锐发现研究发展的新趋势和新动态。其中,引文网络分析工具Citespace基于对目标文献数据计量和分析,利用综合定量分析关键词并形成关键词交互网络,能够以可视化方式直观展现出研究领域在不同发展阶段的热点[3]。本文针对土壤养分循环微生物机制的研究领域,利用Citespace系统梳理了我国在1950~2016年的发展历史和脉络,对比分析了60多年来国内外发展的异同,提出了我国未来的研究方向。
1 文献计量研究方法与数据来源
本文利用 Web of science 和中国知网 (CNKI) 数据库1950~2016年的数据,通过对 “养分循环”、“微生物”和“土壤”主题进行检索,检索到的数据利用Citespace软件进行可视化统计分析。其中养分循环主题词包括碳、氮和磷,微生物主题词包括参与碳、氮、磷转换的微生物数量、微生物种群、功能基因和土壤酶活性,土壤主题词包括不同的土壤类型。
中文检索式为:SU = 碳 + 秸秆还田 + 秸秆腐解 + 有机肥 + 绿肥 + 生物炭 + 土壤有机质 + CO2排放 + CH4排放 + 氮 + 氨氧化 + 硝化 + N2O 排放 + NO +nifH + amoA + hao + nirS + nirK + norB + nosZ + 面源污染 + 磷 + 磷肥 + 有机磷 + 无机磷 + 磷溶解 + 丛枝菌根真菌 + phoD + ALP + phoR + phoB AND SU = 细菌 + 真菌 + 放线菌 + 蚯蚓 + 线虫 + 微生物 + 生物量 + 群落 + 多样性 + 酶活性 + 功能基因 AND SU =农田 + 红壤 + 水稻土 + 耕地 + 黑土 + 潮土 + 褐土 +黄棕壤 + 紫色土 + 黑垆土 + 黄绵土 +土 + 盐碱土 +菜园土 + 果园土壤 + 菜地 + 绿洲土壤。
英文检索式为:TS= (“Carbon ” or “straw return” or “straw decomposition” or “manure” or“mineral fertilizer” or “biochar” or “C cycling”or “C mineralization” or “soil organic matter” or“CO2emission” or “ CH4emission” or“nitrogen” or “N mineralization” or“ammonification” or “nitrification” or“denitrification” or “N2O emission” or “nitrate(NO3)” or “ammonia (NH4)” or “N cycling functional gene” or “AOB” or “AOA” or“nifH” or “amoA” or “hao” or “nirS/K” or“norB” or “nosZ” or “Agricultural Non-Point Source Pollution” or “Phosphorus” or“phosphate” or “ P mineralization” or “P fertilizer” or “P pollution” or “P cycling” or “P fluxes” or “P pool” or “organic P” or “arbuscular mycorrhizal fungi (AMF)” or “phosphorus cycling functional gene” or “phoD” or “ALP” or“phoR” or “phoB” or “nutrient cycling”) AND TS= (“Microorganisms” or “microbial ”* or“enzyme activity”or “Bacteria” or “bacterium”or “fungus” or “Fungi” or “actinomyce”* or“ e a r t h w o r m ” o r “ n e m a t o d e” ) A N D TS=(“farmland” or “arable soil” or “cropland” or“cultivated land” or “field experiment” or “red earth” or “red soil” or “latosol” or “orchard soil” or “vegetable soil” or “cinnamon soil” or“black soil” or “alluvial soil” or “yellow brown earth” or “paddy soil” or “purple soil” or “loess soil” or “lou soil” or “oasis soil” or “saline soil” or “cambisols” or “chernozems” or“ferralsols” or “fulvisol” or “solonetz” or“inceptisols” or “mollisols” or “oxisols” or“ultisols” or “isotopic labelling” or “13C” or“15N” or “growth chamber” or “pot experiment”)。
2 国内土壤养分循环的微生物机制研究
利用中国知网 (核心期刊),检索1950~2016年以“土壤微生物”和“土壤养分循环”为主题的文献,共计3865篇,经人工筛选、去重,删除77篇,保留3788条数据。其中1980年前,年发表论文篇数仅为1~8篇,难以形成网络,故无关键词交互网络图谱。1981~2000年,年发表论文数有所上升,为10~36篇。2000年后,国家加强了对土壤学发展的支持,启动了第一个“973”项目“土壤质量演变规律与持续利用”,2003年的年发表论文数达到108篇,此后呈递增趋势;随后启动实施了“我国农田生态系统重要过程与调控对策研究”、“粮食主产区农田地力提升机理与定向培育对策”和“肥料养分持续高效利用机理与途径”等项目,到2016年,年发表论文数已达383篇。从论文总量上看,在该领域有主要贡献的研究机构包括中国科学院南京土壤研究所 (289篇)、西北农林科技大学(236篇)、南京农业大学 (203篇) 和中国农业大学(121 篇)(图1)。
从1950~2016年研究的热点关键词看,土壤酶活性出现频率最高 (427次),说明利用土壤酶活性研究微生物的肥力和养分转化功能始终是一个重要的研究手段。其次是土壤微生物 (190次) 和土壤有机碳 (173 次),土壤养分 (155 次)、长期施肥 (125 次)和土壤肥力 (111次) 出现的频率也位于前列,说明60年来国内研究以服务农业生产为主,注重地上部与地下部养分的吸收和转化。研究的土壤类型在地带性土壤中,黑土 (140 次) 和红壤 (134 次) 受到广泛关注。对于较为肥沃的黑土侧重于研究如何减缓土壤退化[4–5],而对于贫瘠的红壤侧重于研究如何提高土壤质量[6–7]。在非地带性土壤中,水稻土 (164次)出现频率最高,说明我国作为水稻生产大国,一直重视研究稻田养分循环和微生物功能。此外,对紫色土 (73 次),潮土 (55 次) 和盐碱土 (40 次) 的研究也较多,主要关注了这些土壤的特色问题(图1)。
关键词交互网络图谱可以反映关键词之间的关联程度。从整体的关键词交互网络图谱 (图2,1981~2016) 来看,研究中心热点围绕与长期施肥和土壤养分相关的土壤酶活性和土壤微生物。其中,红壤的研究热点为长期施肥等措施下有机碳和土壤肥力的演变与土壤酶活性,黑土和水稻土的主要研究热点为长期施用有机肥和秸秆还田等措施下的土壤酶活性。为反映不同时期研究热点的变化,从1981~2016年,按5年一个时间片段,分析了不同时期的关键词交互网络图谱,发现国内的研究可以分为起步期 (1981~1990 年)、发展期 (1991~2005 年) 和定型期 (2006~2016年) 3个发展阶段,研究从注重单菌和微生物生物量向注重微生物群落结构与养分循环过程深度耦合的方向发展。
2.1 起步期,研究热点分散,主要研究了红壤、水稻土和紫色土中特定微生物和酶活性的肥力功能
图2显示,在1981~1990年,研究热点较为分散独立,主要包括红壤中有机无机肥配施影响下的酶活性的研究[8],水稻土和紫色土中的与土壤肥力相关的碳氮循环[9]。这一时期的另一个研究特色是关注农业生产过程的特定微生物,如与固氮相关的大豆根瘤菌[10],与提高氮磷利用相关的丛枝菌根真菌[11],与土壤碳循环相关的脲酶活性[12],以及应用于堆肥的双孢蘑菇[13]。此外,也研究了棕色森林土中与碳循环有关的微生物[14]。说明本时期的研究注重农业生产实际需求,特别关注水稻土和红壤施用有机和无机肥对土壤养分和肥力的影响。
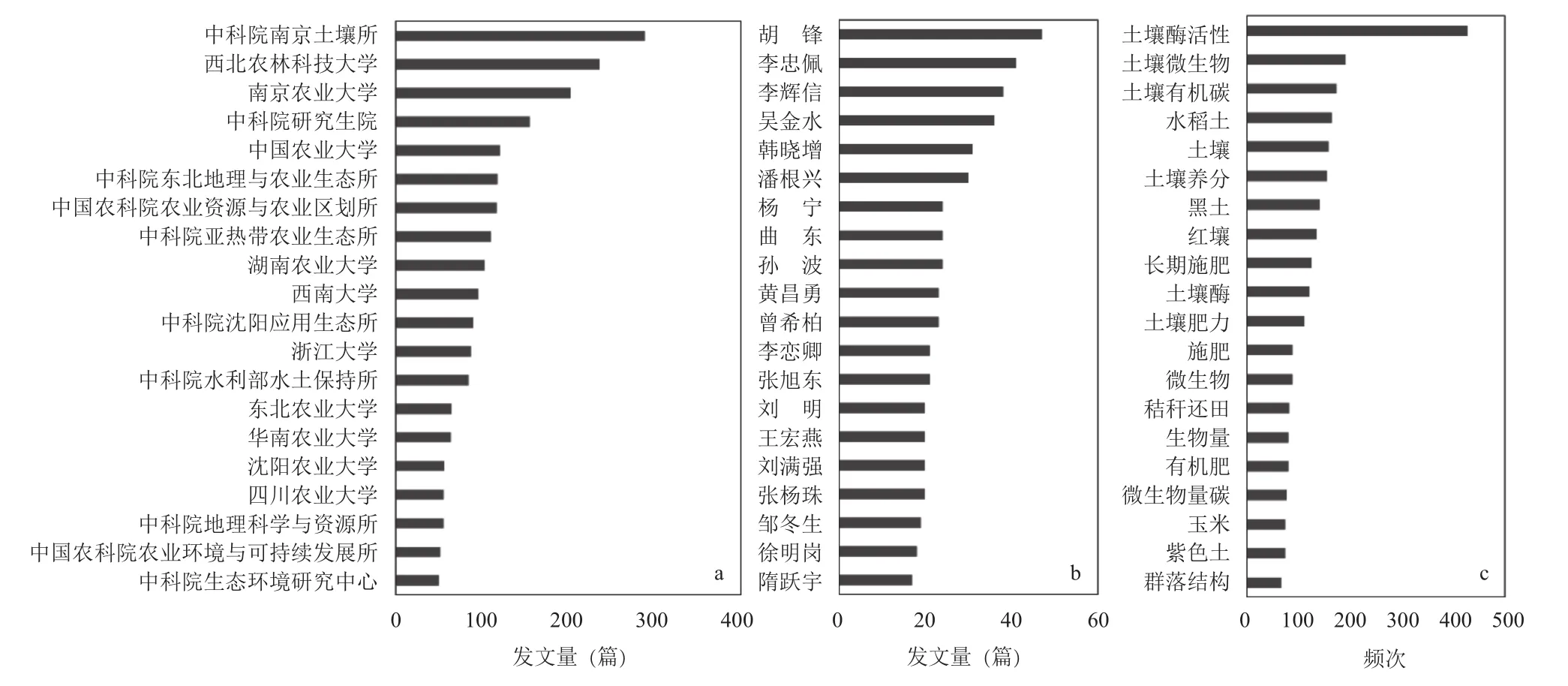
图1 在土壤养分循环微生物机制研究领域主要机构 (a) 和作者 (b) 的发文量以及高频关键词 (c)Fig. 1 The publication numbers of main institutes (a) and authors (b), and the high frequency key words (c) related to microbial mechanisms of soil nutrient cycling
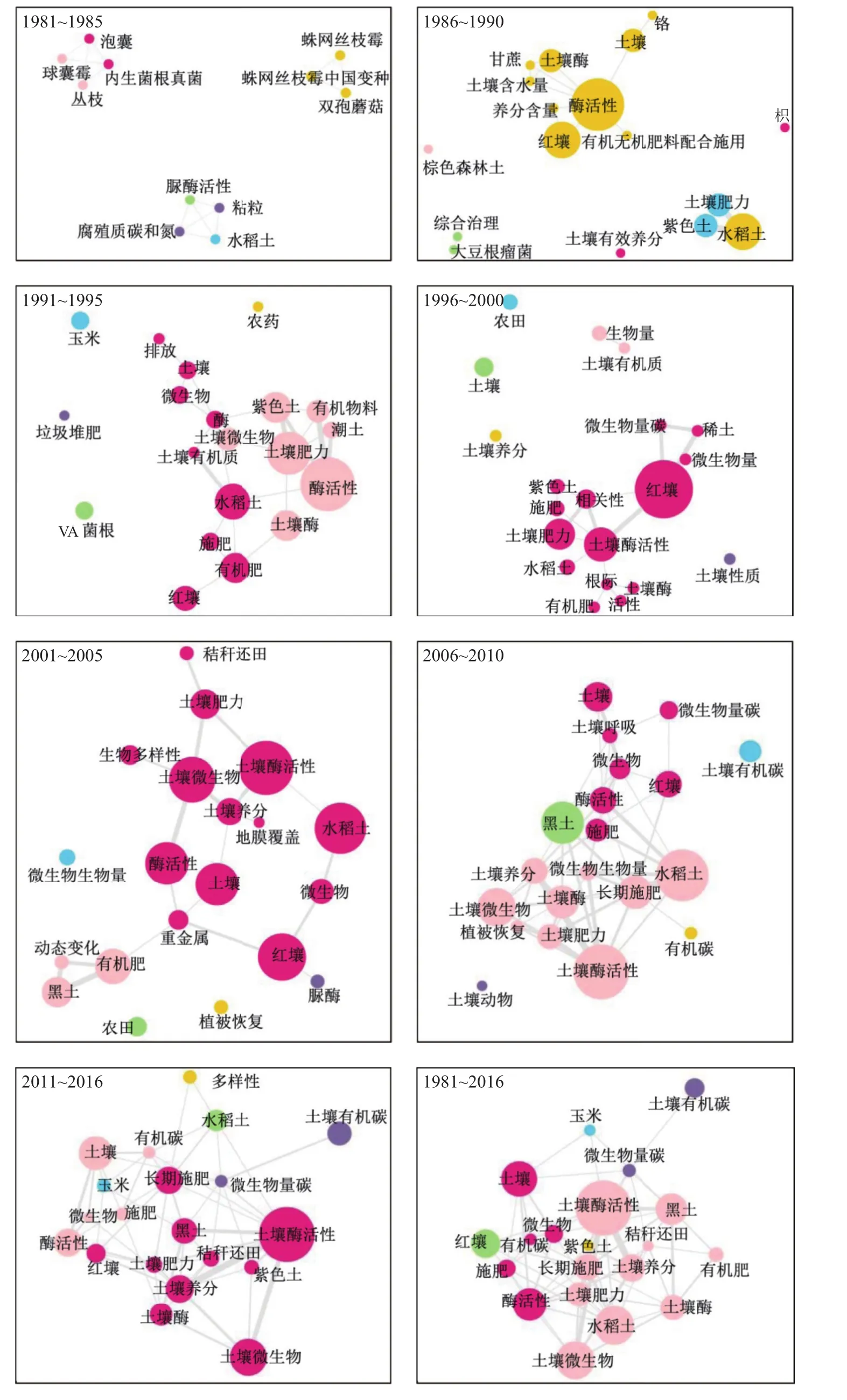
图2 不同年代国内期刊相关研究论文的关键词交互网络Fig. 2 Keyword co-occurring networks of related papers published in Chinese journals during different periods
2.2 发展期,研究热点关联度加强,主要研究了红壤、黑土、水稻土中微生物量和酶活性的肥力和环境功能
1991~2005年,关键词交互网络图谱逐渐呈现网络化,表现出发展态势,但仍然存在分散点 (如VA菌根、有机碳等)。本时期的主要研究热点仍然围绕与土壤肥力和养分相关的酶活性,在土壤类型中水稻土和红壤一直处于研究热点的中心,特别是针对长期施肥下土壤酶活性演变及其调控肥力的作用进行了深入研究[6, 15]。在对土壤酶活性研究的同时,本时期引入了土壤微生物量的概念,但其测定方式通常采用传统的熏蒸浸提方法[16]。在本时期的后段 (2001~2005年) 加强了对治理黑土退化的研究,研究了施用有机肥对土壤酶活性和土壤碳库的影响[17–18]。由于这一时期化肥大量施用导致水体氮磷污染和温室气体排放问题日益突出,国内开始关注N2O排放的研究[19]。此外,也加强了重金属污染对土壤酶活性影响的研究[20](图2)。
2.3 定型期,研究热点间的连接度不断增强,主要研究了土壤养分循环与微生物群落结构耦合的微生物学机制
2006~2016年,不同研究热点间的网络连接度不断增强,孤立的研究热点减少,网络结构趋于稳定。这一时期主要研究热点仍然以土壤酶活性为核心,随着变形梯度凝胶电泳 (DGGE) 和磷脂脂肪酸(PLFA) 等分子生物学研究方法的发展,开始研究微生物群落结构和多样性[21],加强了对微生物生物量和群落结构影响土壤功能的研究[22–23],特别是在土壤氮素转化[24]以及稻田温室气体排放[25]的微生物学机制方面展开了深入研究。同时,本时期加强了对土壤有机碳循环与微生物群落多样性交互作用的研究,揭示了土地利用方式和秸秆还田措施如何影响微生物群落功能多样性进而影响有机碳循环和土壤肥力的机制[22, 26]。此外,本时期出现了对土壤动物的研究,完善了对土壤生物网络功能的研究[27](图2)。
3 国际土壤养分循环的微生物机制研究及其与国内研究的比较
利用 web of science,检索从 1981~2016 年以“土壤微生物”和“土壤养分循环”为主题的SCI论文,共计6023篇,其中中国作者论文数为1494篇。1981~1990年,国际上发表论文仅25篇,难以形成网络,故无关键词交互网络图谱。1991~1995年,发表论文数上升为274篇,此后逐渐增加,至2011~2016年期间,共发表论文3174篇。中国作者发表的SCI论文数也不断增长,但相对滞后,1990年前未检索到发表的论文,1991~2000年仅发表12篇论文,2001~2006年增长到66篇,2011年以后爆发式增长,2011~2016年共发表论文1151篇。
为比较国内学者与国际学者对养分循环微生物学机制研究的发展脉络,从1990~2016年按照5年一个时间片段,分析不同时期的关键词交互网络图谱。总体上看,在不同时期国际学者的研究热点始终围绕土壤氮循环 (nitrogen,nitrification,denitrification) 和碳循环 (carbon,organic matter,decomposition) 与微生物群落的交互作用;其次,磷(phosphorous)、根 (root)、根际 (rhizosphere)、土壤(soil)、植被 (plant) 等关键词也贯穿始终,说明国际学者在该领域研究侧重于农田土壤养分循环及其肥力功能,特别关注土壤“热区”—根际微环境的微生物学驱动机制。与国际学者的研究相比,国内学者的研究起步较晚、发展相对滞后,国内学者的研究热点也关注碳氮养分循环及其与微生物生物量关系的研究,与国际学者的研究热点相似,并日趋一致。总体上,国内学者追赶国际学者研究热点的发展进程可以分为2个阶段,起步追赶期 (1991~2005 年) 和快速追赶期 (2006~2016 年)。
3.1 起步追赶期,国内学者追随国际上对碳氮转化与微生物交互研究的热点,但研究网络成熟程度低,侧重对红壤的研究,缺少对根际的研究
图3显示,在1990~2005年期间,从国际学者研究体系看,关键词交互网络出现早且相对成熟,碳氮循环与微生物群落的交互作用是贯穿始终的研究热点。其中,对氮素 (nitrogen) 的研究一直处于中心连接点地位,1996~2000年出现对硝化和反硝化的研究[28–29],后续深入研究了氮循环中起关键作用的功能基因[30];对有机质 (organic matter) 和碳 (carbon)的研究涉及矿化和分解等方面;对磷的研究相对较弱,但一直持续;此外,1996年以后出现对根际和水稻土的研究,说明这一时期国际学者日益关注根际土壤碳氮磷循环的微生物驱动机制,为提高土壤肥力和氮磷利用率提供理论基础。
与国际学者的研究相比,国内学者在养分循环微生物机制领域的研究起步较晚,发表的论文数量少,关键词的频率较低,不同关键词之间的交互联系较少,关键词交互网络的成熟度较低,直至2001~2005年才形成明显的交互网络。在这一时期国内学者同样关注微生物生物量与有机质、碳、氮养分转化的交互研究,对磷素的研究较少。针对不同的施肥和耕作措施,国内学者研究了如何利用微生物学手段增加养分利用率,如种植不同作物对红壤生物量碳、氮、磷的影响[31],以及菌根真菌促进作物吸收氮磷的机制[32–33]。在土壤类型方面的研究特色是关注红壤,在土壤热区特别是根际研究方面较为薄弱。此外,由于环境污染问题凸显,国内学者还关注土壤重金属污染以及生物降解机制的研究[34]。
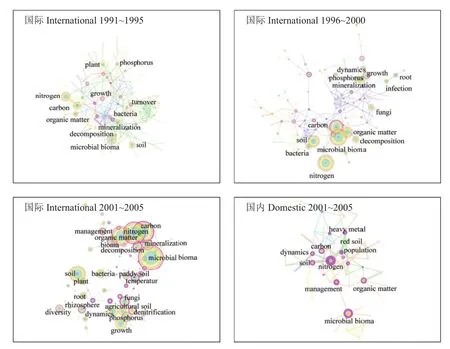
图3 1991~2005 年国际和国内学者发表的相关研究论文的关键词交互网络图谱Fig. 3 Keyword co-occurring networks of related papers published by international and domestic researchers in international journals during 1991 to 2005
3.2 快速追赶期,国内学者从水稻土和根际方面强化了对养分转化微生物机制研究,开展了土壤微生物网络结构和功能的研究,但需要加强土壤生物网络结构和功能的实证研究
图4显示,与前期研究相比,国际学者对土壤有机质、碳、氮周转微生物学机制的研究向纵深发展,关键词交互联系加强,研究网络更加成熟,但对磷素研究较弱的格局仍然持续。这一时期,国际学者的研究特色首先表现为加强了对微生物群落多样性和微生物分子生态网络的研究,特别是对环境变化响应的定量研究。由于分子生物学研究技术快速发展,对微生物群落结构和多样性 (diversity) 的研究日益增加,至2011~2016年关键词diversity的出现频率成为仅次于微生物生物量 (microbial biomass)的关键词,并且位于网络中心的关键节点之一。从近三年 (2014~2016 年) 新出现的关键词来看 (表 1),高通量测序方法开始更新换代,Miseq和Hiseq Illumina取代了454 pyrosequencing,功能基因微阵列(Fuction Gene Arrays) 方法 (如 Geochip) 加强了对特定功能基因的研究,同时微生物系统发育/功能分子生态网络 (phylogenetic/functional molecular ecological networks,pMENs/fMENs) 和结构方程模型 (structural equation modeling,SEM) 等统计分析方法不断发展,土壤微生物学加强了对微生物网络的研究,在微生物网络响应环境条件变化发展方面取得了新的突破[35]。
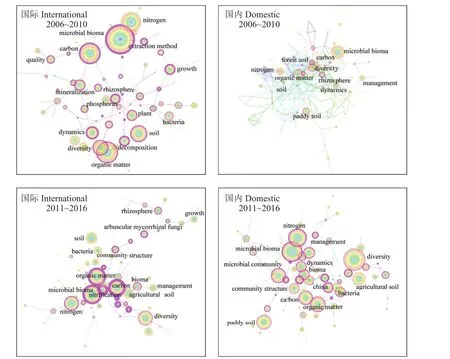
图4 2006~2016 年国际和国内学者发表的相关研究论文的关键词交互网络Fig. 4 Keyword co-occurring networks of related papers published by international and domestic researchers in international journals during 2006 to 2016
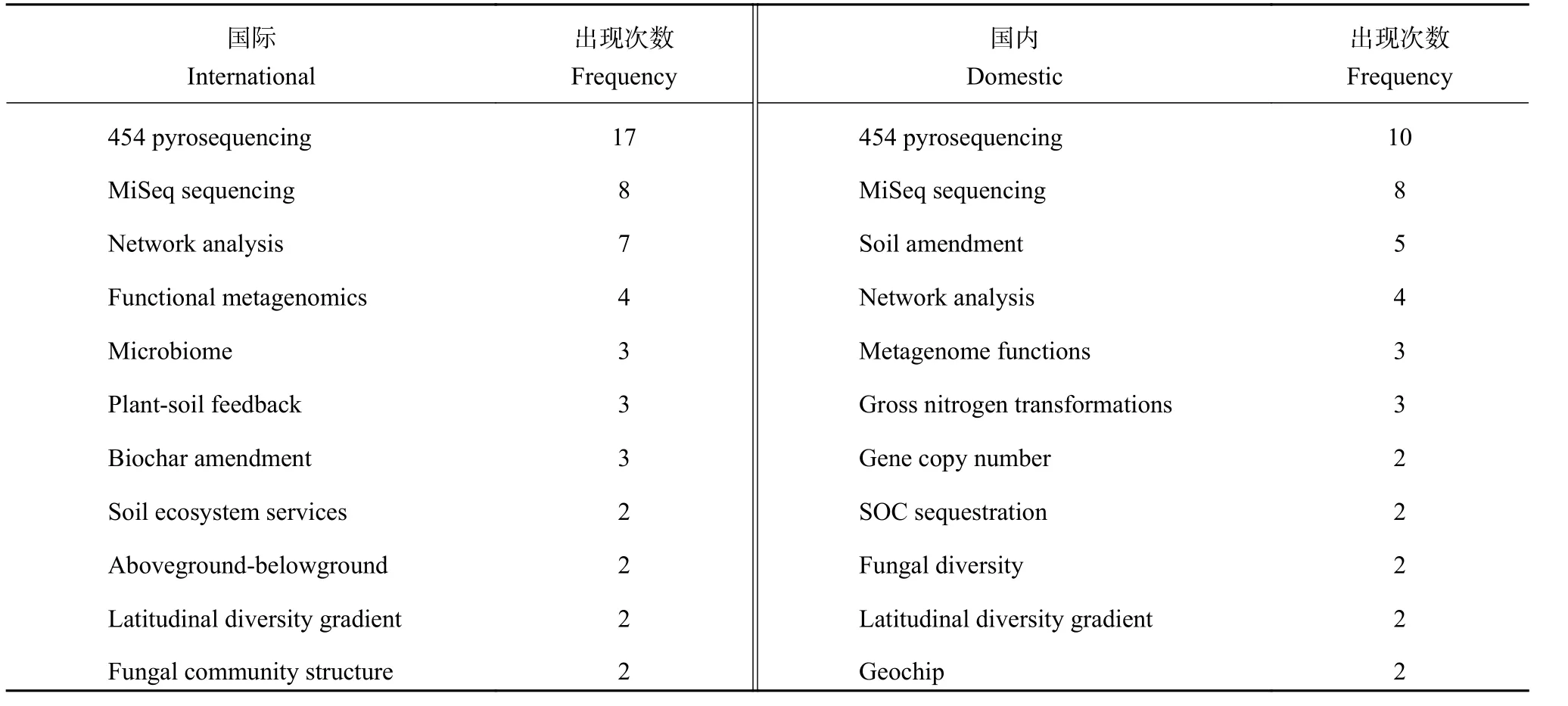
表1 2012~2016 年在国际国内期刊上发表的论文中新出现的关键词及其频率Table 1 The burst frequency of key words in the related papers published in international and Chinese journals during 2012 to 2016
国际学者的第二个研究特色表现为加强了对碳、氮转化的协同研究,以及对土壤–植物互作机制和协同管理的研究。土壤是地球表层系统中最大的碳库,对其研究不断加强,2006~2010年期间,关于碳 (carbon) 以及与碳循环相关的土壤有机质(organic matter)、矿化 (mineralization) 的研究和氮(nitrogen) 一起处于中心连接点的地位。图中共现carbon与nitrogen、management的交互,说明对于固碳机制的认识加深,一方面通过保护性耕作 (conservation tillage)、秸秆还田 (straw returning)、有机肥施用(manure) 等措施协同调控碳、氮转化,达到增加土壤碳库[36]、改良土壤[37]和减少温室气体排放[38]等生态目标;另一方面,深入研究微生物生物量和群落结构对秸秆分解过程的影响,提出利用微生物手段调控秸秆还田的理论依据[39–40]。除了秸秆还田,在此时期兴起对生物炭 (biochar) 的研究,探讨生物炭对土壤固碳、土壤改良以及对土壤微生物的影响[41–42]。此后 (2011~2016年),农田土壤中有机质转化的微生物驱动机制日益成为研究热点,关键词agriculture soil、microbial biomass和 organic matter的交互,说明研究进一步深入和细化。对有机质转化过程的影响因素和土壤稳定有机质形成理论开展了深入研究,揭示了秸秆还田[43, 35]和生物炭[44–45]影响微生物群落演变和土壤养分循环的机制,提出了增加土壤有机质含量和碳库库容的新理论[46–47]。从近三年新出现的关键词来看 (表1),植物与土壤的互作和地上与地下的协同管理成为未来农田养分管理的研究热点。
与国际学者的研究相比,这一时期国内学者从水稻土和根际方面强化了对土壤有机质以及碳氮转化的微生物机制研究。国内学者研究的重点领域与国际趋同,对微生物群落结构和多样性与有机质转化以及氮循环的交互作用研究有所加强[48–49],但仍需加强对微生物群落结构及其多样性与碳氮循环过程交互作用机制的研究。目前国内外学者在根际–微生物互作促进氮磷协同利用方面已有初步研究[50–51],但仍需进一步挖掘微生物促进碳氮磷养分的协同转化和利用机制。这一时期,微生物在关键地球化学过程中的作用成为研究重点,特别是对微生物群落结构与有机质和水稻土 (paddy soil) 的交互研究[52–53];通过长期研究揭示了碳氮养分循环和温室气体排放的协同过程以及微生物群落的作用[38, 54]。国内学者的研究更偏重于农田土壤,特别是水稻土的有机质周转;同时,日益关注根际微生物的群落结构及其对根际养分转化的影响,强调通过微生物调控促进作物养分吸收和增产[55]。虽然国际上对根际和团聚体等“热区”中磷转化微生物机制的研究热度不及碳氮循环,但仍十分关注豆科和禾本科作物间作系统中磷素高效利用的根际微生物学机制[56]。国内学者在探究培肥措施对根际磷转化微生物长期影响的基础上,深入研究了团聚体中土壤线虫捕食解磷菌对其群落组成和活性的影响机制,发现不同粒径团聚体中线虫–解磷菌生态网络的关键控制物种不同[57]。
另外,国内学者追赶国际学者的研究热点,在微生物群落多样性和网络结构及其功能方面迅猛发展。一方面,加强了对不同生境下微生物群落多样性分布格局及其影响因素的研究[58–59],对象也从前期注重细菌扩展到对真菌群落结构多样性的研究 (表1),特别是利用跨气候带的土壤置换试验深入研究了气候变化对土壤养分转化微生物群落结构演替的影响机制[60]。另一方面,近三年通过应用高通量测序和微生物网络分析方法,深入研究了环境条件变化对微生物群落结构的影响[61],通过分析土壤微生物网络结构和关键物种的变化,揭示环境因子和种植管理措施对土壤养分转化的影响机制。这一时期,国内学者也十分重视土壤食物网内不同生物之间的交互作用及其对养分转化的影响,研究了不同管理措施下土壤原生动物 (线虫) 对细菌的捕食作用[62];同时深入研究了不同团聚体中线虫捕食对微生物碳氮转化功能的影响,发现大团聚体中线虫选择性捕食氨氧化细菌促进了土壤硝化作用[63]。然而,国内的研究仍需针对不同环境条件和种植管理措施,实证土壤生物网络结构中关键物种对养分转化功能的调控作用,从而提出土壤生物功能的调控措施。
4 结论
本文针对土壤养分循环的微生物机制研究领域,利用文献计量学及大数据可视化方法定量分析了60多年来 (1950~2016年) 的文献数据,分析了不同时期国内外研究热点关键词交互网络的变化历程,发现国际上自1990年后对养分循环微生物机制的研究呈现迅速发展趋势,伴随2001年以来分子生物学技术的快速升级,其研究向纵深和广度推进,但对土壤碳、氮周转的微生物学机制研究一直处于中心地位,并且重视对碳氮协同转化以及根际土壤–植物互作机制的研究。一方面,国内的研究跟随国际研究热点的变化趋势,同样在2001年以来发展迅猛,研究发展趋势表现为:从孤立地研究特定微生物的功能,发展为综合地研究微生物分子生态网络对土壤养分循环的驱动作用;从微生物土壤肥力功能的研究,发展为研究微生物对全球变化 (温室气体排放) 以及重金属污染等环境功能。另一方面,国内研究热点网络形成相对滞后,但其结构日趋成熟,表现出与国际研究趋同趋势,但偏重研究水稻土等重要农业土壤,近期十分重视研究根际微生物结构以及土壤团聚体中食物网交互作用对养分转化功能的调控作用,研究偏重于提升不同区域农田养分资源的综合利用效率问题。未来应针对不同气候、作物、土壤条件以及不同种植管理措施,深入研究土壤微生物群落结构的演变特征,以及生物之间交互作用对其养分转化功能的影响机制,修正控制土壤微生物群落分布和演替的理论体系 (如代谢理论和中性理论等),实证土壤生物网络关键物种对养分转化的控制作用,提出调控土壤生物网络结构和功能的措施。
[1]汤勇华, 黄耀. 中国大陆主要粮食作物地力贡献率和基础产量的空间分布特征[J]. 农业环境科学学报, 2009, 28(5): 1070–1078.Tang Y H, Huang Y. Spatial distribution characteristics of the percentage of soil fertility contribution and its associated basic crop yield in mainland China[J]. Journal of Agro-Environment Science,2009, 28(5): 1070–1078.
[2]郝锐敏. 农业部首度发布化肥农药利用率数据[N]. 农资导报,2015-12-11.Hao R M. Fertilizer and pesticide use efficiency data was first issued by Ministry of Agriculture [N]. AgriGoods Herald, 2015-12-11.
[3]宋长青. 土壤科学三十年: 从经典到前沿[M]. 北京: 商务印书馆,2016.Song C Q. Development of soil science in the past 30 years: From classic to frontier [M]. Beijing: The Commercial Press, 2016.
[4]孙华, 张桃林, 熊德祥. 退化砂姜黑土的有机磷特性及其活化研究[J]. 山东农业大学学报 (自然科学版), 2003, 34(2): 214–216.Sun H, Zhang T L, Xiong D X. Studies on organic phosphorus and its activation of degraded Shajiang black soils[J]. Journal of Shandong Agricultural University (Natural Science), 2003, 34(2): 214–216.
[5]李东坡, 武志杰, 陈利军, 等. 长期培肥黑土脲酶活性动态变化及其影响因素[J]. 应用生态学报, 2003, 14(12): 2208–2212.Li D P, Wu Z J, Chen L J, et al. Dynamics of urease activity in a long-term fertilized black soil and its affecting factors[J]. Chinese Journal of Applied Ecology, 2003, 14(12): 2208–2212.
[6]孙波, 张桃林, 赵其国. 我国中亚热带缓丘区红粘土红壤肥力的演化Ⅱ.化学和生物学肥力的演化[J]. 土壤学报, 1999, 36(2):203–217.Sun B, Zhang T L, Zhao Q G. Fertility evolution of red soil derived from quaternary red clay in low-hilly region in middle subtropics Ⅱ.Evolution of soil chemical and biological fertilities[J]. Acta Pedologica Sinica, 1999, 36(2): 203–217.
[7]李忠佩, 唐永良, 石华, 张桃林. 不同轮作措施下瘠薄红壤中碳氮积累特征[J]. 中国农业科学, 2002, 35(10): 1236–1242.Li Z P, Tang Y L, Shi H, Zhang T L. Characteristics of C and N accumulation in infertile red soil under different rotation systems[J].Scientia Agricultura Sinica, 2002, 35(10): 1236–1242.
[8]关连珠, 张伯泉, 颜丽, 等. 有机肥料配施化肥对土壤有机质组分及生物活性影响的研究[J]. 土壤通报, 1990, 21(4): 180–184.Guan L Z, Zhang B Q, Yan L, et al. Research on effects of biological activity and organic matter for fertilizer combined with organic matter in soil[J]. Chinese Journal of Soil Science, 1990, 21(4):180–184.
[9]廖先苓, 徐银华, 朱兆良. 淹水种稻条件下化肥氮的硝化–反硝化损失的初步研究[J]. 土壤学报, 1982, 19(3): 257–263.Liao X L, Xu Y H, Zhu Z L. Investigation on nitrificationdenitrification loss of fertilizer nitrogen in submerged paddy soil[J].Acta Pedologica Sinica, 1982, 19(3): 257–263.
[10]郑林用, 黄怀琼, 刘世全. 酸性紫色土中钼对花生–根瘤菌共生固氮的影响[J]. 四川农业大学学报, 1990, 8(2): 129–135.Zheng L Y, Huang H Q, Liu S Q. Effect of molybdenum on symbiotic nitrogen-fixation of rhizobium-peanut in acid purple soils[J]. Journal of Sichuan Agricultural University, 1990, 8(2):129–135.
[11]唐振尧, 张清华, 何首林. 菌根真菌在红壤中对柑桔吸收磷肥的研究[J]. 真菌学报, 1984, 3(3): 170–177.Tang Z Y, Zhang Q H, He S L. The effect of mycorrhizal fungus on phospohate uptake by Citrus in red earth[J]. Acta Mycologica Sinica,1984, 3(3): 170–177.
[12]杨明均, 江育璋. 半旱式耕作下紫色水稻土的脲酶活性与土壤肥力[J]. 西南农业大学学报, 1987, (增刊 2): 86–89.Yang M J, Jiang Y Z. A study on relation of urease activity with fertility in the purple soil with rice ridge culture [J]. Journal of Southwest Agricultural University, 1987, (Suppl. 2): 86–89.
[13]陈吉棣, 刘建国, 陈松生, 等. 寄生双孢蘑菇的蛛网丝枝霉中国变种[J]. 真菌学报, 1984, 3(2): 96–101.Chen J L, Liu J G, Chen S S, et al. Studies on Aphanocladium Aranearum (petch) Gams var. Sinense var. nov. — A fungus parasitic to Agaricus bisporus (Lange) Sing. [J]. Acta Mycologica Sinica,1984, 3(2): 96–101.
[14]杨冬生, 王荣康, 万富寅, 乔光辉. 道孚棕色森林土火烧后主要酶系活性及理化性质的演变[J]. 四川林业科技, 1990, 11(2): 1–7.Yang D S, Wang R K, Wang F Y, Qiao G H. Effect of fire on enzyme activity and soil physical and chemical properties in brown forest soil in Daofu[J]. Journal of Sichuan Forestry Science and Technology,1990, 11(2): 1–7.
[15]孙瑞莲, 赵秉强, 朱鲁生, 等. 长期定位施肥对土壤酶活性的影响及其调控土壤肥力的作用[J]. 植物营养与肥料学报, 2003, 9(4):406–410.Sun R L, Zhao B Q, Zhu L S, et al. Effect of long-term fertilization on soil enzyme activities and its role in adjusting-controlling soil fertility[J]. Plant Nutrition and Fertilizer Science, 2003, 9(4):406–410.
[16]陈国潮. 土壤微生物量测定方法现状及其在红壤上的应用[J]. 土壤通报, 1999, 30(6): 284–287.Chen G C. Situation of soil microbial biomass detection methods and its application in red soils[J]. Chinese Journal of Soil Science, 1999,30(6): 284–287.
[17]李东坡, 武志杰, 陈利军, 等. 长期培肥黑土微生物量碳动态变化及影响因素[J]. 应用生态学报, 2004, 15(8): 1334–1338.Li D P, Wu Z J, Chen L J, et al. Dynamics of microbial biomass C in a black soil under long-term fertilization and related affecting factors[J]. Chinese Journal of Applied Ecology, 2004, 15(8):1334–1338.
[18]李东坡, 武志杰, 陈利军, 等. 长期定位培肥黑土土壤蔗糖酶活性动态变化及其影响因素[J]. 中国生态农业学报, 2005, 13(2): 102–105.Li D P, Wu Z J, Chen L J, et al. Dynamics of invertase activity of black soil treated by a long-term located fertilization and its influence[J]. Chinese Journal of Eco-Agriculture, 2005, 13(2):102–105.
[19]王智平, 曾江海, 张玉铭. 农田土壤 N2O 排放的影响因素[J]. 农业环境保护, 1994, 13(1): 40–42.Wang Z P, Zeng J H, Zhang Y M. Factors impact flux of N2O emission from agricultural soils[J]. Agro-environmental Protection,1994, 13(1): 40–42.
[20]杨志新, 刘树庆. 重金属Cd、Zn、Pb复合污染对土壤酶活性的影响[J]. 环境科学学报, 2001, 21(1): 60–63.Yang Z X, Liu S Q. Effect of compound pollution of heavy metals on soil enzymes’ activities[J]. Acta Scientiae Circumstantiae, 2001,21(1): 60–63.
[21]陈法霖, 张凯, 郑华, 等. PCR-DGGE技术解析针叶和阔叶凋落物混合分解对土壤微生物群落结构的影响[J]. 应用与环境生物学报,2011, 17(2): 145–151.Chen F L, Zhang K, Zheng H, et al. Analyzing the effect of mixed decomposition of conifer and broadleaf litters on soil microbial communities by using PCR-DGGE[J]. Chinese Journal of Applied & Environmental Biology, 2011, 17(2): 145–151.
[22]刘定辉, 舒丽, 陈强, 等. 秸秆还田少免耕对冲积土微生物多样性及微生物碳氮的影响[J]. 应用与环境生物学报, 2011, 17(2): 158–162.Liu D H, Shu L, Chen Q, et al. Effects of straw mulching and littleor zero-tillage on microbial diversity and biomass C and N of alluvial soil in Chengdu Plain, China[J]. Chinese Journal of Applied & Environmental Biology, 2011, 17(2): 158–162.
[23]李鑫, 张会慧, 岳冰冰, 等. 桑树–大豆间作对盐碱土碳代谢微生物多样性的影响[J]. 应用生态学报, 2012, 23(7): 1825–1831.Li X, Zhang H H, Yue B B, et al. Effects of mulberry–soybean intercropping on carbon-metabolic microbial diversity in salinealkaline soil[J]. Chinese Journal of Applied Ecology, 2012, 23(7):1825–1831.
[24]刘若萱, 张丽梅, 白刃, 贺纪正. 模拟条件下土壤硝化作用及硝化微生物对不同水分梯度的响应[J]. 土壤学报, 2015, 52(2): 415–422.Liu R X, Zhang L M, Bai R, He J Z. Response of nitrification and nitrifier to change in soil moisture content under simulated conditions[J]. Acta Pedologica Sinica, 2015, 52(2): 415–422.
[25]张坚超, 徐镱钦, 陆雅海. 陆地生态系统甲烷产生和氧化过程的微生物机理[J]. 生态学报, 2015, 35(20): 6592–6603.Zhang J C, Xu Y Q, Lu Y H. Microbial mechanisms of methane production and oxidation in terrestrial ecosystems[J]. Acta Ecologica Sinica, 2015, 35(20): 6592–6603.
[26]李忠佩, 吴晓晨, 陈碧云. 不同利用方式下土壤有机碳转化及微生物群落功能多样性变化[J]. 中国农业科学, 2007, 40(8): 1712–1721.Li Z P, Wu X C, Chen B Y. Changes in transformation of soil organic carbon and functional diversity of soil microbial community under different land use patterns[J]. Scientia Agricultura Sinica, 2007,40(8): 1712–1721.
[27]朱强根, 朱安宁, 张佳宝, 等. 保护性耕作下土壤动物群落及其与土壤肥力的关系[J]. 农业工程学报, 2010, 26(2): 70–76.Zhu Q G, Zhu A N, Zhang J B, et al. Relation of agricultural soil fauna and soil fertility under conservation tillage systems[J].Transactions of the CSAE, 2010, 26(2): 70–76.
[28]Hyvonen R, Agren G I, Andren O. Modelling long-term: Carbon and nitrogen dynamics in an arable soil receiving organic matter[J].Ecological Applications, 1996, 6(4): 1345–1354.
[29]Simek M, Kalcik J. Carbon and nitrate utilization in soils: the effect of long-term fertilization on potential denitrification[J]. Geoderma,1998, 83(3–4): 269–280.
[30]Throback I N, Enwall K, Jarvis A, Hallin S. Reassessing PCR primers targeting nirS, nirK and nosZ genes for community surveys of denitrifying bacteria with DGGE[J]. FEMS Microbiology Ecology,2004, 49(3): 401–417.
[31]Wang F E, Chen Y X, Tian G M, et al. Microbial biomass carbon,nitrogen and phosphorus in the soil profiles of different vegetation covers established for soil rehabilitation in a red soil region of southeastern China[J]. Nutrient Cycling in Agroecosystems, 2004,68(3): 181–189.
[32]Feng G, Zhang F S, Li X L, Christie P. Uptake of nitrogen from indigenous soil pool by cotton plant inoculated with arbuscular mycorrhizal fungi[J]. Communications in Soil Science and Plant Analysis, 2002, 33(19–22): 3825–3836.
[33]Feng G, Song Y C, Li X L, Christie P. Contribution of arbuscular mycorrhizal fungi to utilization of organic sources of phosphorus by red clover in a calcareous soil[J]. Applied Soil Ecology, 2003, 22(2):139–148.
[34]Liao M, Chen C L, Huang C Y. Effect of heavy metals on soil microbial activity and diversity in a reclaimed mining wasteland of red soil area[J]. Journal of Environmental Sciences, 2005, 17(5):832–837.
[35]Banerjee S, Kirkby C A, Schmutter D, et al. Network analysis reveals functional redundancy and keystone taxa amongst bacterial and fungal communities during organic matter decomposition in an arable soil[J]. Soil Biology & Biochemistry, 2016, 97: 188–198.
[36]Embacher A, Zsolnay A, Gattinger A, Munch J C. The dynamics of water extractable organic matter (WEOM) in common arable topsoils: II. Influence of mineral and combined mineral and manure fertilization in a Haplic Chernozem[J]. Geoderma, 2008, 148(1):63–69.
[37]Cherif H, Ayari F, Ouzari H, et al. Effects of municipal solid waste compost, farmyard manure and chemical fertilizers on wheat growth,soil composition and soil bacterial characteristics under Tunisian arid climate[J]. European Journal of Soil Biology, 2009, 45(2): 138–145.
[38]Ding W X, Meng L, Yin Y F, et al. CO2emission in an intensively cultivated loam as affected by long-term application of organic manure and nitrogen fertilizer[J]. Soil Biology & Biochemistry, 2007,39: 669–679.
[39]Murase J, Matsui Y, Katoh M, et al. Incorporation of 13 C-labeled rice-straw-derived carbon into microbial communities in submerged rice field soil and percolating water[J]. Soil Biology & Biochemistry,2006, 38: 3483–3491.
[40]Moore-Kucera J, Dick R P. Application of C-13-labeled litter and root materials for in situ decomposition studies using phospholipid fatty acids[J]. Soil Biology & Biochemistry, 2008, 40: 2485–2493.
[41]Warnock D D, Mummey D L, McBride B, et al. Influences of nonherbaceous biochar on arbuscular mycorrhizal fungal abundances in roots and soils: Results from growth-chamber and field experiments[J]. Applied Soil Ecology, 2010, 46(3): 450–456.
[42]Yang X B, Ying G G, Peng P A, et al. Influence of biochars on plant uptake and dissipation of two pesticides in an agricultural soil[J].Journal of Agricultural and Food Chemistry, 2010, 58(13):7915–7921.
[43]Sun B, Wang X Y, Wang F, et al. Assessing the relative effects of geographic location and soil type on microbial communities associated with straw decomposition[J]. Applied and Environmental Microbiology, 2013, 79(11): 3327–3335.
[44]Zhang A F, Liu Y M, Pan G X, et al. Effect of biochar amendment on maize yield and greenhouse gas emissions from a soil organic carbon poor calcareous loamy soil from Central China Plain[J]. Plant and Soil, 2012, 351(1–2): 263–275.
[45]Quilliam R S, DeLuca T H, Jones D L. Biochar application reduces nodulation but increases nitrogenase activity in clover[J]. Plant and Soil, 2013, 366(1–2): 83–92.
[46]Schmidt M W I, Torn M S, Abiven S, et al. Persistence of soil organic matter as an ecosystem property[J]. Nature, 2011, 478(7367):49–56.
[47]Lehmann J, Kleber M. The contentious nature of soil organic matter[J]. Nature, 2015, 528(7580): 60–68.
[48]Zhang Y G, Zhang X Q, Liu X D, et al. Microarray-based analysis of changes in diversity of microbial genes involved in organic carbon decomposition following land use/cover changes[J]. FEMS Microbiology Letters, 2007, 266(2): 144–151.
[49]Xie W J, Zhou J M, Wang H Y, et al. Short-term effects of copper,cadmium and cypermethrin on dehydrogenase activity and microbial functional diversity in soils after long-term mineral or organic fertilization[J]. Agriculture Ecosystems & Environment, 2009,129(4): 450–456.
[50]Pii Y, Mimmo T, Tomasi N, et al. Microbial interactions in the rhizosphere: beneficial influences of plant growth-promoting rhizobacteria on nutrient acquisition process. A review[J]. Biology and Fertility of Soils, 2015, 51(4): 403–415.
[51]Li B, Li Y Y, Wu H M, et al. Root exudates drive interspecific facilitation by enhancing nodulation and N2fixation[J]. Proceedings of the National Academy of Sciences of the United States of America, 2016, 113(23): 6496–6501.
[52]Yang C M, Yang L Z, Yan T M. Chemical and microbiological parameters of paddy soil quality as affected by different nutrient and water regimes[J]. Pedosphere, 2005, 15(3): 369–378.
[53]Zhang P J, Zheng J F, Pan G X, et al. Changes in microbial community structure and function within particle size fractions of a paddy soil under different long-term fertilization treatments from the Tai Lake region, China[J]. Colloids and Surfaces B: Biointerfaces,2007, 58(2): 264–270.
[54]Zhong W H, Cai Z C. Long-term effects of inorganic fertilizers on microbial biomass and community functional diversity in a paddy soil derived from quaternary red clay[J]. Applied Soil Ecology, 2007,36(2–3): 84–91.
[55]Nie M, Zhang X D, Wang J Q, et al. Rhizosphere effects on soil bacterial abundance and diversity in the Yellow River Deltaic ecosystem as influenced by petroleum contamination and soil salinization[J]. Soil Biology & Biochemistry, 2009, 41: 2535–2542.
[56]Tang X Y, Placella S A, Dayde F, et al. Phosphorus availability and microbial community in the rhizosphere of intercropped cereal and legume along a P-fertilizer gradient[J]. Plant and Soil, 2016, 407:119–134.
[57]Jiang Y, Liu M, Zhang J B, et al. Nematode grazing promotes bacterial community dynamics in soil at the aggregate level [J]. The ISME Journal, 2017. doi:10.1038/ismej. 2017.120.
[58]Jiang H, Deng Q, Zhou G, et al. Responses of soil respiration and its temperature/moisture sensitivity to precipitation in three subtropical forests in southern China[J]. Biogeosciences, 2013, 10(6):3963–3982.
[59]Liu J J, Sui Y Y, Yu Z H, et al. High throughput sequencing analysis of biogeographical distribution of bacterial communities in the black soils of northeast China[J]. Soil Biology & Biochemistry, 2014, 70:113–122.
[60]Liang Y, Jiang Y, Wang F, et al. Long-term soil transplant simulating climate change with latitude significantly alters microbial temporal turnover[J]. The ISME Journal, 2015, 9: 2561–2572.
[61]Zhou J, Deng Y, Luo F, et al. Phylogenetic molecular ecological network of soil microbial communities in response to elevated CO2[J]. MBio, 2011, 2(4): e00122–11.
[62]Pan F J, Xu Y L, McLaughlin N B, et al. Response of soil nematode community structure and diversity to long-term land use in the black soil region in China[J]. Ecological Research, 2012, 27(4): 701–714.
[63]Jiang Y J, Sun B, Li H X, et al. Aggregate-related changes in network patterns of nematodes and ammonia oxidizers in an acidic soil[J].Soil Biology & Biochemistry, 2015, 88: 101–109.
The historical venation in research on microbial mechanisms of soil nutrient cycling in the past 60 years—Based on bibliometric analysis and big data visualization
SUN Bo1, WANG Xiao-yue1#, LÜ Xing-hua2*
( 1 State Key Laboratory of Soil and Sustainable Agriculture, Institute of Soil Science, Chinese Academy of Sciences, Nanjing 210008, China; 2 Wuhan Branch of the National Science Library, Chinese Academy of Sciences, Wuhan 430071, China )
【Objectives】 Nutrient cycling plays an essential role in soil fertility and plant growth, which is mainly driven by soil microorganisms. Exploiting the potentiality of microbe in regulating nutrient turnover in soil–plant systems has become an important tendency to increase nutrient use efficiency. Reviewing the historical advance in researches on nutrient cycling and soil microbial community, and figuring out ouradvantages and disadvantages in these fields on the world will be great helpful to find the key research fields and further promote the domestic research level. 【Methods】 In this study, bibliometrics and big data visualisation were used to quantitatively analyze the historical advances achieved in nutrient cycling and microbial community researches in the past 60 years. The similarities and differences in different periods were compared between domestic and international researches. 【Main advances】 The domestic studies could be classified into starting (1981–1990), developing (1991–2005) and maturing stages (2006–2016). At starting stage, the“hot spots” were scattered and researchers mainly focused on the fertility related function of specific microorganisms and enzyme activities in red soil, paddy soil and purple soil. During the developing stage,researchers were mainly focused on microbial biomass and enzyme activities that related to soil fertility and environmental functions, and the connecting among the research works was strengthened. After entering the maturing stage, the connections among research hot spots were more developed, and the interaction of microbial community structure and soil nutrient cycling became the dominant hot spots. Since the 1990s, the domestic researchers have followed the cutting-edge international research. This process could be categorized into initial (1990–2005) and rapid catch-up stages (2006–2016). During initial catch-up stage, domestic scholars followed the international trends and studied the interaction between microbial community and carbon and nitrogen cycling. However, the domestic networks were still immature, lacking of rhizosphere studies and merely concentrated on red soil. When coming to rapid catch-up stage, domestic scholars strengthened the studies on microbial mechanisms of nutrient dynamics in paddy soil and rhizosphere. In addition, the rising of research on soil microbial network structure and function conducted by domestic academia appeared at the same time as that by the international academia. However, the new findings on soil microbial network structure and functions need more solid proofs. Over all, domestic studies developed dramatically fast, although relatively later than international ones. Within the past 20 years, more and more researches have transferred from simply enzyme activities to the interaction of microbial co-occurring network and soil function. The domestic network are maturing, and showing the convergent tendency with international network. However, the studies are relatively weak on the driving mechanism of microbial community evolution applied to nutrient cycling.【Conclusions and Prospectives】 In the future studies, efforts should be put on developing appropriate regional practices that are suitable for microbial regulation to improve nutrient use efficiency, and on developing the fundamental theories of the synergetic changes in microbial community structure and function as well.
nutrient cycling; soil microbes; community structure; diversity; bibliometric analysis;keyword co-occurring network
2017–07–29 接受日期:2017–09–07
国家自然科学基金重点项目(41530856);国家重点研发计划项目(2016YFD0200300);国家绿肥产业技术体系(CARS-22-G-14)资助。
孙波(1968—),男,江苏南京人,博士,研究员,主要从事农田土壤养分转化微生物机制与调控技术研究。
Tel:025-86881282;E-mail:bsun@issas.ac.cn。#王晓玥与第一作者同等贡献。 * 通信作者 E-mail:lxh@mail.whlib.ac.cn
