气候变化背景下1964—2015年秦岭植物物候变化
2018-01-04邓晨晖白红英翟丹平黄晓月贺映娜
邓晨晖,白红英,*,翟丹平,高 山,黄晓月,孟 清,贺映娜
1 西北大学城市与环境学院,西安 710127 2 西安市气象局,西安 710016
气候变化背景下1964—2015年秦岭植物物候变化
邓晨晖1,白红英1,*,翟丹平1,高 山2,黄晓月1,孟 清1,贺映娜1
1 西北大学城市与环境学院,西安 710127 2 西安市气象局,西安 710016
以1964—2015年物候观测数据为基础,选取17种含乔木、灌木及藤本树种为研究对象,分析探讨了气候变化背景下秦岭地区植物物候变化规律及其差异性。结果表明:①52年来,秦岭地区物候始期普遍呈提前趋势,提前速率1.2d/10a,物候末期普遍呈推迟趋势,推迟速率3.5d/10a,物候生长期普遍延长;②秦岭地区物候突变发生于20世纪80年代,始期于1985年,末期于1984年。突变后,物候特征发生了显著变化,始期的提前速率较突变前显著加快,末期由突变前的提前趋势转变为极显著的推迟趋势,且变化速率和显著性均高于始期;始期与末期变化均表现出“趋同效应”;物候年代际变化趋势显示,始期自2001—2005年起提前速率减缓,植物对气候变化的响应表现出适应性及滞后性。③秦岭物候变化存在树种差异,3大类树种始期的提前速率呈藤本、乔木、灌木依次增大,而末期的推迟速率则呈藤本、灌木、乔木依次减小。④秦岭物候变化存在南北差异,北坡始期的提前速率均高于南坡,而南坡末期的推迟速率均高于北坡。
植物物候;变化特征;突变;差异性;秦岭地区
物候是受环境影响而出现的以年为周期的自然现象[1-3]。自然物候记录是全球环境变化最直接和最有效的证据[4],物候现象不仅能反映自然季节的变化,还能指示生态系统对全球环境变化的响应和适应,因而也被视为大自然的“语言”[1]和全球变化的“诊断指纹”[5-6]。植物物候作为气候变化与自然环境变化共同作用于植物体上的直接表现[7],其变化也是生物圈对气候变暖反应最敏感和最明显的指示[8-9],已成为全球变化研究重点关注对象[10]。
IPCC第五次评估报告指出,全球近130a(1880—2012年)升温0.85℃[11],1951—2012年平均地表温度的升温率0.12℃/10a[12];中国近50年地表气温加速上升,1951—2007年的升温率达0.23℃/10a,气候变暖十分明显[13]。大量研究表明,随着气候暖化,许多物候现象发生了明显变化。近几十年来,持续增温使得北半球不同区域植物春季物候呈提前趋势,秋季物候呈推迟趋势,生长季呈延长趋势[14-25]。国际物候园观测资料显示欧洲中西部地区现在的春季物候比50a前提前10—20d,变化速率在物种间、地区间和年际间有差异[26];地中海地区大多数落叶植物生长期比50a前提早了16d,落叶期推迟了13d[27];北美大部分地区春季物候平均提前了5—6d[28]。我国的物候观测资料也表明,过去50年温带季风区木本植物始花期以1.93d/10a的速度显著提前[29];东部地区木本植物秋季叶全变色期整体表现为推迟趋势,且1980s后推迟趋势明显[30];东北地区木本植物展叶初期提前速率为0.23d/a,枯黄初期推后0.19d/a,生长季延长速率0.30d/a[31];华北地区1980s以后春季物候亦呈大幅度提前趋势[32];西北荒漠区植物也表现出春季物候期提前,落叶末期推迟[33],且1985年后生长季的增加趋势达到显著性[34]。上述研究表明,全球及中国的植物物候表现出与气候变暖协同变化的特征[35-36]且存在区域差异。
山地被认为是全球变化的前哨,秦岭作为我国具有独特意义的地域单元,生态敏感而脆弱,近半个世纪以来气温呈显著上升趋势[37]。然而,由于受物候数据的限制,有关秦岭地区植物物候研究相对较少,仅白红英[9]、贺映娜[38]、马新萍等[39]基于地面观测数据和遥感数据初步分析了秦岭植物物候始末期及遥感生长季的变化;夏浩铭等[40]利用遥感资料研究了秦岭森林物候随海拔变化的特征。本文采用最新资料在前人的基础上进一步深入分析了:随着气候变化,近50多年来秦岭地区植物物候正在发生着怎样的变化?物候始期与末期是否提前或推迟?物候变化是否存在转折点?树种和空间上存在怎样的差异性?旨在揭示秦岭物候的变化规律,以期为农业生产结构调整与秦岭森林植被生态系统管理提供理论依据。
1 数据与方法
1.1 研究区概况
本文以狭义的秦岭为研究区(图1),即秦岭腹地,位于陕西南部,渭河以南,汉江以北,东西以陕西省省界为界。105°30′—110°05′E,32°40′—34°35′N,总面积约6.19万km2,约占陕西省总土地面积30%,海拔高度195—3771.2m。

图1 研究区地理位置Fig.1 The geographical location of the research area
秦岭南北气候差异明显,表现为北干南湿。北坡属于暖温带半湿润气候,广泛分布暖温带落叶阔叶林;南坡为北亚热带湿润气候,分布着北亚热带常绿阔叶—落叶阔叶混交林。
1.2 数据来源与处理
1.2.1 物候数据来源
西安站的物候数据来源于国家地球系统科学数据共享平台(http://www.geodata.cn)及西安植物园,咸阳站、武功站、渭南站、商洛站、安康站、城固站的物候数据来源于陕西省气象局。
1.2.2 物候数据选取与处理
本文筛选了研究区内分布广泛且记录完善,时间序列长又可代表秦岭地区气候特征的7种木本植物作为研究对象;考虑到空间分布对物候变化的影响,又收集了20世纪80年代后建立的物候观测站中记录时间序列相对较长的10个树种资料,共计17个树种(表1)。
采用儒略日(Julian days)换算法将逐年物候期出现的日期转化为距当年1月1日的实际天数,即为年序列累计天数,得到物候期的时间序列,以此计算物候期提前或推迟的天数。在物候期的选取上,为了与后期研究以遥感监测所表征的生长季始期(返青期)和末期(枯黄期)相对应,故将“展叶盛期”作为研究的“物候始期”,“叶全部变色期”作为研究的“物候末期”,从始期开始至末期结束的时间长度为研究的“物候生长期”。此外,泡桐因属于先开花后展叶植物,故将“开花盛期”作为其“物候始期”。
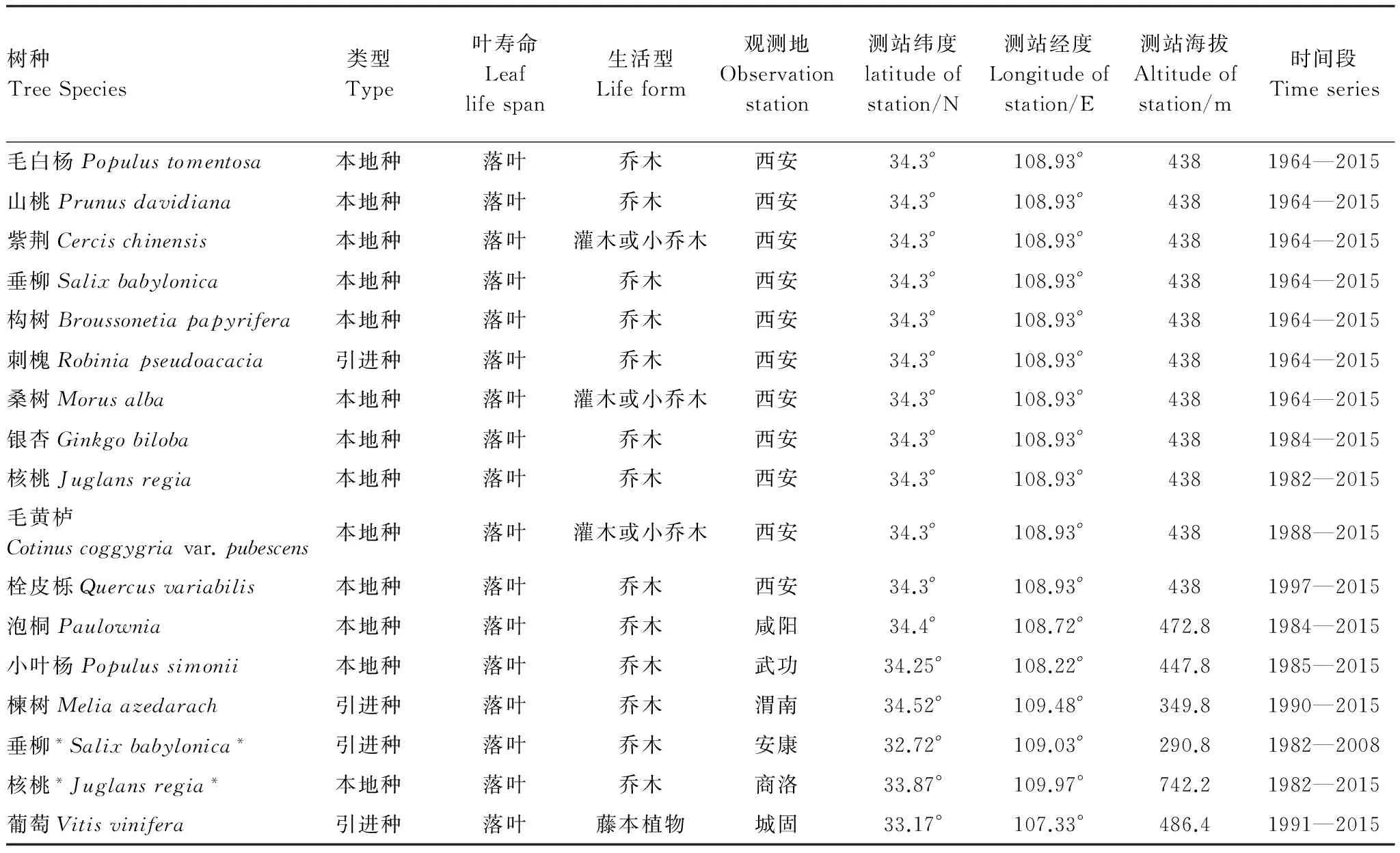
表1 秦岭地区物候研究主要观测树种
核桃与垂柳带*为南坡树种,不带*为北坡树种
对于有多个年份(如西安站1966年、1969—1972年、1989年、1992年)数据记录中断或缺失的不做处理,对于个别树种物候期有漏缺的,根据前后各3年的数据均值进行插补。
1.2.3 气象数据来源
研究区内32个气象台站1959—2015年逐月平均气温与降水量数据来源于中国气象数据科学共享服务网(http://data.cma.cn)、陕西省气象局和西安市气象局,用于分析研究区的气候特征和水热条件。
1.3 研究方法
1.3.1 物候变化趋势分析
利用线性拟合方法分析物候期的年际变化趋势,统计物候期提前和推迟趋势的变化速率,并对变化趋势进行显著性检验,进而分析秦岭52年来物候始期与末期的年际变化趋势特征。
1.3.2 物候突变分析
基于最小二乘法原理的二次曲线拟合方法,在物候期的趋势变化图中建立二次拟合曲线及曲线方程,通过一阶求导得出曲线斜率变化线,曲线斜率线与0斜率线相交的点所对应的年份为物候突变年份,即物候数据序列的突变点。
2 结果与分析
2.1 秦岭地区气候特征及水热条件
图2为1959—2015年秦岭地区气温和降水年平均距平的变化序列。显示,50多年来,秦岭地区的地表气温加速上升,年均温的升温率为0.20℃/10a(P<0.01),气候变暖明显。北坡的线性升温率达0.21℃/10a(P<0.01),南坡为0.18℃/10a(P<0.01),北坡的暖化趋势较南坡明显。从气温距平5年滑动曲线看,气温在80年代初急剧下降,1985年开始呈稳定上升趋势。秦岭地区的降水量整体上未见显著趋势性变化,但区域变化趋势差异明显,北坡的干旱趋势明显高于南坡。从降水量距平5年滑动曲线看,20世纪80年代为多雨期,90年代为少雨期,2000年以来降水有所增加。
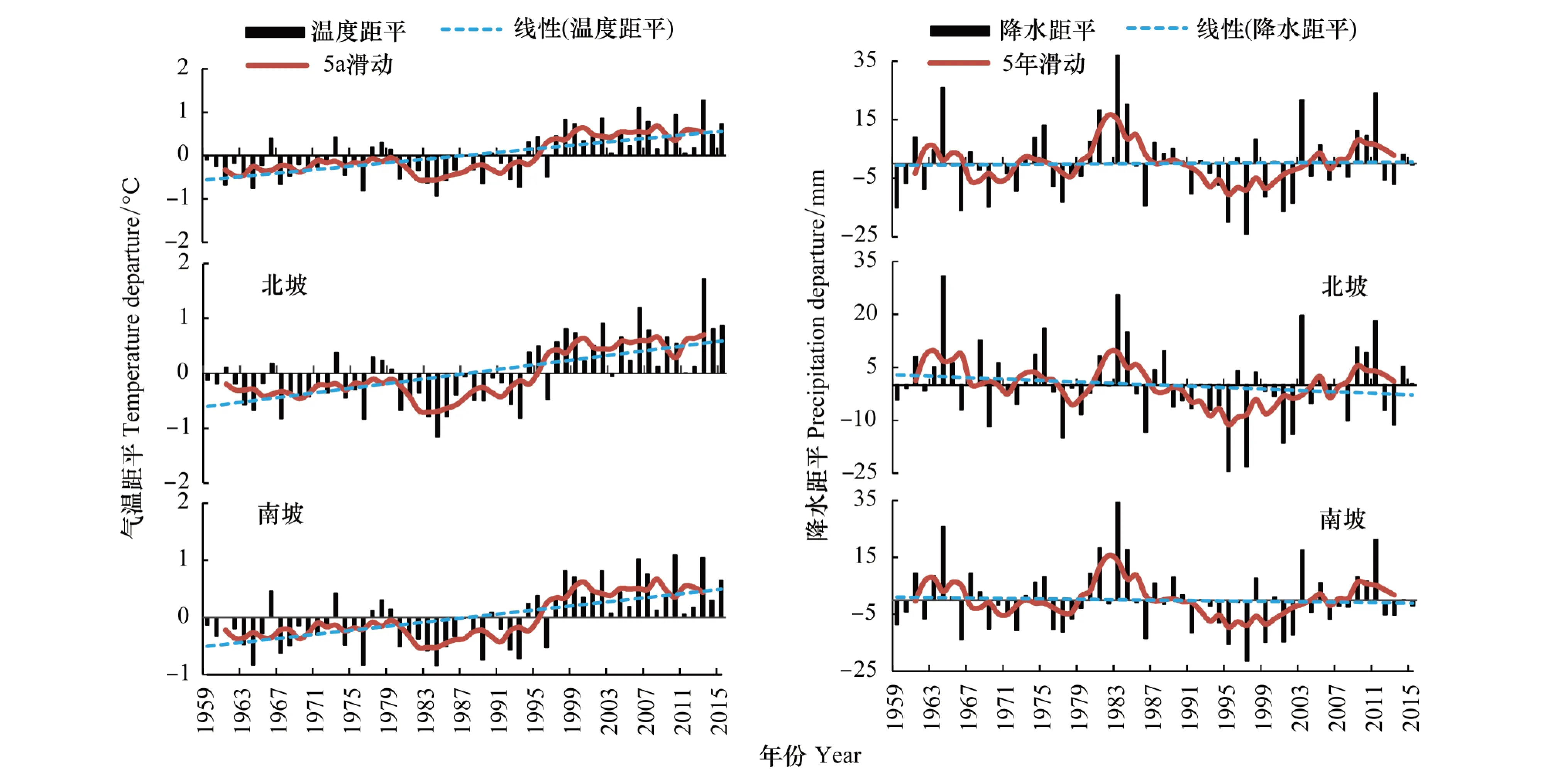
图2 1959—2015年秦岭年均气温距平与降水距平变化Fig.2 The departure change of annual temperature and precipitation in the Qinling Mountains during 1959—2015
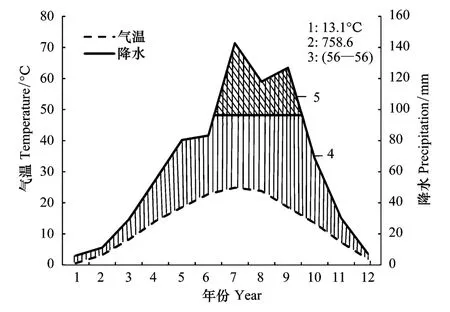
图3 秦岭地区1959—2015年生物气候图解 Fig.3 The climate diagram of the Qinling Mountains during 1959—20151.年均温;2.年均降水量;3.温度降水的观测年数;4.湿润期(阴影区);5.月降雨量平均超过100mm
图3为秦岭1959—2015年生物气候图解。据32个气象站的数据统计,研究区内年均温13.1℃,最热月7月均温24.9℃,最冷月1月均温0.5℃,年均降水量758.57mm。图3显示,全年温度曲线在下,降水曲线在上,7—9月降雨量均在100mm以上。可知,秦岭地区水热组合条件较好,气候湿润,利于植物生长。
2.2 52年来物候变化特征
图4为1964—2015年物候始期与末期的变化趋势。由图4和表2可知,52年间,7种木本植物的物候始期与末期变化趋势均一致,物候始期提前,平均提前1.2d/10a(P<0.05);物候末期推迟,平均推迟3.5d/10a(P<0.01)。即物候生长期延长,物候末期的推迟趋势较始期的提前趋势表现得更为显著,变化速率更快。
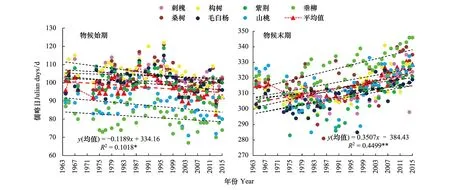
图4 1964—2015年物候始期与末期变化Fig.4 The change in the start and end of plant phenophases during 1964—2015
7个树种物候始期发生于第67—122天(3—4月),物候末期发生于第281—346天(10—12月上旬)。物候始期,构树的变化速率最大2.3d/10a(P<0.01),紫荆和山桃的变化显著,其他树种的变化均不显著;物候末期,垂柳的变化速率最大4.9d/10a (P<0.01),除刺槐变化不显著及山桃(P<0.05)外,其他树种均呈极显著变化。
2.3 物候突变前后变化特征
2.3.1 物候突变的判定
图5为1964—2015年7个树种物候始期均值与末期均值的二项式拟合曲线与斜率线变化趋势。显示,各树种物候始期与末期的变化曲线均呈朝向相反的二次抛物线形式。表明,物候始期的提前与末期的推迟效应,存在一个转折点。
*P<0.05;**P<0.01
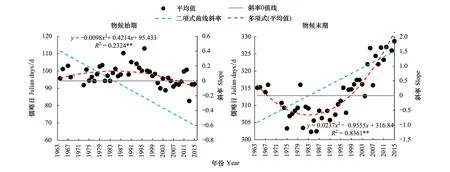
图5 1964—2015年物候始期与末期变化与曲线斜率Fig.5 The change in the start and end of plant phenophases and curve slope during 1964—2015
由图5可知,物候始期的突变年份发生于1985年,末期的突变年份发生于1984年。这与以往研究气温突变[41-44]结论“通过阶段性分析和滑动平均方法得到全国年均温变化的转折时间1984年”较为一致。通过对每一个树种物候期的二项式曲线进行一阶求导发现,各树种的突变年份各不相同,除垂柳的突变发生于20世纪70年代末外,其他树种均发生于20世纪80年代。
2.3.2 物候突变前变化特征
图6为突变前物候始末期的变化趋势。由图6和表3可知,22年间,物候始期,7个树种中垂柳、山桃、构树、刺槐呈不明显的下降趋势,其他树种呈不明显的上升趋势,即突变前物候始期变化不显著;物候末期,除垂柳外,其他树种均呈不同程度的提前趋势,山桃和刺槐达到了极显著,桑树达到了显著,即突变前物候末期呈显著的提前趋势,提前速率4.6d/10a (P<0.01)。
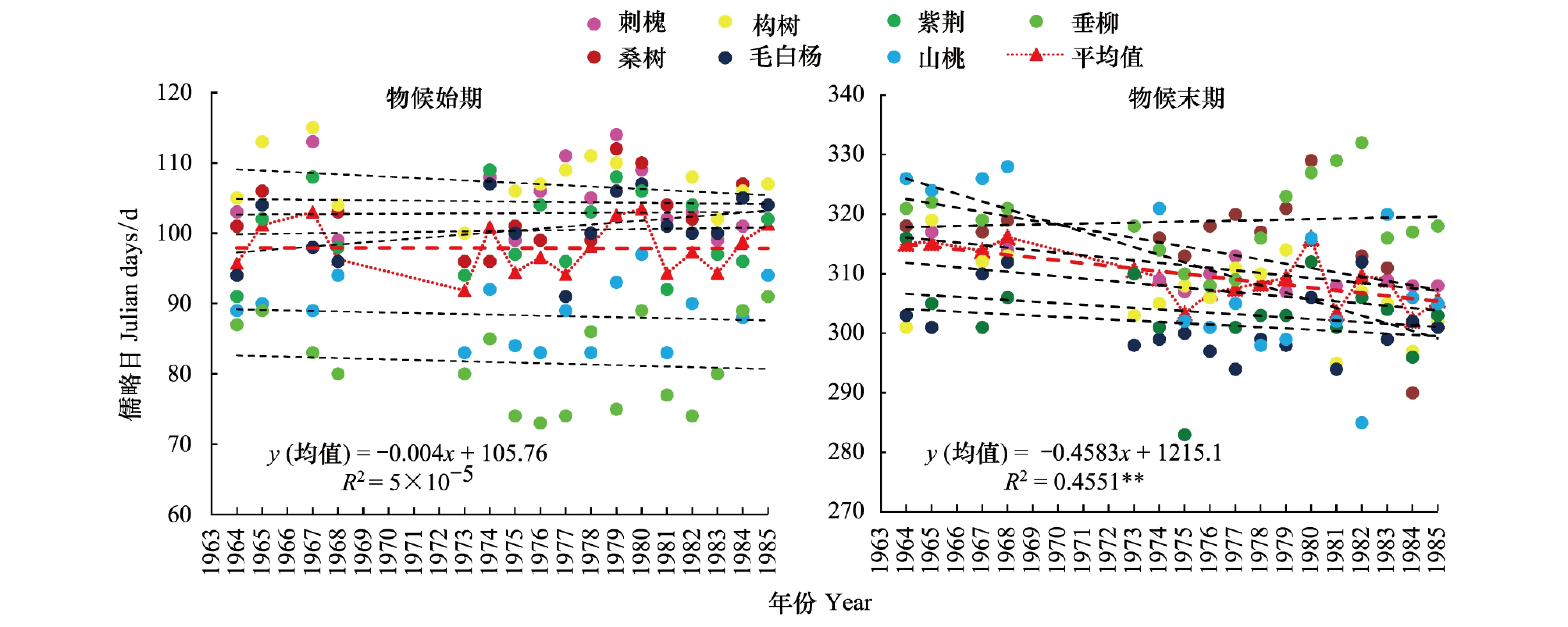
图6 1964—1985年物候始期与末期变化Fig.6 The change in the start and end of plant phenophases during 1964—1985
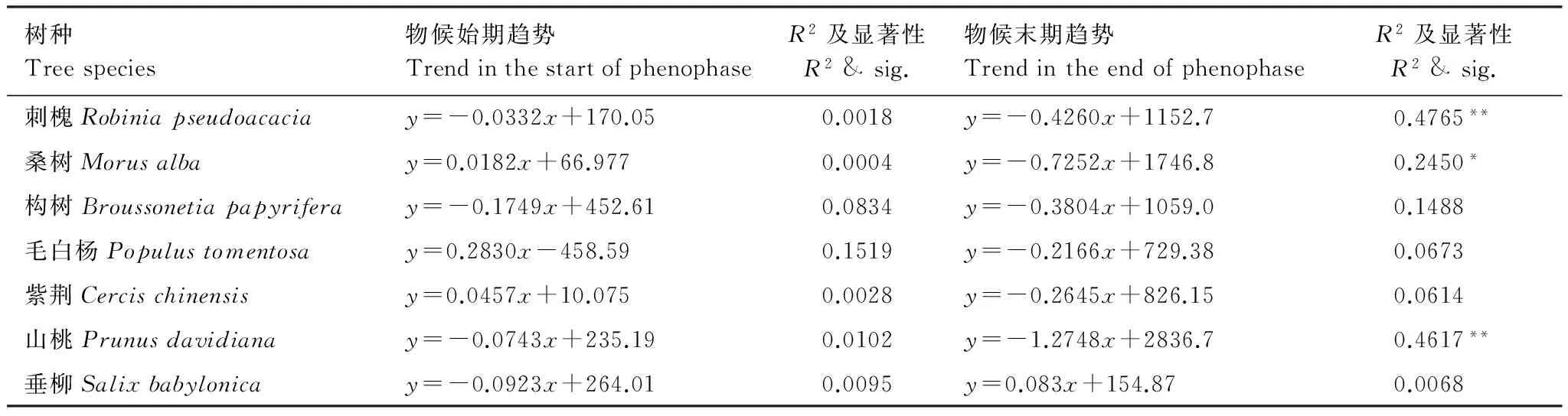
树种Treespecies物候始期趋势TrendinthestartofphenophaseR2及显著性R2&sig.物候末期趋势TrendintheendofphenophaseR2及显著性R2&sig.刺槐Robiniapseudoacaciay=-0.0332x+170.050.0018y=-0.4260x+1152.70.4765**桑树Morusalbay=0.0182x+66.9770.0004y=-0.7252x+1746.80.2450*构树Broussonetiapapyriferay=-0.1749x+452.610.0834y=-0.3804x+1059.00.1488毛白杨Populustomentosay=0.2830x-458.590.1519y=-0.2166x+729.380.0673紫荆Cercischinensisy=0.0457x+10.0750.0028y=-0.2645x+826.150.0614山桃Prunusdavidianay=-0.0743x+235.190.0102y=-1.2748x+2836.70.4617**垂柳Salixbabylonicay=-0.0923x+264.010.0095y=0.083x+154.870.0068
*P<0.05;**P<0.01
考虑到早期数据资料的不连续性及历史原因,剔除1973之前的数据,1973—1985年,7个树种物候始期均值呈推迟趋势,推迟速率3.0d/10a;末期均值呈提前趋势,提前速率1.9d/10a,生长期缩短,与1964—1985年间生长期的变化相比,此结论相对更为合理。
2.3.3 物候突变后变化特征
图7为突变后物候始末期的变化趋势。由图7和表4可知,物候始期,7个树种均呈极显著的提前趋势,平均提前4.3d/10a(P<0.01);物候末期均呈极显著的推迟趋势,平均推迟8.4d/10a (P<0.01)。即突变后的31年间,物候生长期延长,末期由突变前的提前趋势转变为极显著的推迟趋势,且变化速率和显著性均高于始期。
物候始期,展叶最晚的构树与最早的垂柳在1985年相差26d,到2015年相差21d,二者展叶期31年间缩短了5d;物候末期,叶全变色最晚的垂柳与最早的紫荆在1985年相差26d,到2015年相差18d,二者叶全变色期31年间缩短了8d。表明,突变后,随着气候变暖,植物对气候的感应趋于一致,始期和末期变化均表现出“趋同效应”。
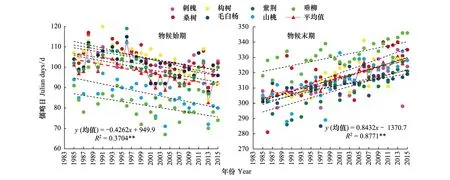
图7 1985—2015年物候始期与末期变化Fig.7 The change in the start and end of plant phenophases during 1985—2015
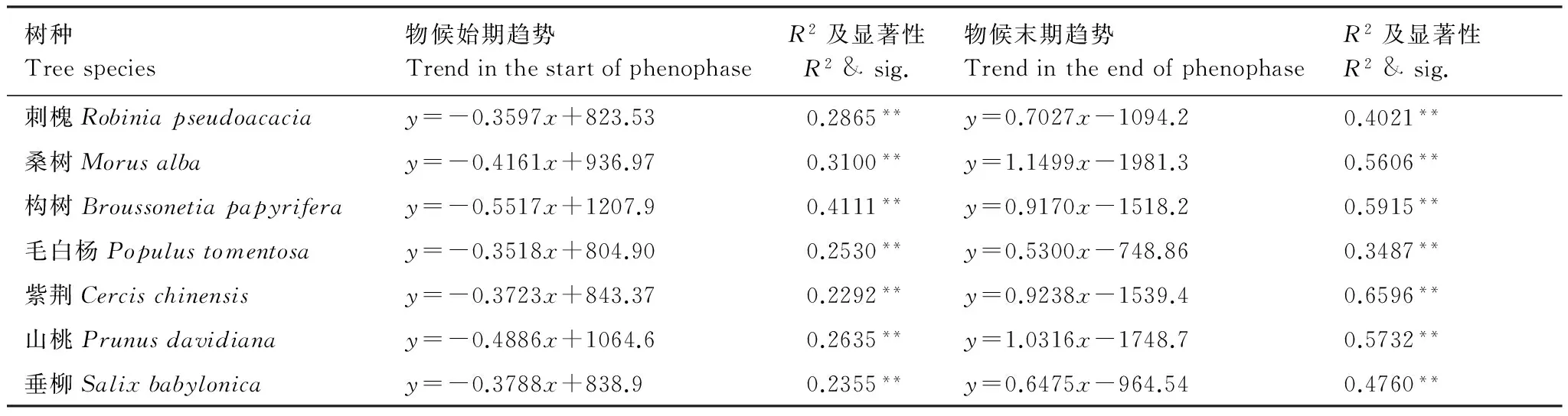
树种Treespecies物候始期趋势TrendinthestartofphenophaseR2及显著性R2&sig.物候末期趋势TrendintheendofphenophaseR2及显著性R2&sig.刺槐Robiniapseudoacaciay=-0.3597x+823.530.2865**y=0.7027x-1094.20.4021**桑树Morusalbay=-0.4161x+936.970.3100**y=1.1499x-1981.30.5606**构树Broussonetiapapyriferay=-0.5517x+1207.90.4111**y=0.9170x-1518.20.5915**毛白杨Populustomentosay=-0.3518x+804.900.2530**y=0.5300x-748.860.3487**紫荆Cercischinensisy=-0.3723x+843.370.2292**y=0.9238x-1539.40.6596**山桃Prunusdavidianay=-0.4886x+1064.60.2635**y=1.0316x-1748.70.5732**垂柳Salixbabylonicay=-0.3788x+838.90.2355**y=0.6475x-964.540.4760**
*P<0.05;**P<0.01.
图8直观地表示了突变后7个树种物候始期与末期每5年均值的年代际变化。显示,物候始期的年代际变化整体呈明显的提前趋势,末期呈明显的推迟趋势。而且,物候始期展叶相对较早的树种自2001—2005年始提前速率减缓,一方面,这与植物自身对气候变化有了一定的适应性,能够通过自身的调控机制,采取相应生活史的改变来适应环境变化有关;另一方面,据资料[11,45-46]显示,全球及我国的气温自1998年以来出现变暖趋缓现象,表明植物物候变化随着气候变化而变化,且存在一定的滞后性。

图8 突变后物候始期与末期年代际变化Fig.8 The interdecadal variation in the start and end of plant phenophases after the abrupt change
2.4 物候变化差异性
2.4.1 树种差异性
图9为15个树种物候突变后的变化趋势。由图9和表5及表4可知,无论是乔木、灌木还是藤本植物,均表现为物候始期提前,平均提前3.9d/10a;物候末期推迟,平均推迟7.5d/10a;生长期延长。
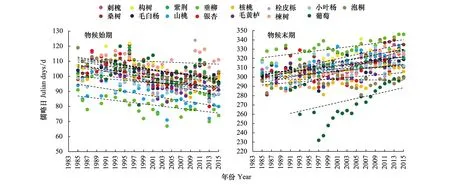
图9 1985—2015年15种植物物候始期与末期变化Fig.9 The change in the start and end of plant phenophases about 15 tree species during 1985—2015
11个乔木树种中,物候始期平均提前3.8d/10a,物候末期平均推迟6.8d/10a,生长期延长。其中,物候始期,构树的变化速率最大5.5d/10a(P<0.01),物候末期,楝树的速率最大17.2d/10a(P<0.01);3个灌木树种中,物候始期平均提前4.8d/10a;物候末期平均推迟8.9d/10a,生长期延长。其中,物候始期,毛黄栌的变化速率最大6.7(P<0.01),物候末期,桑树的速率最大11.5(P<0.01);以及藤本植物葡萄的物候始期提前2.9d/10a,末期推迟11.5d/10a,生长期延长。
由以上分析可知,3大类型树种表现各异,物候始期的提前速率呈藤本、乔木、灌木依次增大,而末期的推迟速率则是藤本、灌木、乔木依次减小,即不同树种对气候变化的敏感性存在类型和种群的差异。
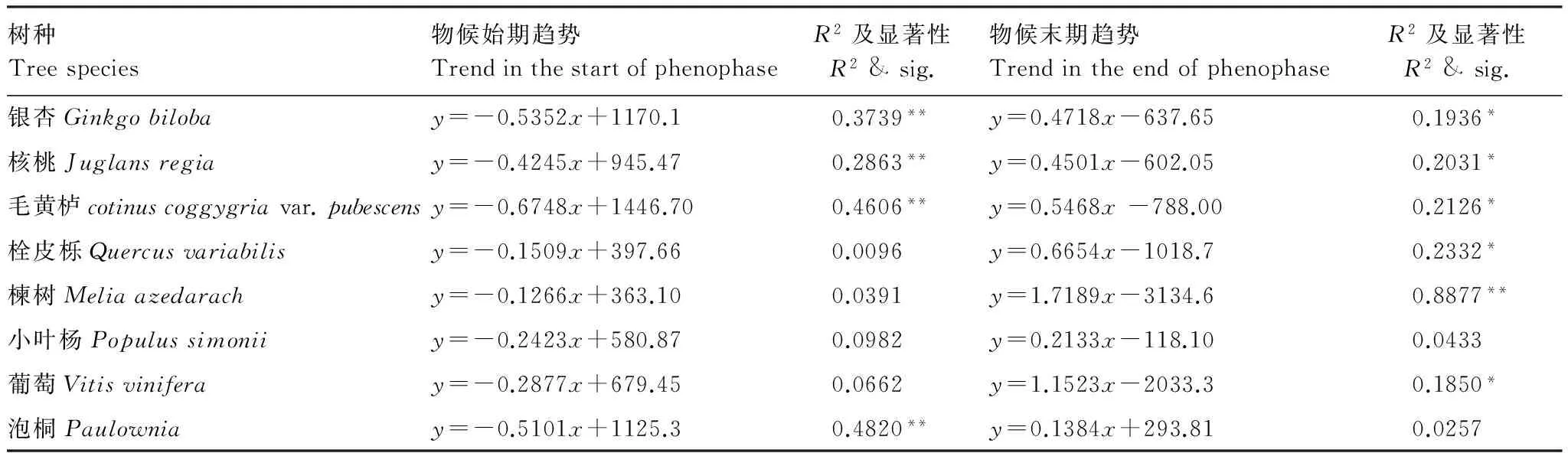
表5 1985—2015年8树种植物物候始末期变化趋势
*P<0.05;**P<0.01
2.4.2 空间差异性
秦岭南北气温差异明显,植物物候对气候变化的响应是否存在着南北差异?本文选择了观测资料完整、时间序列较长的核桃(1982—2015年)和垂柳(1982—2008年) 2个树种,进行秦岭南北坡物候变化比较。
图10为秦岭南北坡2个树种物候始末期的变化趋势。显示,物候始期,北坡核桃提前了3.1d/10a(P<0.05),南坡则提前1.6d/10a,北坡垂柳提前了3.9d/10a(P<0.05),南坡则提前2.0d/10a;物候末期,北坡核桃推迟了4.7d/10a(P<0.01),南坡则推迟6.9d/10a(P<0.01),北坡垂柳推迟了3.2d/10a,南坡则推迟10.0d/10a。 可知,无论是核桃还是垂柳,均表现为始期提前,末期推迟,生长期延长,即不同树种在南北坡的变化趋势是一致的,但北坡始期的提前速率均高于南坡,而南坡末期的推迟速率均高于北坡。
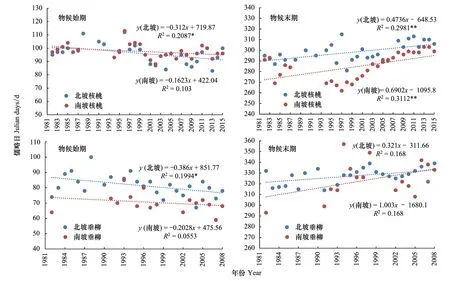
图10 秦岭南北坡物候始期与末期变化比较Fig.10 The Comparison of the start and end of plant phenophases in South & North Slopes of the Qinling Mountains
3 结论与讨论
(1) 52年来,7种木本植物物候始期均呈提前趋势,提前速率1.2d/10a,物候末期均呈推迟趋势,推迟速率3.5d/10a,生长期均呈延长趋势。据资料[9,37]显示,秦岭近50多年气温呈增加趋势,春季与秋季的干暖化趋势明显,春季增温率0.31℃/10a,秋季为0.2℃/10a。表明,温度的升高是导致秦岭地区物候始期提前,末期推迟,这可能是物候生长期延长的重要原因。从以上分析发现,秋季的增温速率低于春季,而物候末期的变化率却显著高于始期,这一结论在我国学者对不同区域物候研究中也得到证实,北京木本植物40多年春季物候提前了2.7d/10a[47],秋季物候推迟了4.9d/10a[48];西安2003—2011年与1963—1996年相比,展叶始期提前了5.54d,叶开始变色期推迟了10.59d[49];贵阳1978—2007 年展叶始期平均提前5.2d/10a,叶完全变色期推迟了6.3d/10a[50]。由此可知,物候始期与末期对温度变化的响应具有不一致性。
(2) 秦岭植物物候突变发生于1980s,始期于1985年,末期于1984年,突变后,物候特征发生了显著变化。始期的提前速率较突变前显著加快,提前速率4.3d/10a,末期由突变前的提前趋势转变为极显著的推迟趋势,推迟速率8.4d/10a;始期与末期的变化均表现出“趋同效应”;物候年代际变化显示,始期自2001—2005年起提前速率减缓,植物对气候变化的响应表现出适应性及滞后性。据资料[9,37]显示,秦岭气温突变发生于1984年,突变前,秦岭年均温的升温率为0.09℃/10a;而突变后达0.58℃/10a,尤以春季增温最为显著,达1.09℃/10a,而秋季则为0.40℃/10a。可见,气温是影响物候始末期变化的主导因素。
(3) 秦岭植物物候变化存在树种差异。无论是乔木、灌木还是藤本植物,均表现为始期提前,末期推迟,生长期延长;但3大类型的树种表现各异,物候始期的提前速率呈藤本、乔木、灌木依次增大,而末期的推迟速率则呈藤本、灌木、乔木依次减小。这可能与树种自身的生理特性对气候变化反应的敏感性不同有关。秦岭山系庞大,地形复杂,区域气候差异大,物种多样性丰富,如何系统、快速、及时地研究反映气候变化对植被生态系统的影响,单以地面观测数据难以实现区域尺度上植物物候变化的动态监测,因此进一步结合高精度的遥感监测数据来获取相应的物候表征参数,对秦岭山地物候的综合研究就具有重要意义。
(4) 秦岭物候变化存在南北差异。不同树种在南北坡的变化趋势具有一致性,无论是核桃还是垂柳均表现为始期提前,末期推迟,生长期延长;但北坡始期的提前速率均高于南坡,而南坡末期的推迟速率均高于北坡。这可能与区域温度变化的差异性有关,据资料显示, 我国增温趋势总体上呈北高南低的分布特征,北方升温幅度较南方大、变化速度快[51];秦岭50多年来春季北坡升温率0.38℃/10a,南坡为0.24℃/10a,而秋季北坡的升温率0.24℃/10a,南坡为0.16℃/10a[9,37]。由此可知,南北坡在物候始期的提前速率与温度变化速率相对一致。而在物候末期的推迟速率则不一致。这主要源于南北气候的差异性,可能与物候末期北坡受温带季风的影响,而南坡则是受亚热带季风气候的影响有关,秋季物候变化的复杂性和受多因素的综合影响[50],这一问题有待深入研究。
[1] 竺可桢, 宛敏渭. 物候学. 北京: 科学出版社, 1973.
[2] 张福春. 物候. 北京: 气象出版社, 1985.
[3] 陆佩玲, 于强, 贺庆棠. 植物物候对气候变化的响应. 生态学报, 2006, 26(3): 923- 929.
[4] 葛全胜, 戴君虎, 郑景云. 物候学研究进展及中国现代物候学面临的挑战. 中国科学院院刊, 2010, 25(3): 310- 316.
[5] Root T L, Price J T, Hall K R, Schneider S H, Rosenzweig C, Pounds J A. Fingerprints of global warming on wild animals and plants. Nature, 2003, 421(6918): 57- 60.
[6] Schwartz M D. Monitoring global change with phenology: the case of the spring green wave. International Journal of Biometeorology, 1994, 38(1): 18- 22.
[7] 莫非, 赵鸿, 王建永, 强生才, 周宏, 王绍明, 熊友才. 全球变化下植物物候研究的关键问题. 生态学报, 2011, 31(9): 2593- 2601.
[8] 方修琦, 余卫红. 物候对全球变暖响应的研究综述. 地球科学进展, 2002, 17(5): 714- 719.
[9] 白红英. 秦巴山区森林植被对环境变化的响应. 北京: 科学出版社, 2014.
[10] Menzel A. Phenology: it′s importance to the global change community. Climatic Change, 2002, 54(4): 379- 385.
[11] IPCC. Summary for policymakers//Stocker T F, Qin D, Plattner G K, Tignor M, Allen S K, Boschung J, Nauels A, Xia Y, Bex V, Midgley P M, eds. of Climate Change 2013: the Physical Science Basis Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press, 2013.
[12] 秦大河, Stocker T. IPCC第五次评估报告第一工作组报告的亮点结论. 气候变化研究进展, 2014, 10(1): 1- 6.
[13] Ren G Y, Ding Y H, Zhao J C, Zheng J Y, Wu T W, Tang G L, Xu Y. Recent progress in studies of climate change in China. Advances in Atmospheric Sciences, 2012, 29(5): 958- 977.
[15] Parmesan C. Influences of species, latitudes and methodologies on estimates of phenological response to global warming. Global Change Biology, 2007, 13(9): 1860- 1872.
[16] Fitter A H, Fitter R S R. Rapid changes in flowering time in British plants. Science, 2002, 296(5573): 1689- 1691.
[17] Gordo O, Sanz J J. Long-term temporal changes of plant phenology in the western Mediterranean. Global Change Biology, 2009, 15(8): 1930- 1948.
[18] Beaubien E G, Freeland H J. Spring phenology trends in Alberta, Canada: links to ocean temperature. International Journal of Biometeorology, 2000, 44(2): 53- 59.
[19] Abu-Asab M S, Peterson P M, Shetler S G, Orli S S. Earlier plant flowering in spring as a response to global warming in the Washington, DC, area. Biodiversity & Conservation, 2001, 10(4): 597- 612.
[20] Matsumoto K, Ohta T, Irasawa M, Nakamura T. Climate change and extension of theGinkgobilobaL. growing season in Japan. Global Change Biology, 2003, 9(11): 1634- 1642.
[21] Primack R B, Higuchi H, Miller-Rushing A J. The impact of climate change on cherry trees and other species in Japan. Biological Conservation, 2009, 142(9): 1943- 1949.
[22] Chen X Q, Xu L. Phenological responses ofUlmuspumila(Siberian Elm) to climate change in the temperate zone of China. International Journal of Biometeorology, 2012, 56(4): 695- 706.
[23] Lu P L, Yu Q, Liu J D, He Q T. Effects of changes in spring temperature on flowering dates of woody plants across China. Botanical Studies, 2006, 47(2): 153- 161.
[24] Schwartz M D, Chen X Q. Examining the onset of spring in China. Climate Research, 2002, 21(2): 157- 164.
[25] 王连喜, 陈怀亮, 李琪, 余卫东. 植物物候与气候研究进展. 生态学报, 2010, 30(2): 447- 454.
[26] Menzel A. Plant phenological anomalies in Germany and their relation to air temperature and NAO. Climatic Change, 2003, 57(3): 243- 263.
[27] Penuelas J, Filella L. Herbaria century record of increasing eutrophication in Spanish terrestrial ecosystems. Global Change Biology, 2001, 7(4): 427- 433.
[28] Schwartz M D, Reiter B E. Changes in North American spring. International Journal of Climatology, 2000, 20(8): 929- 932.
[29] 戴君虎, 王焕炯, 葛全胜. 近50年中国温带季风区植物花期春季霜冻风险变化. 地理学报, 2013, 68(5): 593- 601.
[30] 仲舒颖, 郑景云, 葛全胜. 近40年中国东部木本植物秋季叶全变色期变化. 中国农业气象, 2010, 31(1): 1- 4.
[31] 李荣平, 周广胜. 1980- 2005年中国东北木本植物物候特征及其对气温的响应. 生态学杂志, 2010, 29(12): 2317- 2326.
[32] 郑景云, 葛全胜, 郝志新. 气候增暖对我国近40年植物物候变化的影响. 科学通报, 2002, 47(20): 1582- 1587.
[33] 常兆丰, 邱国玉, 赵明, 杨自辉, 韩富贵, 仲生年, 李爱德, 刘淑娟. 民勤荒漠区植物物候对气候变暖的响应. 生态学报, 2009, 29(10): 5195- 5206.
[34] 韩福贵, 徐先英, 王理德, 王键, 张应昌, 韩生慧. 民勤荒漠区典型草本植物马蔺的物候特征及其对气候变化的响应. 生态学报, 2013, 33(13): 4156- 4164.
[35] Cleland E E, Chuine I, Menzel A, Mooney H A, Schwartz M D. Shifting plant phenology in response to global change. Trends in Ecology & Evolution, 2007, 22(7): 357- 365.
[36] Ge Q S, Wang H J, Rutishauser T, Dai J H. Phenological response to climate change in China: a meta-analysis. Global Change Biology, 2015, 21(1): 265- 274.
[37] 高翔, 白红英, 张善红, 贺映娜. 1959- 2009年秦岭山地气候变化趋势研究. 水土保持通报, 2012, 32(1): 207- 211.
[38] 贺映娜. 秦岭植被物候期及遥感生长季的变化研究[D]. 西安: 西北大学, 2012.
[39] 马新萍, 白红英, 贺映娜, 秦进. 基于NDVI的秦岭山地植被遥感物候及其与气温的响应关系——以陕西境内为例. 地理科学, 2015, 35(12): 1616- 1621.
[40] 夏浩铭, 李爱农, 赵伟, 边金虎, 雷光斌. 2001- 2010年秦岭森林物候时空变化遥感监测. 地理科学进展, 2015, 34(10): 1297- 1305.
[41] 王少鹏, 王志恒, 朴世龙, 方精云. 我国40年来增温时间存在显著的区域差异. 科学通报, 2010, 55(16): 1538- 1543.
[42] Ding Y H, Ren G Y, Zhao Z C, Xu Y, Luo Y, Li Q P, Zhang J. Detection, causes and projection of climate change over China: an overview of recent progress. Advances in Atmospheric Sciences, 2007, 24(6): 954- 971.
[43] 陈隆勋, 周秀骥, 李维亮, 罗云峰, 朱文琴. 中国近80年来气候变化特征及其形成机制. 气象学报, 2004, 62(5): 634- 646.
[44] 任国玉, 郭军, 徐铭志, 初子莹, 张莉, 邹旭凯, 李庆祥, 刘小宁. 近50年中国地面气候变化基本特征. 气象学报, 2005, 63(6): 942- 956.
[45] 丁一汇, 柳艳菊, 梁苏洁, 马晓青, 张颖娴, 司东, 梁萍, 宋亚芳, 张锦. 东亚冬季风的年代际变化及其与全球气候变化的可能联系. 气象学报, 2014, 72(5): 835- 852.
[46] 苏京志, 温敏, 丁一汇, 郜永祺, 宋亚芳. 全球变暖趋缓研究进展. 大气科学, 2016, 40(6): 1143- 1153.
[47] 韩超, 郑景云, 葛全胜. 中国华北地区近40年物候春季变化. 中国农业气象, 2007, 28(2): 113- 117.
[48] 仲舒颖, 郑景云, 葛全胜. 1962- 2007年北京地区木本植物秋季物候动态. 应用生态学报, 2008, 19(11): 2352- 2356.
[49] Dai J H, Wang H J, Ge Q S. Multiple phenological responses to climate change among 42 plant species in Xi′an, China. International Journal of Biometeorology, 2013, 57(5): 749- 758.
[50] 白洁, 葛全胜, 戴君虎. 贵阳木本植物物候对气候变化的响应. 地理研究, 2009, 28(6): 1606- 1614.
[51] 尹云鹤, 吴绍洪, 陈刚. 1961- 2006年我国气候变化趋势与突变的区域差异. 自然资源学报, 2009, 24(12): 2147- 2157.
VariationinplantphenologyintheQinlingMountainsfrom1964—2015inthecontextofclimatechange
DENG Chenhui1,BAI Hongying1,*,ZHAI Danping1,GAO Shan2,HUANG Xiaoyue1,MENG Qing1,HE Yingna1
1CollegeofUrbanandEnvironmentalScience,NorthwestUniversity,Xi′an710127,China2Xi′anMeteorologicalBureau,Xi′an710016,China
In the context of global climate change, plant phenology has become the subject of intensive research. Mountains are considered indicators of global climate change. The Qinling mountain region, known as a unique geographical unit in China because of its ecological sensitivity and vulnerability, underwent a significant temperature increase in the last 50 years. Based on phenological data collected from 1964—2015, 17 plant species, including trees, shrubs, and vines, were selected as the subjects of the present study. This paper analyzes and discusses changing conditions and changes in plant phenology in the context of global climate change in the Qinling Mountains. Our analyses generated four primary results. First, over the past 52 years, the beginning of the plant phenophase has advanced at a rate of 1.2 days decade-1, and the end of the plant phenophase has delayed at a rate of 3.5 days decade-1, causing a significant prolongation of the growth period. The primary reason for this prolongation is the rising temperature in the region. There was a inconsistency on the response at the beginning and the end of the phenophase to temperature. Second, the abrupt change of phenophase occurred in the 1980s in Qinling area, the start of phenophase occurred in 1985 andthe end of phenophase did in 1984. After the abrupt change occurs, plant phenological characteristics changed significantly, compared with abrupt change before, showing that the advanced rate in the start of phenophase was faster, and the trend in the end of the phenophase turned into a very significant delay; the rate of change and the significance at the end of the plant phenophase were higher than that at the beginning of the period. These changes in the beginning and end points of plant phenophases manifest as a "convergence effect". Interdecadal variations in phenology indicate that the rate of advance in the beginning of plant phenophases slowed from 2001 to 2005, and that the response of plants to climate change showed qualities of adaptability and hysteresis. These results indicate that the dominant factor leading to changes in the beginning and end points of the plant phenophase was temperature, and that the response of plant phenology to changes in temperature is particularly sensitive, further demonstrating that there is a correlation between plant phenophase and climate change. Third, changes in phenophase varied by species. The advancement of the beginning of the phenophase promoted the prevalence of shrubs, trees, and vines, in this order, at the beginning of the phenophase, whereas the delay in the ending of the phenophase decreased the abundance of trees, shrubs, and vines, in this order, at the end. These differences in abundance are related to differences in physiological characteristics between the different species in response to climate change. Finally, plant responses to climate change in the Qinling Mountains differed along a north-south axis. The rate of advance in the beginning of the phenophase was higher in the northern slope than in the southern slope, whereas the rate of delay in ending of the phenophase was more severe in the southern slope than in the northern slope.
plant phenology; variation in characteristics; abrupt change in phenophase; difference; the Qinling region
国家林业公益性行业科研专项(201304309)
2017- 02- 20; < class="emphasis_bold">网络出版日期
日期:2017- 08- 18
*通讯作者Corresponding author.E-mail: hongyingbai@163.com
10.5846/stxb201702200277
邓晨晖,白红英,翟丹平,高山,黄晓月,孟清,贺映娜.气候变化背景下1964—2015年秦岭植物物候变化.生态学报,2017,37(23):7882- 7893.
Deng C H,Bai H Y,Zhai D P,Gao S,Huang X Y,Meng Q,He Y N.Variation in plant phenology in the Qinling Mountains from 1964—2015 in the context of climate change.Acta Ecologica Sinica,2017,37(23):7882- 7893.
