Neural computational modeling reveals a major role of corticospinal gating of central oscillations in the generation of essential tremor
2018-01-05HongenQuChuanxinNiuSiLiManzhaoHaoZixiangHuQingXieNingLan
Hong-en Qu, Chuanxin M. Niu, Si Li Man-zhao Hao Zi-xiang Hu Qing Xie, Ning Lan
1 Laboratory of Neurorehabilitation Engineering, School of Biomedical Engineering, Shanghai Jiao Tong University, Shanghai, China
2 Department of Rehabilitation Medicine, Ruijin Hospital, School of Medicine, Shanghai Jiao Tong University, Shanghai, China
3 Division of Biokinesiology and Physical Therapy, Ostrow School of Dentistry, University of Southern California, Los Angeles, CA, USA
Neural computational modeling reveals a major role of corticospinal gating of central oscillations in the generation of essential tremor
Hong-en Qu1,#, Chuanxin M. Niu1,2,#, Si Li1, Man-zhao Hao1, Zi-xiang Hu1, Qing Xie2,*, Ning Lan1,3,*
1 Laboratory of Neurorehabilitation Engineering, School of Biomedical Engineering, Shanghai Jiao Tong University, Shanghai, China
2 Department of Rehabilitation Medicine, Ruijin Hospital, School of Medicine, Shanghai Jiao Tong University, Shanghai, China
3 Division of Biokinesiology and Physical Therapy, Ostrow School of Dentistry, University of Southern California, Los Angeles, CA, USA
Essential tremor, also referred to as familial tremor, is an autosomal dominant genetic disease and the most common movement disorder.It typically involves a postural and motor tremor of the hands, head or other part of the body. Essential tremor is driven by a central oscillation signal in the brain. However, the corticospinal mechanisms involved in the generation of essential tremor are unclear. Therefore, in this study, we used a neural computational model that includes both monosynaptic and multisynaptic corticospinal pathways interacting with a propriospinal neuronal network. A virtual arm model is driven by the central oscillation signal to simulate tremor activity behavior.Cortical descending commands are classi fied as alpha or gamma through monosynaptic or multisynaptic corticospinal pathways, which converge respectively on alpha or gamma motoneurons in the spinal cord. Several scenarios are evaluated based on the central oscillation signal passing down to the spinal motoneuronsviaeach descending pathway. The simulated behaviors are compared with clinical essential tremor characteristics to identify the corticospinal pathways responsible for transmitting the central oscillation signal. A propriospinal neuron with strong cortical inhibition performs a gating function in the generation of essential tremor. Our results indicate that the propriospinal neuronal network is essential for relaying the central oscillation signal and the production of essential tremor.
nerve regeneration; neurodegeneration; essential tremor; propriospinal neurons; motoneuron; re flex; modeling; motor control;oscillation; neurological disorder; movement disorder; computational neuroscience; neural regeneration
Introduction
Essential tremor, one of the most common neurological disorders (Louis, 2005; Zhao et al., 2011; Hallett, 2014), is often characterized as postural (Martinelli et al., 1987) and/or kinetic tremors accompanying a motor action. Both the postural and kinetic types of essential tremor typically occur in the forearm and hand with frequencies ranging from 4 to 12 Hz (Deuschl et al., 1998; Brennan et al., 2002). A strong correlation was found between the frequencies on electroencephalogram and limb electromyogram in essential tremor patients (Halliday et al., 2000; Hellwig et al., 2001, 2003; Raethjen et al., 2007; Hellriegel et al., 2012), providing insight into the pathological origin of the disorder. Essential tremor is considered to result from the activity of a single central oscillator that involves the cerebello-thalamo-cortical pathway(Pinto et al., 2003; Raethjen and Deuschl, 2012). However,the efferent pathway of the central oscillation signal (COS),from the brain to the spinal cord, and the neuronal circuits within the spinal cord involved in the generation of essential tremor remain to be identi fied.
An efferent pathway that could potentially convey oscillation signals for essential tremor is the propriospinal system.The involvement of propriospinal neurons in the neural control of movements has been suggested in previous studies (Isa et al., 2006; Alstermark et al., 2007), possibly indirectlyviacorticospinal projections. Although the roles of propriospinal neurons in human movement control are unclear, a realistic virtual arm model with authentic spinal neural circuits (Lan and He, 2012; He et al., 2013; Niu et al., 2017) makes it possible to test hypotheses on the role of propriospinal neurons in generating subcortical movement patterns.
A previous computational study (Hao et al., 2013) suggested that Parkinsonian tremor is transmitted from the motor cortex to the peripheral neuromuscular system through the multisynaptic corticospinal pathway, which involves the propriospinal neuronal network (Alstermark et al., 2007). The study revealed that the propriospinal neuronal network plays an important role in generating alternating tremor patterns in antagonistic muscles from central oscillations in Parkinson’s disease patients. In clinical practice, essential tremor and Parkinson’s disease often cause diagnostic confusion because of overlapping similarities in symptoms (Jain et al., 2006).
In this study, we used a computational model of the corticospinal virtual arm (CS-VA) with propriospinal neurons(Alstermark et al., 2007) and spinal circuitry (Mileusnic et al., 2006; Mileusnic and Loeb, 2006) to investigate the corticospinal mechanisms involved in essential tremor. Several simulation experiments were performed to investigate: (I)the corticospinal efferent pathway related to essential tremor; (II) the role of the propriospinal neuronal network in producing essential tremor; and (III) the effect of peripheral parameters, such as re flex gain, on essential tremor. We also discussed the qualitative differences between our simulation results and data from patients with essential tremor in other clinical studies (Elble et al., 1987; Britton et al., 1994; Elble et al., 1994a, b; Gao, 2004; Heroux et al., 2006; Hellwig et al.,2009; Mostile et al., 2012).
Materials and Methods
Corticospinal-virtual arm model
In our previous studies (Lan and He, 2012; He et al., 2013,2015), we developed a computational model that simulates the movement of the human arm, and which includes details of the spinal circuitry thought to contribute to the neural control of movement. This virtual arm model successfully explains the functional significance of alpha-gamma coactivation during arm movements (Li et al., 2014, 2015).Here, we improve the original virtual arm model by adding corticospinal projections. This CS-VA model is a multiscale model which can compute central motor commands into peripheral neural signals and activate lower motor neurons(MNs) in the forelimbs of humans and other mammals. Simultaneously, the proprioceptive afferents inform the brain of the dynamic and kinematic states of the forelimbs and regulate the activation of MNs through spinal re flex circuitry. To implement these functions, the CS-VA model comprises three parts: the propriospinal neuronal network (Alstermark et al., 2007; Hao et al., 2013), spinal re flex circuitry(Mileusnic et al., 2006; Mileusnic and Loeb, 2006), and the virtual arm model (He et al., 2013) (Figure 1).
As shown in Figure 1, cortical motor commands are conveyed to α MNs through monosynaptic or multisynaptic pathways. Cortical motor commands are also delivered to γ static (γs, monosynaptic pathway) and γ dynamic (γd, multisynaptic pathway) MNs, which innervate muscle spindles.The movement commands of α dynamic MNs are integrated with the γ dynamic commands at the propriospinal neuronal network. The virtual muscles (Cheng et al., 2000) and spindles (Mileusnic et al., 2006) are activated by the outputs of α and γ MNs, respectively. The dynamic and kinematic movement trajectories are calculated using a biomechanical model of the arm. These trajectories are encoded in the primary and secondary afferents from the muscle spindles and Golgi tendon organs of the recruited muscles. The spinal re flex circuitry in Figure 2 regulates the outputs of α MNs according to the recurrent inhibition and re flex actions provided by the Ia, Iband IIafferents. The spinal re flex circuitry and virtual arm subsystems are based on real physiological data (Song et al., 2008a, b), and have been veri fied in previous studies (Song et al., 2008a; He et al., 2013). All the component models were integrated in the SIMULINKMATLAB(MathWorks, Natick, MA, USA) platform for simulation.
Six muscles were included in the virtual arm model, including three pairs of antagonistic muscles with two degrees of freedom. The pectoralis major (clavicle portion) and deltoid posterior were selected as the flexor and extensor of the shoulder joint, respectively. The brachialis (BS) and triceps lateral (Tlt) were selected for the elbow joint; and the biceps short head (Bsh) and triceps long head (Tlh) were selected as the bi-articular muscles (Figure 1).
Simulation experiment
Simulation design
Both the physiological and mechanical signals simulated by the CS-VA model were collected and analyzed. The simu-lated data were compared with experimental data from patients with essential tremor.

Table 1 Details of model initiation
We simulated a fast elbow extension movement using the CS-VA model. To focus on the origin of essential tremor,the simulation experiments were simplified into a pair of antagonistic muscles, the biceps and triceps, which controlled the elbow movement. In the simulation, the COS was considered always present. Hence, after the CS-VA was initialized and the virtual arm was in a stable resting state (see Table 1 for details of model initiation), one of the cortical oscillating commands was used to drive the CS-VA model in different stimulation experiments. After this, a set of pulses were passed to the α multisynaptic pathway in the CS-VA model to drive the model to perform a fast elbow extension,similar to what occurs in the real world. In this movement,the virtual arm changed its elbow angle and maintained this new posture for a while after the movement.
Simulation with different central commands
Four different sources of COS were simulated to identify the one producing the most realistic essential tremor. This experiment was performed because only a few studies found a strong association between the COS and limb electromyogram within the frequency range of essential tremor (Hellwig et al., 2001, 2003; Raethjen et al., 2007). This suggests that the route from COS to muscle activity might be convoluted.The four sources of COS were the following: α in the monosynaptic pathway (αmono), α in the multisynaptic pathway(αmulti), and γs and γdtracts. By matching the simulation results to the biomechanical features of essential tremor in the clinic, we should be able to identify the neural tract most likely to cause essential tremor.
αmonoas the COS: Evidence of direct cortico-motoneuronal connections was found in studies on the voluntary control of the hand (Lemon et al., 1998), and this pathway from the cortex is critical in controlling hand dexterity (Lemon, 2008;Isa et al., 2013). Thus, it was possible that the oscillation in the cortex was delivered through this pathway. The first set of stimulation experiments was performed using αmonoas the COS. In this case, the COS directly impinged on the α MN pool (Figure 1), which was further modulated by the spinal re flex involving Iaand Ibafferents, and by recurrent inhibition. The value of αmultiwas set at 0. The γssignal, which was placed directly upon the γ MN, was at a constant value. The γd, which was set at a constant bias value of 0.5, was placed into the propriospinal neuronal network and on the γ MN separately. The gains of the spinal reflex were set within a range used in other published studies (Lan and He, 2012;He et al., 2013). In the simulation, a recti fied sinusoid was selected to approximate the αmono, at a frequency of 4–7 Hz and an amplitude of 0.1–0.5. The αmonocould be described as follows:

whereA= 0.1, 0.2, 0.3, 0.4 and 0.5; ω = 4, 5, 6 and 7; and k= 1, 2, 3...
In the simulation experiments, formula 1 was also used for the other COSs.
αmultias the COS: Two descending pathways connect the brain to the MN pool–the direct and indirect pathways. The direct pathway connects the primary motor cortex to the MN pool, and was simulated in the first simulation experiment. In contrast, the indirect pathway is a multisynaptic pathway connecting the brain to MNsviathe propriospinal neural network. The presence of numerous indirect pathways from the brain to MNs has been shown in primates and humans (Alstermark et al., 2007; Riddle et al., 2009;Alstermark et al., 2011). These indirect pathways are mediated by intercalated neurons, such as segmental interneurons,propriospinal neurons and reticulospinal neurons. Propriospinal neurons are involved in the control of fine voluntary movements (Alstermark et al., 2011), as shown in experiments with monkeys. Hence, the COS could be transmitted through this pathway to produce essential tremor. We again chose formula 1 to approximate the COS from αmulti. The COS was modulated in the propriospinal neuronal network (Figure 2) by γdbefore impinging on the MN pool. The γdvalues used in the first simulation were also used here. The propriospinal neuronal process could be described as follows:


Figure 1 Corticospinal virtual arm (CS-VA) model used in this simulation study.

Figure 2 Model of the propriospinal neuronal (PN) network in the corticospinal pathways of one pair of antagonistic muscles.
where Pfand Peare the propriospinal neuronal outputs to the α MN of the flexor and extensor muscles, respectively;αmultiis the α signal in the multisynaptic pathway; dsand dfare the inhibition of the gains of γd; aeand afare the feedback gains of the Iaafferent onto the propriospinal neurons;and υeand υfare proportional to Iaand are afferent discharge frequencies of the spindles from the flexor and extensor muscles, respectively. The subscripts f and e are variables for the flexor and extensor muscles, respectively. This formula was obtained from a previous study (Hao et al., 2013) and was modi fied based on physiological evidence in other studies (Zehr et al., 2004; Zehr and Hundza, 2005; Alstermark et al., 2007; Alstermark and Isa, 2012). This model highlights the importance of spinal circuits in the modulation of muscle activity patterns.
γsas the COS: Both α and γ commands are known to be involved in normal voluntary movement. Based on the correlation of the behavior of these commands during midbrain stimulation with changes in the muscle spindle afferent responses to muscle stretching, the cortical γ commands were classified into two types–γ static (γs) and γ dynamic (γd)(Taylor et al., 2000). Some researchers consider γsto be the temporal template of the intended movement and afferent biasing through bag intrafusal fibers (Ellaway et al., 2002).According to this concept, if the COS was added to the γs,a tremor movement might occur. In the third simulation experiment, we used γs(the COS) to drive the model. This signal was directly passed onto the γ MN, similar to formula 1. In the CS-VA model, this signal could affect spindle feedback, which is the input to the α MN pool. Therefore, when the subject wants to move, tremor might result.
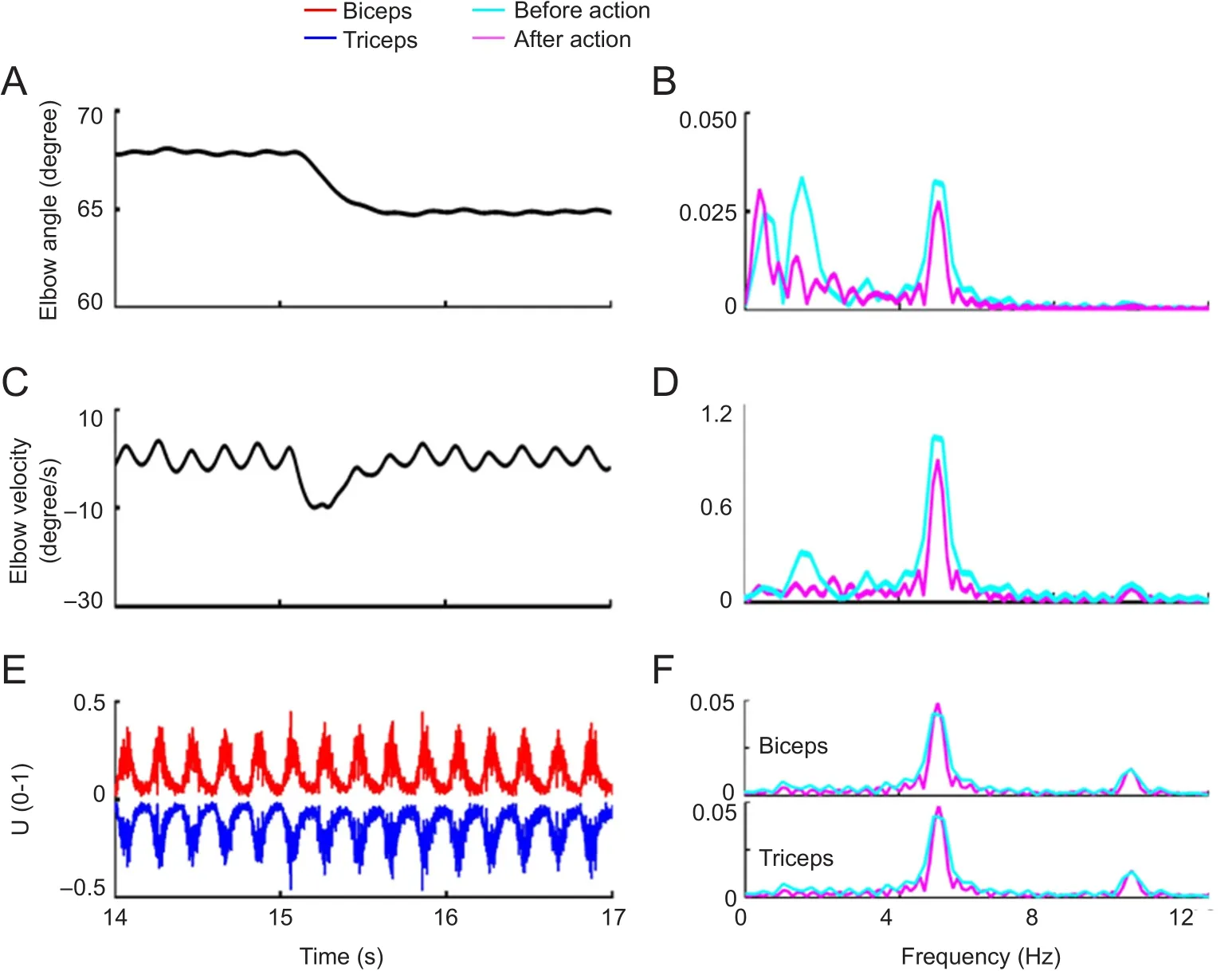
Figure 3 Features of movement and muscle activity in the simulation experiment (αmono).

Figure 4 Features of movement and muscle activity in the simulation experiment (αmulti).
γ dynamic as the COS: According to previous studies (Taylor et al., 2000, 2004, 2006; Alstermark et al., 2007), γ dynamic activity is related to movement control during locomotion in decerebrated cats. These studies show that γdfiring frequency increases unexpectedly from zero to maximum during the onset of muscle shortening, and then continues into the beginning of lengthening (Taylor et al., 2000). This observation suggests that γdserves an important function in movement control. Thus, γ dynamic commands might be associated with the COS at the essential tremor frequency.In the CS-VS model, similar to the second simulation, the γdwas entered into the propriospinal neuronal network and the γ MN, but the value was a combination of the bias value of 0.5 and the recti fied sinusoidal wave described in formula 1. According to formula 2, the inhibition is not a constant value, but rather a fluctuating value. If the movement command is entered into propriospinal neurons, the abnormal γdcan modulate the movement command in an oscillating way, affecting the α MN pool indirectly. In addition, the COS was entered into the γ MN to adjust the spindle output,which is also related to the MN pool. Hence, this might be another cause of tremor.
Simulation with different stretch re flex gains
We also examined factors that could potentially affect the amplitude and frequency of essential tremor. Based on data from essential tremor patients (Elble et al., 1994b; Matsumoto et al., 1999; Gao, 2004; Mostile et al., 2012), we proposed the following: (I) The frequency of essential tremor is only determined by the COS, as shown by corticomuscular coherence studies (Hellwig et al., 2001; Raethjen et al., 2007); and (II)The amplitude of essential tremor is related to two components, the COS and the mechanical re flex (Elble et al., 1987).Previous studies have examined the contribution of the mechanical re flex to essential tremor by varying the inertial loading. However, obtaining accurate reflex gain values in human movements is difficult, and adjusting these values to determine a relationship between the amplitude of essential tremor and the stretch re flex is even more difficult. The main re flexes in our CS-VA model can be conveniently adjusted to determine biomechanical changes. Hence, in another set of simulations, stretch reflex gains were adjusted from 0.1 to 0.5. For each stretch re flex gain value, both the amplitude and frequency of the COS were adjusted. From the biomechanical data obtained with our model, we could quantitatively analyze the relationship between the stretch gain and the amplitude of the essential tremor.
Results
Tremor generated by the COS
With the hypothesis that the central oscillation is the originating cause of essential tremor, the abnormal joint angles and muscle behavior were calculated from the CS-VA model(plotted in Figures 3–6), and the spectra were analyzed. In these figures, all COSs were set at a frequency of 5 Hz and an amplitude of 0.3.
In the first experiment, αmonowas set as the COS pathway.Under this condition, the elbow angle always oscillated before and after the action (Figure 3A, C). The tremor amplitude was approximately 0.3 degree, and the amplitude decreased after the action occurred, according to the movement and velocity spectra (Figure 3B, D). The biceps and triceps were activated synchronously by this type of COS(Figure 3E), and the two muscles had nearly the same amplitude of activity during co-contraction, according to the spectral analysis (Figure 3F, G). The movement and velocity spectra of the elbow movement before and after the action did not show any obvious difference, contradicting the widely-held concept that essential tremor is an action tremor.
In the second experiment, αmultiwas taken as the COS pathway. As shown in Figure 4A, the elbow angle did not change at the resting state, and muscle activity showed no corresponding changes (Figure 4B). A tremor occurred after the elbow extension. The tremor amplitude was very small,but the spectral analysis of the movement revealed the existence of two peaks, namely, one at 5 Hz and another at 1 Hz,which may have caused confusion in the observation of the tremor. However, the spectral analysis showed an obvious single peak at the tremor frequency (5 Hz) in elbow velocity after the extension movement (Figure 4D). The maximum value of the spectrum at 5 Hz was over 0.75, re flecting the oscillating movement. After the action, the muscle activity was in a co-contraction firing pattern of antagonistic muscles, caused by the α COS in the multisynaptic pathway. This firing pattern caused elbow oscillation, but the tremor amplitude was very small. The spectrum of the muscle activity demonstrated the co-contraction pattern at 5 Hz. The kinetic features before and after the elbow action agreed with some characteristics of essential tremor in clinical studies,and the muscle activity had a co-contraction pattern similar to the electromyogram in essential tremor patients.
In the third experiment, γswas taken as the initial cause of essential tremor. The commands for postural control showed an abnormal firing pattern and reached the γ MNs directly. The elbow angle showed very small tremor after the movement (Figure 5A). Muscle activity was enhanced after the action (Figure 5E), but the spectral analyses of the two muscles showed that muscle activity was neither synchronous nor alternating. In addition, the muscle activity spectra showed that there was no difference in tremor frequency,whether the arm moved or not (Figure 5F, G).
The last experiment used hdas the COS. Before it reached the γ MN, d was conveyed through the propriospinal neuronal network. Similar to the second experiment, no tremor was found at resting state, and the tremor behavior was observed after the action (Figure 6A). Compared with the second experiment, the tremor in the elbow trace in Figure 6A was easily detectable because tremor amplitude was relatively high.The elbow angle oscillated between 76° and 77° at a frequency of 5 Hz, the same as the COS frequency. The biggest difference between the simulation experiments was that muscle activity showed an alternating pattern in the present simulation(Figure 6E). The spectra showed that muscle activity peaked at 5 Hz, the tremor frequency, in both the biceps and triceps,despite different activity levels (Figure 6F, G).
In fluence of the oscillating parameters of COS on the simulated tremor
Our current simulation experiments indicate that αmultiand αdare the COSs underlying essential tremor. We also examined the COS parameters that in fluence the amplitude of elbow oscillation,i.e. the amplitude and frequency. Figure 7 shows the relationships between these COS parameters and the elbow oscillating amplitude. The amplitude of umultiand ndranged from 0.1 to 0.5. The frequency of rmultiand αdwas adjusted from 4 to 7 Hz. Regardless of the COS (whether αmultior od), the same regulatory effect was found–frequen-cy had a greater impact on tremor amplitude. The lower the COS frequency, the larger the tremor amplitude. When the frequency was reduced from 4 to 5 Hz, the tremor amplitude decreased drastically. However, the tremor amplitude changed less in the range between 5 and 6 Hz. Moreover, the tremor disappeared when COS in the tmultipathway was at 7 Hz. As the COS was delivered through the αmultipathway, the tremor amplitude first increased along with the increase in COS amplitude from 0.1 to 0.4, and reached the maximum value when the COS amplitude was 0.4, and then decreased as COS amplitude increased from 0.4 to 0.5 (Figure 7A).The tremor amplitude monotonously increased when COS was wd, as shown in Figure 7B. These findings show that whether the COS is from the αmultior hdpathway, the tremor frequency is determined by the frequency of the COS. COS amplitude only in fluences tremor amplitude.
Stretch reflex-caused oscillation contributed to essential tremor
Figure 8 shows the effect of stretch re flex gain on the amplitude. Given that the COS was from the tmultipathway, with an amplitude of 0.3, the tremor amplitude was only affected by the COS frequency. Increasing stretch re flex gain did not influence tremor amplitude, as shown in Figure 8A. When the COS frequency was fixed at 5 Hz, and the COS amplitude was within 0.3, increasing stretch reflex gain did not affect tremor amplitude, as shown in Figure 8B. However, increasing stretch reflex gain reduced tremor amplitude when the COS amplitudes were 0.4 and 0.5 (Figure 8B). As shown in Figure 8C, except when the d was at 4 Hz (the wdamplitude was maintained at 0.3), increasing stretch re flex gain weakly affected tremor amplitude, compared with the effect of tdfrequency. The amplitude of fdand the stretch re flex gain had similar effects on the tremor amplitude (Figure 8D).
Discussion
In this study, a set of simulation experiments were performed to identify the efferent spinal pathway conveying the COS from the brain, and whether the propriospinal neuronal network is involved in the generation of essential tremor.In our simulation experiments, nmonoand αswere excluded as the COS pathways causing essential tremor, because the kinetic and physiological characteristics did not match the clinical features of essential tremor.
Essential tremor can be classi fied into two types, based on clinical assessment: type A and type B (Deuschl et al., 1987;Milanov, 2001). Deuschl et al. showed that in type A essential tremor, electromyography shows synchronous activity in the antagonistic muscles, whereas electromyography shows alternating activity in the antagonists in type B essential tremor. Moreover, in Milanov’s study, more than 200 essential tremor patients were recruited, and similar results were obtained. Correlating the clinical characteristics with the simulation results suggests that the tremor caused by ymultiis most likely the type A variety. In the second experiment, the tremor caused by COS from the αmultipathway did not appear in the resting state and had low amplitude after elbow extension. These kinetic features are similar to the clinical characteristics. In addition, the muscle activity was of a co-contraction pattern. The tremor caused by the γdpathway COS in the simulation experiments can be identi fied as type B using the same method. The kinetic characteristics of the elbow and muscle activities matched the signs of type B essential tremor, with the tremor amplitude larger than in type A, with alternating muscle activity (Britton et al., 1994).
Both types of COS were modulated by the propriospinal neuronal network in the spinal cord. This shows the importance of the propriospinal neuronal network in controlling human movement. The propriospinal neuronal network,which serves a gating function, modulated the central commands from the brain to the spinal cord. Our findings could be related to observations of the corticospinal tract in patients with ET and PD (Lu et al., 2016). In terms of movement control angle, essential tremor can be identi fied clearly at the spinal cord level. In type A essential tremor, propriospinal neurons gate the αmultiCOS. Indeed, essential tremor patients show no tremor in the resting state. However, when COS was added to the movement control signals, it surmounted the gating by the propriospinal neurons to reach the α MNs. This explains the occurrence of tremors when an action is performed. In type B essential tremor, the γdCOS overcame the propriospinal neuronal gating function by lowering the gating value. At resting state, α is too small to cause tremor, even when the propriospinal neuronal gating function is impaired. When a patient wants to move or maintain a posture, and α is sufficiently large, propriospinal neuronal gating would be unable to prevent tremor.
In previous studies, we used a similar model that included the propriospinal neuronal network to simulate Parkinson’s disease tremors. The simulation results matched the clinical features of Parkinson’s disease tremors. Thus, the propriospinal neuronal network is involved in both tremors. Essential tremor and Parkinson’s disease tremors share the same circuits in the spinal cord. Indeed, the signs of both tremors are similar and overlapping, and distinguishing them is clinically difficult.
In a previous study (Elble et al., 1987), the stretch re flex in essential tremor was examined, and the tremor was found to be in fluenced by two components–COS and the mechanical reflex. However, tremor frequency was determined by the central oscillation. Our current results are in agreement with these observations. COS frequency not only impacted tremor frequency, but also strongly influenced tremor amplitude in our simulation experiments, regardless of the COS pathway (fmultior rd) (Figures 7 and 8). An advantage of the CS-VA model, the in fluence of the stretch re flex gain can be directly investigated in the simulation. In type A essential tremor, the stretch re flex gain had none or little effect on the tremor amplitude, as shown in Figure 8. In type B essential tremor, increasing stretch re flex gain aggravated the tremor amplitude until the tremor amplitude was over 0.035. Our results provide novel insight into the neurological basis of essential tremor.

Figure 5 Features of movement and muscle activity in the simulation experiment (γs).
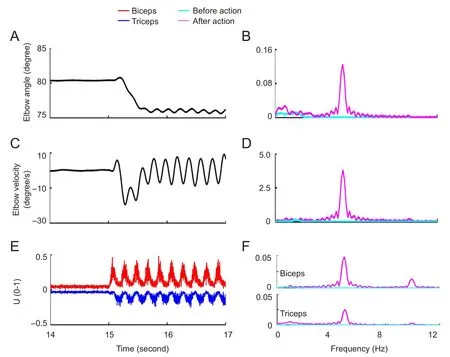
Figure 6 Features of movement and muscle activity in the simulation experiment (γd).
In our study, essential tremor was elicited by the COS through the multisynaptic corticospinal pathway involving the propriospinal neuronal network. The propriospinal neuron plays an important role in gating the COS at resting state, preventing the COS from passing down to the peripheral neuromuscular system. During the performance of an active task, cortical inhibition of the propriospinal neuron is reduced or removed by task commands, and the COS can be transmitted to the peripheral neuromuscular system freely, causing a tremor activity that accompanies the task.The simulation analyses here suggest that the propriospinal neuronal network plays a role in the gating of the cortical oscillation signal, preventing it from reaching the peripheral muscles in the resting state, but allowing it to pass to the muscles during an active task.
In conclusion, COS frequency is the major factor influencing tremor amplitude, with a greater impact than COS amplitude or stretch re flex gain. Computational modeling is an effective method of simulating essential tremor and identifying abnormal neural networks.
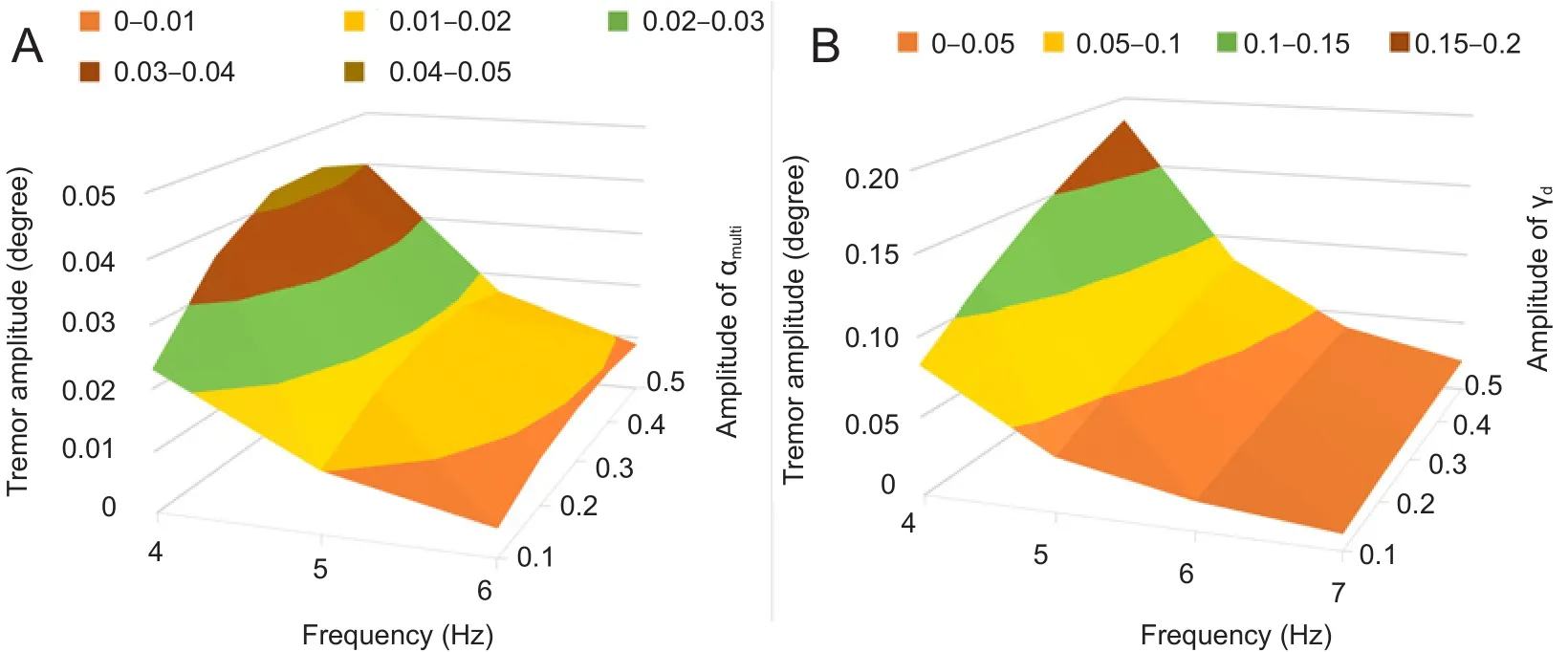
Figure 7 In fluence of the frequency and amplitude of the central oscillation signal(COS) on tremor amplitude.

Figure 8 Relationship between tremor amplitude and re flex gain.
Author contributions:HEQ constructed the simulation, collected the data, analyzed the results, and participated in the production of the manuscript. CMN contributed to the design of the experiment, analyzed the results, and produced part of the manuscript. SL, MZH and ZXH contributed to the modeling, simulation and data collection. QX contributed the design and analysis of the study. NL provided the original concept and design of the study, analyzed the results, and jointly produced the manuscript. All authors approved the final version of the paper.
Con flicts of interest:None declared.
Data sharing statement:Datasets analyzed during the current study are available from the corresponding author on reasonable request.
Plagiarism check:Checked twice by iThenticate.
Peer review:Externally peer reviewed.
Open access statement:This is an open access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 3.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as the author is credited and the new creations are licensed under identical terms.
Alstermark B, Isa T (2012) Circuits for skilled reaching and grasping.Annu Rev Neurosci 35:559-578.
Alstermark B, Isa T, Pettersson LG, Sasaki S (2007) The C3-C4 propriospinal system in the cat and monkey: a spinal pre-motoneuronal centre for voluntary motor control. Acta Physiol 189:123-140.
Alstermark B, Pettersson LG, Nishimura Y, Yoshino-Saito K, Tsuboi F,Takahashi M, Isa T (2011) Motor command for precision grip in the macaque monkey can be mediated by spinal interneurons. J Neurophysiol 106:122-126.
Brennan KC, Jurewicz EC, Ford B, Pullman SL, Louis ED (2002) Is essential tremor predominantly a kinetic or a postural tremor? A clinical and electrophysiological study. Mov Disord 17:313-316.
Britton TC, Thompson PD, Day BL, Rothwell JC, Findley LJ, Marsden CD (1994) Rapid wrist movements in patients with essential tremor.The critical role of the second agonist burst. Brain 117(Pt 1) :39-47.
Cheng EJ, Brown IE, Loeb GE (2000) Virtual muscle: a computational approach to understanding the effects of muscle properties on motor control. J Neurosci Methods 101:117-130.
Deuschl G, Bain P, Brin M (1998) Consensus statement of the Movement Disorder Society on Tremor. Ad Hoc Scientific Committee.Mov Disord 13 Suppl 3:2-23.
Deuschl G, Lucking CH, Schenck E (1987) Essential tremor: electrophysiological and pharmacological evidence for a subdivision. J Neurol Neurosurg Psychiatry 50:1435-1441.
Elble RJ, Higgins C, Hughes L (1994a) Essential tremor entrains rapid voluntary movements. Exp Neurol 126:138-143.
Elble RJ, Higgins C, Leffler K, Hughes L (1994b) Factors in fluencing the amplitude and frequency of essential tremor. Mov Disord 9:589-596.
Elble RJ, Higgins C, Moody CJ (1987) Stretch reflex oscillations and essential tremor. J Neurol Neurosurg Psychiatry 50:691-698.
Ellaway P, Taylor A, Durbaba R, Rawlinson S (2002) Role of the fusimotor system in locomotion. Sensorimotor Control Mov Posture 508:335-342.
Gao JB (2004) Analysis of amplitude and frequency variations of essential and Parkinsonian tremors. Med Biol Eng Comput 42:345-349.
Hallett M (2014) Tremor: pathophysiology. Park Relat Disord 20 Suppl 1:S118-22.
Halliday DM, Conway BA, Farmer SF, Shahani U, Russell AJ, Rosenberg JR (2000) Coherence between low-frequency activation of the motor cortex and tremor in patients with essential tremor. Lancet 355:1149-1153.
Hao M, He X, Xiao Q, Alstermark B, Lan N (2013) Corticomuscular transmission of tremor signals by propriospinal neurons in Parkinson’s disease. PLoS One 8:e79829.
He X, Du YF, Lan N (2013) Evaluation of feedforward and feedback contributions to hand stiffness and variability in multijoint arm control. IEEE Trans Neural Syst Rehabil Eng 21:634-647.
He X, Hao M-Z, Wei M, Xiao Q, Lan N (2015) Contribution of inter-muscular synchronization in the modulation of tremor intensity in Parkinson’s disease. J Neuroeng Rehabil 12:1.
Hellriegel H, Schulz EM, Siebner HR, Deuschl G, Raethjen JH (2012)Continuous theta-burst stimulation of the primary motor cortex in essential tremor. Clin Neurophysiol 123:1010-1015.
Hellwig B, Haussler S, Schelter B, Lauk M, Guschlbauer B, Timmer J,Lucking CH (2001) Tremor-correlated cortical activity in essential tremor. Lancet 357:519-523.
Hellwig B, Mund P, Schelter B, Guschlbauer B, Timmer J, Lucking CH(2009) A longitudinal study of tremor frequencies in Parkinson’s disease and essential tremor. Clin Neurophysiol 120:431-435.
Hellwig B, Schelter B, Guschlbauer B, Timmer J, Lucking CH (2003)Dynamic synchronisation of central oscillators in essential tremor.Clin Neurophysiol 114:1462-1467.
Heroux ME, Parisi SL, Larocerie-Salgado J, Norman KE (2006) Upper-extremity disability in essential tremor. Arch Phys Med Rehabil 87:661-670.
Isa T, Kinoshita M, Nishimura Y (2013) Role of direct vs. indirect pathways from the motor cortex to spinal motoneurons in the control of hand dexterity. Front Neurol 4:191.
Isa T, Ohki Y, Seki K, Alstermark B (2006) Properties of propriospinal neurons in the C3-C4 segments mediating disynaptic pyramidal excitation to forelimb motoneurons in the macaque monkey. J Neurophysiol 95:3674-3685.
Jain S, Lo SE, Louis ED (2006) Common misdiagnosis of a common neurological disorder: how are we misdiagnosing essential tremor?Arch Neurol 63:1100-1104.
Lan N, He X (2012) Fusimotor control of spindle sensitivity regulates central and peripheral coding of joint angles. Front Comput Neurosci 6:66.
Lemon RN (2008) Descending pathways in motor control. Annu Rev Neurosci 31:195-218.
Lemon RN, Baker SN, Davis JA, Kirkwood PA, Maier MA, Yang HS(1998) The importance of the cortico-motoneuronal system for control of grasp. Novartis Found Symp 218:202-215; discussion 215-218.
Li S, He X, Lan N (2014) Modular control of movement and posture by the corticospinal alpha-gamma motor systems. In, pp 4079-4082.IEEE.
Li S, Zhuang C, Hao M, He X, Marquez JC, Niu CM, Lan N (2015)Coordinated alpha and gamma control of muscles and spindles in movement and posture. Front Comput Neurosci 9:122.
Louis ED (2005) Essential tremor. Lancet Neurol 4:100-110.
Lu MK, Chen CM, Duann JR, Ziemann U, Chen JC, Chiou SM, Tsai C-H (2016) Investigation of motor cortical plasticity and corticospinal tract diffusion tensor imaging in patients with Parkinsons disease and essential tremor. PLoS One 11:e0162265.
Martinelli P, Gabellini AS, Gulli MR, Lugaresi E (1987) Different clinical features of essential tremor: a 200-patient study. Acta Neurol Scand 75:106-111.
Matsumoto JY, Dodick DW, Stevens LN, Newman RC, Caskey PE,Fjerstad W (1999) Three-dimensional measurement of essential tremor. Mov Disord 14:288-294.
Milanov I (2001) Electromyographic differentiation of tremors. Clin Neurophysiol 112:1626-1632.
Mileusnic MP, Brown IE, Lan N, Loeb GE (2006) Mathematical models of proprioceptors. I. Control and transduction in the muscle spindle.J Neurophysiol 96:1772-1788.
Mileusnic MP, Loeb GE (2006) Mathematical models of proprioceptors. II. Structure and function of the Golgi tendon organ. J Neurophysiol 96:1789-1802.
Mostile G, Fekete R, Giuffrida JP, Yaltho T, Davidson A, Nicoletti A,Zappia M, Jankovic J (2012) Amplitude fluctuations in essential tremor. Park Relat Disord 18:859-863.
Niu CM, Jalaleddini K, Sohn WJ, Rocamora J, Sanger TD, Valero-Cuevas FJ (2017) Neuromorphic meets neuromechanics, part I: the methodology and implementation. J Neural Eng 14:025001.
Pinto AD, Lang AE, Chen R (2003) The cerebellothalamocortical pathway in essential tremor. Neurology 60:1985-1987.
Raethjen J, Deuschl G (2012) The oscillating central network of Essential tremor. Clin Neurophysiol 123:61-64.
Raethjen J, Govindan RB, Kopper F, Muthuraman M, Deuschl G (2007)Cortical involvement in the generation of essential tremor. J Neurophysiol 97:3219-3228.
Riddle CN, Edgley SA, Baker SN (2009) Direct and indirect connections with upper limb motoneurons from the primate reticulospinal tract. J Neurosci 29:4993-4999.
Song D, Lan N, Loeb GE, Gordon J (2008a) Model-based sensorimotor integration for multi-joint control: development of a virtual arm model. Ann Biomed Eng 36:1033-1048.
Song D, Raphael G, Lan N, Loeb GE (2008b) Computationally efficient models of neuromuscular recruitment and mechanics. J Neural Eng 5:175-184.
Taylor A, Durbaba R, Ellaway PH (2004) Direct and indirect assessment of gamma-motor firing patterns. Can J Physiol Pharmacol 82:793-802.
Taylor A, Durbaba R, Ellaway PH, Rawlinson S (2006) Static and dynamic gamma-motor output to ankle flexor muscles during locomotion in the decerebrate cat. J Physiol 571:711-723.
Taylor A, Ellaway PH, Durbaba R, Rawlinson S (2000) Distinctive patterns of static and dynamic gamma motor activity during locomotion in the decerebrate cat. J Physiol 529 Pt 3:825-836.
Zehr EP, Carroll TJ, Chua R, Collins DF, Frigon A, Haridas C, Hundza SR, Thompson AK (2004) Possible contributions of CPG activity to the control of rhythmic human arm movement. Can J Physiol Pharmacol 82:556-568.
Zehr EP, Hundza SR (2005) Forward and backward arm cycling are regulated by equivalent neural mechanisms. J Neurophysiol 93:633-640.
Zhao J, Yang B, Li JQ, Yan JH, Zhu YH (2011) Application of shifted band-limited multiple fourier linear combiner in pre-estimation and filtering of human tremor. Zhongguo Zuzhi Gongcheng Yanjiu 15:4837-4840.
How to cite this article:Qu HE, Niu CM, Li S, Hao MZ, Hu ZX, Xie Q, Lan N (2017) Neural computational modeling reveals a major role of corticospinal gating of central oscillations in the generation of essential tremor. Neural Regen Res 12(12):2035-2044.
Funding: This work was supported in part by the National Natural Science Foundation of China, No. 61361160415, 81271684, 81501570; the Major State Basic Research Development of China (973 Program), No. 2011CB013304; the Medicine-Engineering Interdisciplinary Research Grant from Shanghai Jiao Tong University in China, No. YG2014ZD09; and a grant from the Youth Eastern Scholar Program at Shanghai Institutions of Higher Learning in China, No. QD2015007.
Graphical Abstract

*Correspondence to:Ning Lan, Ph.D. or Qing Xie,M.D., ninglan@sjtu.edu.cn or ruijin_xq@163.com.
#These authors contributed equally to this paper.
orcid:0000-0001-6061-5419(Ning Lan)0000-0002-3298-1072(Qing Xie)
10.4103/1673-5374.221161
2017-11-05
The authors thank Drs. Yong Bao and Li-jun Cui for their help during the discussion.
Copyedited by Patel B, Frenchman B, Wang J, Li CH, Qiu Y, Song LP, Zhao M
杂志排行

中国神经再生研究(英文版)的其它文章
- Roles of neural stem cells in the repair of peripheral nerve injury
- Advanced diffusion-weighted magnetic resonance imaging in the evaluation of white matter axons in patients with idiopathic normal pressure hydrocephalus
- The contribution of oligodendrocytes and oligodendrocyte progenitor cells to central nervous system repair in multiple sclerosis: perspectives for remyelination therapeutic strategies
- MicroRNAs in Parkinson’s disease and emerging therapeutic targets
- Surgical reconstruction of spinal cord circuit provides functional return in humans
- Environmental cues determine the fate of astrocytes after spinal cord injury